火炬树横走侧根不定芽的发生及其形态解剖学特性的研究
2016-11-10许玉凤徐丹阳郭文铮邵美妮
许玉凤 徐丹阳 郭文铮 邵美妮 关 萍 曲 波*
(1.沈阳农业大学生物科学技术学院,沈阳 110866; 2.沈阳农业大学林学院,沈阳 110866)
火炬树横走侧根不定芽的发生及其形态解剖学特性的研究
许玉凤1徐丹阳2郭文铮1邵美妮1关 萍1曲 波1*
(1.沈阳农业大学生物科学技术学院,沈阳 110866;2.沈阳农业大学林学院,沈阳 110866)
火炬树是一种典型的主要以根蘖进行繁殖的木本植物,超强的根蘖繁殖能力和极高的适应能力,使其具有非常强的入侵性。试验通过野外挖根调查,研究了火炬树横走侧根不定芽的分布规律,用常规石蜡切片法研究了不定芽的发生特点,并且对根蘖苗的发生进行了物候学观察和分析。结果表明:火炬树的根系分布较浅,横走侧根分布在地表以下15 cm范围内,其中2~5 cm土层内侧根上的不定芽分布最多。不定芽在整个生长季都有萌发,但春季萌发数量最多。形态解剖学研究表明不定芽起源于横走侧根的中柱鞘细胞,为根源性克隆植物。根蘖苗的形成在时间上要比地上母株和其他树木萌发早,林分的充分光照使其迅速生长,增强了竞争力,为其种群更新和扩张创造了条件,促进了其入侵。
火炬树;克隆生长;根蘖;解剖学;入侵
火炬树(RhustyphinaL.)漆树科(Anacardiaceae)盐肤木属(Rhus)的落叶灌木或小乔木,原产于北美,现在欧洲、亚洲及大洋洲等许多国家都有栽培。我国于1959年由中国科学院植物研究所北京植物园引种,1974年以来在全国各省区推广种植[1]。火炬树雌雄异株,具有喜光、耐寒、耐旱、耐瘠薄、耐盐碱、适应性广等优点;其雌花序及果穗鲜红色,花形独特,形同火炬,秋季来临叶片变成红色,十分美观;根系分布较浅,水平根系非常发达,因此它是干旱地区植被恢复与构建、环境美化和护坡、风景林营建的优良树种之一。由于火炬树具有超强的根蘖繁殖能力和抗逆性等特点,使其具有非常强大的生命力、种间竞争力和种群扩张力,影响了本地植物的生长,具有外来入侵植物的基本特征。目前,一些学者已经把火炬树定为外来入侵树种,如果继续大范围推广种植会对生态以及生物多样性造成严重威胁。根据有关入侵种的定义火炬树目前已经达到入侵阶段,不适合继续作为北京绿化的主要树种,应该对其采取一定的控制措施[2]。李传文等调查发现1993年营造的火炬树、黄栌块状混交林,现在已经很难寻找到黄栌的踪迹,因此认为火炬树具有外来入侵植物的基本特征,应该引起警惕[3]。
火炬树,主要以根蘖繁殖方式进行种群更新和扩张,种植过火炬树的土壤很难清除干净。火炬树超强的克隆繁殖特点使其侵占本地物种的生态位,与本地植物竞争水分、营养和光照,影响本地植物的生长,对其入侵性具有非常重要的贡献。
目前国内外对火炬树入侵机理方面的研究主要集中在化感作用[4]、表型可塑性[5]、生物学特性[6]、生理生态特性[7]等方面,而关于火炬树横走侧根不定芽的发生、分布等研究报道相对较少。本研究观察火炬树横走侧根的空间分布和克隆分株的特点,借助植物解剖学的研究方法探讨火炬树横走侧根的解剖结构及其不定芽形成的特点,结合根蘖苗发生的物候学观察分析,旨在为解析火炬树入侵机制提供形态解剖学基础,为进一步探讨火炬树根蘖苗发生、生长和空间分布格局响应生境异质性资源提供理论依据。
1 试验地区与试验方法
1.1 试验区概况
本试验地点位于沈阳农业大学天柱山。该地区属于大陆性温带季风气候,春秋两季气温变化迅速为时较短,冬季时间较长,严寒少雨,夏季多雨而炎热,年平均降水量为712.8 mm[8]。2008年将火炬树作为护坡植物栽植,平均胸径为10 cm。该地区植物群落主要为紫穗槐与火炬树混合栽植。林中还伴有大量的由根蘖繁殖而生的火炬树幼苗,同时还向外延伸了近7 m,侵入到了农田。
1.2 试验方法
1.2.1 火炬树横走侧根及其不定芽分布的调查
于2014年4月5日在试验地内选取成年单株,采用挖根的方法,以基株为中心,向四周沿横走侧根扩展,在有根蘖苗不定芽分布的区域挖取地下横走侧根,调查记录横走侧根在土壤中的分布深度、粗度以及着生不定芽根蘖苗的状况,并拍照。
1.2.2火炬树横走侧根及其不定芽形态解剖学特性研究
在调查地下横走侧根及不定芽分布的同时,在试验地内挖取带有不定芽的侧根,迅速带回实验室,用清水洗净,选取长有不定芽的部分,用刀片切成0.5 cm左右的小段放入事先配好的FAA固定液中固定,固定时间在24 h以上。用常规石蜡切片法制片,酒精脱水,二甲苯透明,切片厚度为12 μm,用番红、固绿染色,加拿大树胶封片。用Olympus光学显微镜观察侧根的横切面、不定芽的发生位置并用蔡司(AxioCam MRc CCD)显微照相系统拍照。
1.2.3 火炬树根蘖苗发生的物候学观察
在林地内设10个1 m×1 m的样方,从2014年4月1日开始,每月末调查一次,注意观察地上的枝条和地下横走侧根不定芽发生的时间,根蘖苗的萌发调查一直持续到9月末。
2 结果与分析
2.1 火炬树横走侧根上不定芽的生长与分布特点
通过挖根观察发现,能够发生不定芽的横走侧根通常分布在近地表5 cm以内,发生不定芽最多的横走侧根应该在近地表1~2 cm的范围内。火炬树横走侧根发生根蘖苗的能力非常强,有的甚至裸露在地表的侧根也能产生根蘖苗(图1A)。着生不定芽的横走侧根直径平均在0.3 cm左右,最细的直径为0.2 cm上也有不定芽发生,最粗的直径可以达0.6 cm以上。通过栽培试验观察,2年生以上的火炬树横走侧根上即可长出不定芽,而且不定芽在一级横走侧根和其分枝上均有生长。火炬树横走侧根上的不定芽可以有一个单生(图1B),一般两个不定芽丛生在一起的比较多(图1C),有时也有多个不定芽聚集分布在一起的情况(图1D),不定芽之间的距离、大小也都不尽相同。在早些年生长出来的根蘖苗基部也有不定芽的发生(图1E)。有的不定芽分布于横走侧根的同一侧(图1F),也有的不定芽环绕在横走侧根的不同位置上(图1G)。通过后期的观察,不是所有的不定芽都会萌发形成根蘖苗,一般丛生的不定芽,仅有2~4个芽会萌发,其它的芽一直处于休眠状态或慢慢地干瘪退化。

图1 火炬树横走侧根不定芽的生长和分布Fig.1 The growth and distribution of adventitious buds on the horizontal lateral roots of R.typhina
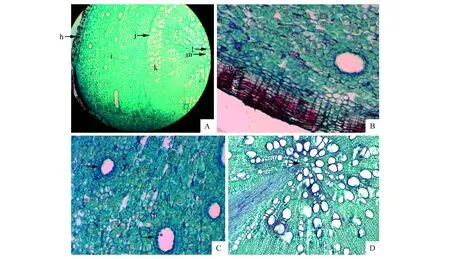
图2 火炬树横走侧根解剖结构 A.横切面;B.外面的周皮组织;C.分泌道;D.不定芽的发生部分 h.周皮;i.次生韧皮部;j.维管形成层;k.次生木质部;l.初生木质部;m.不定芽的发生部位;n.木栓层;o.木栓形成层;p.栓内层Fig.2 The anatomical structure of horizontal lateral roots of R.typhina A.Transverse; B.The periderm out; C.The secretory canals; D.The occurrence of adventitious buds h. Pperiderm;i. Secondary phloem;j. Vascular cambium;k. Secondary xylem;l. Primary xylem;m.The occurrence of adventitious bud;n. Cork layer;o. Cork cambium layer;p. Phelloderm
2.2火炬树横走侧根及其不定芽发生的解剖结构观察
火炬树横走侧根的解剖结构由外向内依次为周皮、次生韧皮部、维管形成层、次生木质部和中央的初生木质部(图2A)。周皮为保护组织,是由木栓层(6层细胞)、木栓形成层(3层细胞)和栓内层(3~4层细胞)组成的(图2B)。维管形成层是由2~3层扁平细胞组成,外侧为次生韧皮部,内侧为次生木质部。韧皮部组织比较发达,其中有许多分泌道(图2C)。分泌道为椭圆形,内层有一圈排列紧密的分泌细胞,近似长方形,排列比较规则,细胞质比较浓。分泌道外围有2~3层薄壁细胞组成的鞘细胞。散生在韧皮部中的分泌道,越靠近外层,其直径越大,鞘细胞也越厚,越靠近内层,鞘细胞也越薄。次生木质部是由导管、管胞、木纤维和木薄壁细胞组成的。中央为初生木质部,火炬树横走侧根为四原型的根。通过观察火炬树横走侧根着生不定芽的连续石蜡切片发现,火炬树的不定芽起源于横走侧根的中柱鞘组织细胞,由于中柱鞘细胞仍保留有分生能力,因此由这些细胞的分裂活动形成了不定芽原基(图2D)。不定芽原基细胞经过分裂、生长和分化形成了在横走侧根表面可观察到的不定芽,由不定芽直接发育成根蘖苗。
2.3 火炬树根蘖苗发生的物候学观察
由图3可知,在沈阳地区,火炬树地下横走侧根不定芽在4月初就开始萌动,且在4月5日以后,在地上就逐渐能看见已经萌发的不定芽,形成幼苗,不定芽形成和萌发的数量在4月份最多,5月份数量逐渐减少,次之。到6月份,火炬树地下横走侧根仍然有不定芽的发生和根蘖苗的形成,但是数量明显减少。在7月份也能看到有不定芽的发生和根蘖苗的形成,但是数量比较少,在8和9月份基本上不再有不定芽和根蘖苗的形成。

图3 在一个生长季里火炬树根蘖苗萌发数量Fig.6 The number of clonal seedings on the horizontal lateral roots of R.typhina in a growth season
通过观察,火炬树地下横走侧根不定芽发生和根蘖苗的形成在时间上要早于火炬树地上多年生枝条和其它树木芽的萌发。也就是说,火炬树地下横走侧根不定芽在4月初就有发生,4月中旬在地上就已经有了大量的根蘖苗出土,而地上多年生枝条芽的萌发时间特别晚,大致在5月中旬才开始萌发。而在此时新生长出来的根蘖苗已经迅速生长高达50 cm左右。待地上的火炬树母株和其它树木芽萌发、展叶,林分郁闭后,林下的光照减弱,根蘖苗的生长速度逐渐减小。
3 讨论
火炬树既可以种子繁殖,又可以营养繁殖,并且根蘖能力极强,造林成活的当年即可形成新的克隆分株,以后还逐年增多,所以在一般土壤条件下,火炬树完全能通过根蘖繁殖建立种群[9~10]。火炬树根系具有旺盛的生长和扩散能力,6年以上的火炬树根系生长最远可达7 m[11]。火炬树主根长到10 cm左右就停止生长,但由主根上生长出很多发达的一级横走侧根,同时一级横走侧根上会分枝出二级横走侧根,这些侧根在土壤中分布比较浅,多数在距土壤表层5 cm以内。侧根上都会长出不定芽,分布密集,长出大量根蘖苗。火炬树超强的克隆繁殖能力和适应性影响了本地植物的生长,降低了生物多样性,对生态环境造成了威胁。由于火炬树水平侧根形态和生长发育的特殊性,可以认为,火炬树具有极强的入侵潜力,入侵行为正在表达[12]。
克隆植物(cIonalplant)是指在自然生境条件下,能通过营养繁殖产生与其亲本在基因型上几乎完全一致的新个体的植物。克隆植物通过营养生长产生的新分株,具有潜在的独立性,可以在空间上移动从而占据新的生境位点,且各分株间可以通过克隆整合相互传递资源和信号,使各种异质性资源得以共享[13~14]。常见的克隆植物类型有根源型,如疏叶骆驼刺(Alhagisparsifolia)[15]、欧洲灌木型山杨(Trembling Aspen)、沙棘(Hippophaerhamnoides)[16]、胡杨(Populuseuphratica)[17]等;根茎型,如箭竹(FargesiaspathaceaFranch)、沙鞭(Psammochloavillosa)等;匍匐茎型,如蛇莓(Duchesneaindica)、白三叶(Trifoliumrepens)等;分蘖型,如黑麦草(Loliumperenne)[13]等。根源型的克隆分株如沙棘、欧洲灌木型山杨、胡杨等起源于根周皮中的木栓形成层。木栓形成层细胞进行分裂、生长和分化进而形成不定芽[13,18]。火炬树的克隆方式是根茎型还是根源性的,通过查阅大量文献并没有明确的解释,有些学者把火炬树当作根茎型克隆植物。本文通过形态观察和解剖学特性两方面的研究证实了,火炬树属于根源性克隆植物,即克隆器官为根或其衍生物的克隆植物[19]。根茎型克隆植物虽然也是靠不定芽长出萌蘖苗,但是其不定芽长在根状茎上,根茎是植物显著变异的地下茎,普遍存在于多年生湿地草本植物中[20],根茎具有明显的节和节间。通过挖根观察火炬树的横走侧根上没有明显的节和节间,并且通过制作石蜡切片观察横走侧根中有明显的中柱鞘,不定芽起源于中柱鞘部分,由于中柱鞘仍保留有分生能力,因此由这些细胞的分裂活动形成了不定芽原基。不定芽原基细胞分裂、生长和分化形成在横走侧根表面可观察到的不定芽,由不定芽直接发育成根蘖苗。
本研究还发现,在火炬树横走侧根和不定芽的解剖结构中均观察到了分泌道。前人的研究中也报道了火炬树的分泌道分布于各个器官中,如叶、叶柄、花、果实、茎、根等,其发育方式为裂生[21]。分泌道在漆树科植物中均有分布。有的植物如臭椿的分泌道和创伤性分泌道在抵御生物和非生物胁迫中起重要作用[22]。火炬树根、茎、叶片和果穗等水浸体液均对小麦等作物的种子萌发具有显著的化感作用[23~24]。郭阿君等人研究了火炬树在生长过程中释放的挥发性物质,其中有大量的萜烯类物质如A-蒎烯、柠檬烯、龙脑、乙酸龙脑酯等,具有杀虫、灭菌、镇痛、降血压等保健功效。火炬树释放的挥发性物质能够有效抑制空气中的细菌[25]。而火炬树分泌的化学物质是否会对外部环境的胁迫有防御作用将有待于进一步研究。
物候现象是指植物的生命活动在时间上对变化的环境所做出的一种反应[26]。在时空变化的环境下,植物对资源的捕获能力和利用效率对其存活、生长和扩散等有重要的影响[27]。植物捕获光能的主要器官是叶片,其发生时间、凋落时间以及发育进程等,也就是叶的物候是植物在季节性和非季节性变化的环境中,为了提高碳积累量而采取的一种适应策略[28]。林下入侵种波氏小檗(也叫日本小檗)Berberisthunbergii展叶时间比本地共生种高丛越桔(也叫北方高丛蓝莓)Vacciniumcorymbosum早1个月,比林冠郁蔽早2周,它可以利用早春林内充足的光照进行光合作用,使其碳积累“补贴”占其整个生长季碳累积量的36%,促进了其入侵[29]。Küppers发现Ribesuvacrispa能成功定居林下也是因为它能提前展叶,延长了生长季,提前进行了碳积累,这决定了它较强的竞争力[30]。火炬树根蘖苗发生的物候学观察结果表明,地上根蘖苗的发生早于地上多年生枝条芽的萌发。根蘖苗形成后能利用早春林分环境中充足的光照、水分和营养进行光合作用,提前进行了碳积累而迅速的生长,根蘖苗与其母株和其它物种在发育进程上的时间差为其迅速生长赢得了时间,从而增强了其竞争力,为种群的扩张创造了有利条件。
1.张川红,郑勇奇,李继磊,等.北京地区火炬树的萌蘖繁殖扩散[J].生态学报,2005,25(5):978-985.
2.蒋高明.绿色奥运要警惕外来物种入侵[J].生命世界,2004(6):6.
3.李传文,逄宗润,陈勇.火炬树——一个值得警惕的危险外来树种[J].中国水土保持,2004(2):31-38.
4.侯玉平,柳林,王信,等.外来植物火炬树水浸液对土壤微生态系统的化感作用[J].生态学报,2013,33(13):4041-4049.
5.陆霞梅,周长芳,安树青,等.植物的表性可塑性、异速生长及其入侵能力[J].生态学杂志,2007,26(9):1438-1444.
6.王长宝,杨洪升,徐增奇,等.入侵种火炬树的生物学特性及推广运用[J].安徽农业科学,2013,41(12):5402-5403.
7.喻晓丽,邸雪颖,宋丽萍.水分胁迫对火炬树幼苗生长和生理特性的影响[J].林业科学,2007,43(11):57-61.
8.安昕,孟鹏,廖国进.沈阳市降水序列趋势变化及周期分析[J].中国农学通报,2004,30(26):184-189.
9.周立彪,闫兴富,杜茜.不同火炬树种群的克隆分株能力比较研究[J].北方园艺,2009(9):170-172.
10.郑亚琼,周正立,李志军.灰叶胡杨横走侧根空间分布与克隆繁殖的关系[J].生态学杂志,2013,32(10):2641-2646.
11.曹振岭,佟亚辉,孙蕾,等.火炬树[J].特种经济动植物,2003,(6):34-35.
12.张明如,翟明普,贾黎明,等.火炬树克隆植株生长和生物量特征的研究[J].林业科学,2004,40(3):38-45.
13.王洪义,王正文,李凌浩,等.不同生境中克隆植物的繁殖倾向[J].生态学杂志,2005,24(6):670-676.
14.Laura L Jaquish,Frank W Ewers.Seasonal conductivity and embolism in the roots and stems of two clonal ring-porous trees,Sassafrasalbidum(Lauraceae) andRhustyphina(Anacardiace)[J].American Journal of Botany,2001,88(2):206-212.
15.罗维成,曾凡江,刘波,等.疏叶骆驼刺母株与子株间的水分整合[J].植物生态学报,2013,37(2):164-172.
16.张小民,王岚,林美珍,等.先锋植物沙棘Ⅱ——克隆行为和根系特化是实现沙棘先锋作用的又一根本因素[J].国际沙棘研究与开发,2006,4(2):1-5.
17.李志军,焦培培,周正立,等.胡杨横走侧根及不定芽发生的形态解剖学研究[J].北京业大学学报,2011,33(5):42-48.
18.李志军,焦培培,周正立,等.灰叶胡杨根蘖繁殖的形态解剖学特征[J].植物学报,2012,47(2):133-140.
19.董鸣,于飞海.克隆植物生态学术语和概念[J].植物生态学报,2007,31(4):681-694.
20.邹元春,吕宪国,姜明.湿地克隆植物根茎对变境适应的表型可塑性[J].湿地科学,2007,5(4):305-310.
21.马淑英,胡正海.火炬树分泌道的发育解剖学研究[J].西北植物学报,1997,17(5):112-117.
22.史宏勇,周亚福,郭建胜,等.臭椿茎中分泌道的发育及其组织化学研究[J].西北植物学报,2011,31(7):1291-1296.
23.闫兴富,周云峰,杜茜.火炬树根和凋落叶水浸提液对小麦种子萌发的化感作用[J].种子,2011,30(5):17-20.
24.白丽荣,时丽冉,徐振华,等.火炬树浸提液对几种农作物的化感作用[J].种子,2010,29(6):91-93.
25.郭阿君,王志英,邹丽.火炬树挥发性有机物释放动态及其抑菌作用[J].东北林业大学学报,2010,38(7):103-104.
26.王睿芳,冯玉龙.叶物候、构建消耗和偿还时间对入侵植物碳积累的影响[J].生态学报,2009,5(29):2568-2577.
27.Bazzaz F A.Plants in changing environments:linking physiological,population,and community ecology[J].Endeavour:Cambridge University Press,1997,21(2):92-93.
28.Kikuzawa K.The basis for variation in leaf longevity of plants[J].Vegetatio,1995,121:89-100.
29.Xu C Y,Griffin K L.Leaf phenology and seasonal variation of photo synthesis of invasiveBerberisthunbergii(Japanese barberry) and two co2 occurring native understory shrubs in a north eastern United States deciduous forest[J].Oecologia,2007,154:11-21.
30.Küppers M.Carbon relations and competition between woody species in a central European hedgerowⅠ[J].Photosynthetic characteristics Oecologia,1984,64:332-343.
CharacteristicsoftheFormationandMorphologicalAnatomyoftheAdventitiousBudsontheHorizontalLateralRootofRhustyphina
XU Yu-Feng1XU Dan-Yang2GUO Wen-Zheng1SHAO Mei-Ni1GUAN Ping1QU Bo1*
(1.College of Biological Science and Technology,Shenyang Agricultural University,Shenyang 110866;2.Faculty of Forestry,Shenyang Agricultural University,Shenyang 110866)
Rhustyphina, as a typical kind of woody plant, is with the strong abilities of adaptation and root sucker in invasion. We studied the distribution of horizontal lateral roots and adventitious buds ofR.typhinaby field investigation, anatomic characteristics of the occurrence of adventitious buds and conventional paraffin method, and observed the development of clonal seedling ofR.typhinain phenology. The distribution of the horizontal lateral roots ofR.typhinais shallow, within 15 cm below the surface, and adventitious buds mainly distributes on the horizontal lateral roots within 2-5 cm soil layer. Adventitious buds germinate throughout the growing season, but there are more buds in spring than those in other three seasons by the field observation. Adventitious buds originate from pericycle cells of horizontal lateral root. Clonal seedlings germinate earlier than their mother trees, they grow rapidly under enough sun light in the forest, which enhanced their competitive strength, provide a good condition for population regeneration and expansion, and promote its invasion.
Rhustyphina;clone growth;root sprout;anatomy;invade
国家水体污染控制与治理科技重大专项(2012ZX07202-004-01);沈阳市科技创新专项资金——大型仪器共享服务建设专项(F13-306-4-00)
许玉凤(1970—),女,副教授,硕士研究生导师,主要从事植物分类及生理生态学研究。
* 通信作者:E-mail:syau_qb@163.com
2015-11-10
S68
A
10.7525/j.issn.1673-5102.2016.03.005
