甘肃中西部地区红砂种群结构及空间格局特征
2016-10-26段桂芳单立山段雅楠
段桂芳, 单立山, 李 毅, 段雅楠, 张 荣
(甘肃农业大学 林学院, 兰州 730070)
甘肃中西部地区红砂种群结构及空间格局特征
段桂芳, 单立山, 李 毅, 段雅楠, 张 荣
(甘肃农业大学 林学院, 兰州 730070)
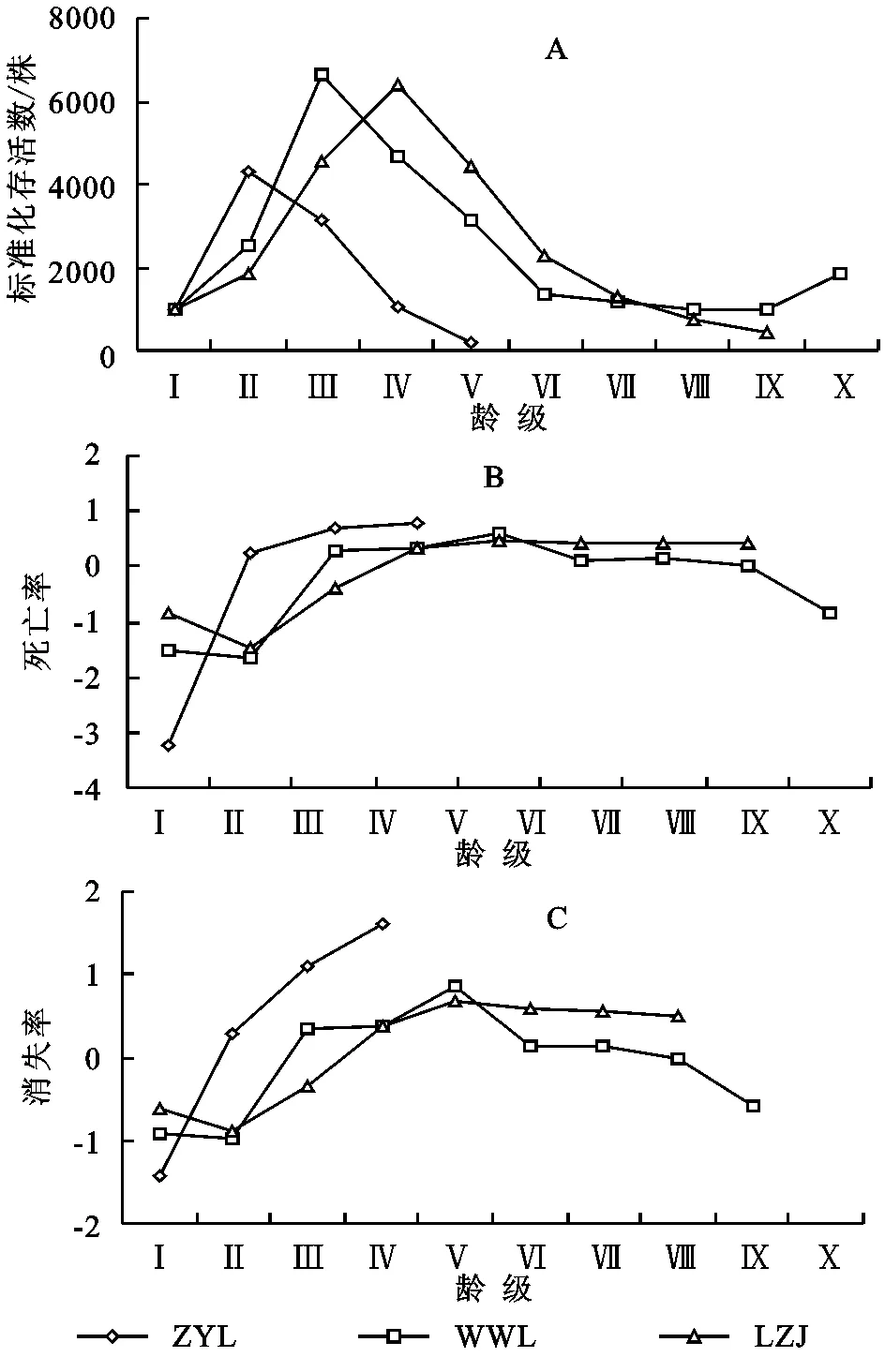
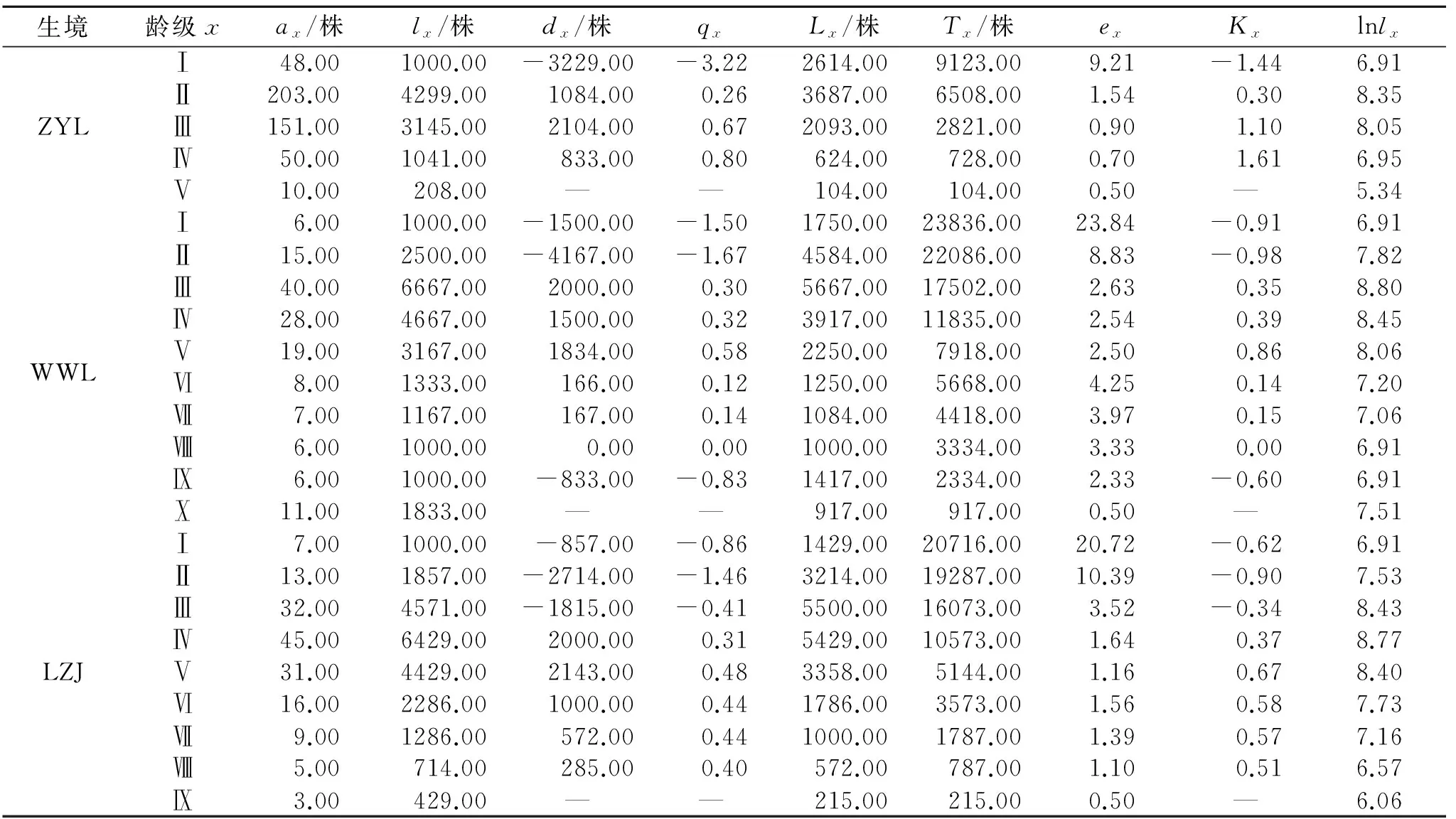
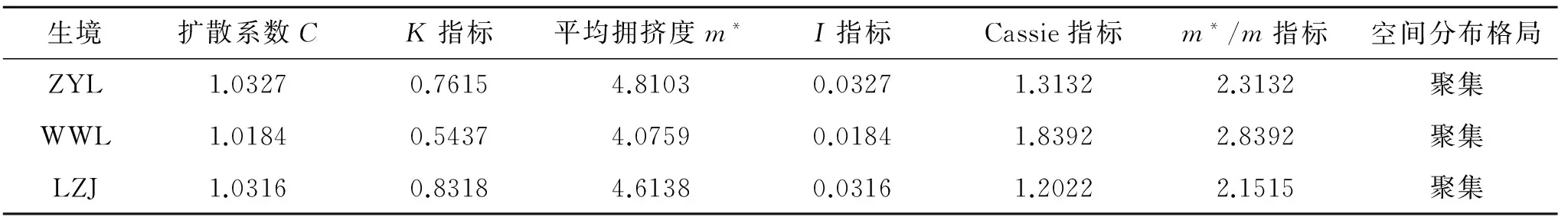
为了揭示甘肃中西部生态脆弱区红砂种群的分布特点,结合冠幅结构、静态生命表和存活曲线,对甘肃中西部地区3种典型生境条件下红砂种群年龄结构和动态进行了研究,并应用理论分布模型和聚集强度指数进行了种群分布格局分析。结果显示:在不同生境条件下红砂种群结构差异明显,山前戈壁红砂种群Ⅰ(0 cm 年龄结构; 静态生命表; 存活曲线; 时间序列预测; 空间分布 种群结构及数量动态一直是种群生态学研究的热点问题[1-2]。种群结构不仅反映了种群不同个体的配置情况,也反映了种群数量动态及其发展趋势,并在很大程度上体现了种群与环境之间的相互关系以及种群在群落中的作用和地位[3-6]。生命表和存活曲线是研究种群结构及动态变化的重要工具[7-8],它能直观展现种群各龄级的实际生存个体数、死亡数及存活趋势[9-10],结合时间序列分析能有效地预测同龄级种群未来变化的波动状态[8]。种群空间分布格局是指种群个体在水平空间上的配置状况或分布状况,是由种群特性、种群关系和环境条件综合决定的,是种群与环境长期适应与选择的结果。它能够反映出种内种间关系、空间异质性、环境适应机制等,有助于进一步加深对种群生态学的理解[3,11]。因此,通过植物种群年龄结构和空间分布格局的研究,不仅可以揭示种群的动态变化及其成因,同时对阐明生态系统的形成与维持、群落的稳定性与演替规律、种群的动态特征、种群与环境之间的互作过程以及种群更新具有极为重要的意义[12-14]。 红砂(Reaumuriasoongorica)是一种多年生的、能耐极度干旱的半灌木,在我国从东到西都有分布,特别广布于西北干旱半干旱区,是该区域的建群种和优势种[15]。因其抗逆性强,生态可塑性大,是该区重要的生物屏障,对维护荒漠地区生态环境有着重要作用。然而,红砂分布多为生态脆弱地带,长期受自然和人类活动的影响,致使其分布面积缩小,种群数量减少,在更新上产生断层,极大地影响了该区生态系统的稳定性。关于红砂的地理分布[16-17]、抗旱生理生态特性[18-21]已有大量报道,就某一生境下红砂的种群结构和分布格局也有研究[22-24],然而,在一个较大尺度范围内针对生态脆弱区不同生境下红砂种群结构特征的研究尚未见报道。本研究在一个较大尺度的自然地域上,对甘肃中西部生态脆弱地区不同生境下红砂种群结构特征和空间分布格局进行调查,探讨红砂的种群分布和动态规律,为更好地保护、合理利用生态脆弱地带红砂种群提供理论依据。 1.1研究区概况 选取3个以红砂为主要建群种的典型分布区——甘肃省张掖市龙首山山前戈壁(简称ZYL)、武威老虎口荒漠绿洲过渡带(简称WWL)以及兰州九州台黄土丘陵沟壑区(简称LZJ)作为研究区域。其中兰州九州台黄土丘陵沟壑区地处选择点最东端,该区属典型的黄土峁阶地;张掖龙首山山前戈壁地处选择点最西端,该区属大陆性荒漠草原气候;武威老虎口荒漠绿洲过渡带地处甘肃省河西走廊东北部,属典型的温带大陆性荒漠气候。各研究区自然概况、群落特征如表1—2所示。 表1不同生境的环境概况 生境经度纬度海拔/m年均降水/mm年均气温/℃0—100cm土壤含水量/%土壤容重/(g·cm-3)生境特点ZYL100°15'E39°22'N1685118.47.02.84±0.531.45±0.03砂砾质地戈壁WWL102°58'E38°44'E1378113.27.63.05±0.041.38±0.02平缓沙质地LZJ103°42'E36°57'E1432349.98.94.11±0.191.16±0.01黄土峁阶地 表2不同红砂种群研究区群落特征 生境群落组成种群平均株高/cm种群平均冠幅/m2ZYL红砂+珍珠群落35.08±3.480.20±0.02WWL红砂+白刺+沙拐枣群落39.21±5.740.87±0.09LZJ红砂+柠条+柽柳群落37.33±5.010.45±0.04 1.2研究方法 1.2.1调查方法分别选取张掖市龙首山山前戈壁、武威老虎口荒漠绿洲过渡带、兰州九州台黄土丘陵沟壑区3个天然红砂分布典型地段,在每个典型地段分别设置20 m×20 m的标准样地。把每个20 m×20 m的大样方划分为100个2 m×2 m的样方,采用相邻格子法在样方内测量每一株红砂的高度,用十字交叉法测量冠幅大小,并且准确定位每一株红砂在以样方某一顶点为圆心的相对坐标位置(x,y)[25],同时记录其他物种的种名、株高、盖度、株数等。 1.2.2径级划分通常乔木的年龄用年轮来确定。近年来,以大小级结构代替年龄结构的研究方法也被广泛应用于种群动态的研究中[26]。也有研究表明梭梭个体大小与年龄呈直线关系[27]。而红砂属于西北干旱半干旱区较为常见的一种超旱生灌木,没有明显的主干且分枝较多,同时年轮不易辨别而且存在假年轮现象,运用传统的径级研究或生长锥的方法测定其年龄不但会对植株造成伤害而且难度很大。李昌龙等[23]研究发现灌木的年龄差异主要表现在冠幅方面而在高度上表现不明显。所以本文采用王继和等[25]用冠幅梯度指示荒漠灌木植物年龄的研究方法,即用椭圆面积的计算公式C=πxy/4(其中C是椭圆面积,x和y分别是十字交叉法所测的冠幅大小轴)来计算红砂的冠幅,用冠幅梯度指示其年龄。以冠幅直径d作为划分龄级的依据,即Ⅰ级为0 cm 1.2.3种群动态数量化方法根据陈晓德[28]种群动态数量化方法,定量描述种群动态: (1) (2) 式中:Vn——种群从n到n+1级的个体数量变化;Sn,Sn+1——第n级、第n+1级种群个体数;Vpi——整个种群结构的数量变化动态指数;k——种群大小级数量。公式(2)仅适用于不考虑未来外部环境干扰的种群结构动态的比较,当考虑未来的外部干扰时,则种群结构动态还与大小级数量(k)及各大小级个体数(S)两因素相关,因此将公式(2)修正为: (3) 1.2.4种群静态生命表编制方法静态生命表可以直观明确地显示种群内不同龄级的实际存活个体数、各龄级的死亡率、种群亏损率和种群期望寿命等指数,反映了种群在这一时期内的动态变化规律,是统计种群死亡过程的一种有效的手段[29]。本文根据不同生境红砂种群不同龄级个体数,编制静态生命表,采用空间代替时间的方法,即各级冠幅对应的是各龄级,统计各龄级株数,编制种群生命表,进而分析其动态变化[30]。1.2.5种群存活率及死亡率根据红砂种群静态生命表,以龄级为横坐标,分别以存活量及死亡率为纵坐标,绘制种群存活曲线和死亡率曲线。这两条曲线是特定年龄存活率和死亡率对年龄的相关曲线,曲线的走势反映了生存率和死亡率随年龄的变化状况[31]。 1.2.6种群数量动态的时间序列分析本文采用时间序列分析的一次移动平均法[32]对红砂种群龄级结构进行模拟和预测。 (4) 式中:n——需要预测的未来时间年限;t——龄级,是近期n个观测值在t时刻的平均值,称为第n周期的移动平均,即表示未来n年时t龄级的种群大小;Xk——当前k龄级的种群大小。本文t分别取种群经历2,4,6,8 a的时间段,对种群未来发展趋势进行预测分析。 1.2.7种群空间分布格局种群空间分布格局研究采用相邻格子样方统计数据,应用离散分布理论拟合,进行方差(V)/均值(m)(亦称扩散系数,C)的t检验,以扩散系数(C)、负二项指数(K)、平均拥挤度(m*)、聚块性指标(m*/m)、丛生指数(I)、Cassie指标(CA)来进行种群集聚强度的测定[30]。主要采取以下方法:扩散系数(C),C=1时为随机分布,C>1时为集群分布,C<1时为均匀分布;负二项指数(K),K值越小聚集程度越大,如果K值无限大则分布趋于泊松分布;平均拥挤度(m*),m*值越大拥挤程度越大;聚块性指标(m*/m),m*/m=1时为随机分布,m*/m<1时为均匀分布,m*/m>1时为聚集分布,值越大聚集强度越大;丛生指数(I),I=0时为随机分布,I>0时为聚集分布,值越大聚集强度越大,I<0时则为均匀分布,值越小强度越大;Cassie指标(CA),CA=0为随机分布,CA>0为聚集分布,CA<0为均匀分布。根据6个分布类型指数判定同一生境红砂种群的空间格局分布类型。 2.1年龄结构特征 由图1可以看出,不同生境条件下红砂种群的龄级结构有所差异。张掖龙首山山前隔壁红砂种群年龄结构不完整,大于Ⅴ龄级的个体缺失,种群结构图呈非典型的“J”字型,Ⅰ,Ⅱ和Ⅲ幼龄级储量丰富,其个体数占所有径级个体总数的87.01%,Ⅳ,Ⅴ中龄级个体也达到了12.99%,种群属于增长型种群。武威老虎口荒漠绿洲过渡带和兰州九州台黄土梁峁坡地红砂种群年龄结构较为完整,各龄级都有分布,且中龄级红砂植株最为丰富,种群年龄结构符合钟型结构,种群属于稳定型。经统计,这两个区域红砂种群在Ⅱ—Ⅵ龄级个体数比较丰富,分别占整个种群个体数的75.34%和85.09%,Ⅰ龄级个体数均相对较少,仅占整个种群个体数的4.11%和4.35%;大于Ⅵ龄级个体数存在一定数量,分别占整个种群个体数的20.55%和10.56%。从整体来看,各生境条件下红砂种群中Ⅰ龄级个体数目均少于Ⅱ,Ⅲ龄级的个体数目,调查中也发现各生境条件下特别是以种子萌发的幼龄级个体的补充较少,一定程度上说明该种群以有性繁殖实现自然更新的能力相对较差。 根据种群动态量化方法分析红砂种群相邻大小级个体变化,以对种群动态进行更加客观精确地评价。由表3可以看出,不同生境条件下红砂种群相邻各级间个体数量变化动态指数为V1均小于0,说明各生境条件下Ⅰ龄级个体数目少于Ⅱ龄级个体数目。在不考虑外界的环境干扰时,各生境条件下红砂种群大小级结构的动态指数 >0,受随机干扰时的种群年龄结构动态指数 >0,但趋于零。 2.2红砂种群静态生命表及存活曲线 静态生命表在一定程度上反映了植物种群的历史累积和未来发展。由表4可以看出,不同生境条件下红砂种群第Ⅰ级个体数量均少于第Ⅱ级,出现死亡率为负的现象,说明该种群在各生境条件下幼苗补充量较少(特别是以种子萌发幼苗),自然更新受到阻碍,如若这一情况长期存在,种群将由于没有足够的幼苗输入而走向衰亡。然而,不同生境条件下红砂种群各龄级间的数量变化及死亡与成活率变化又有所不同,山前戈壁红砂种群死亡率呈增加趋势,且红砂种群在第Ⅰ级向第Ⅱ级转化过程中,种群数量波动较大,这说明在该生境条件下红砂幼苗对于环境的要求较高,受到环境的筛选存活下来的个体较少;而在荒漠绿洲过度带和黄土丘陵沟壑区红砂种群死亡率呈先增加后减少的变化趋势,各龄级在数量转化过程中相对平稳,我们认为是这两处土壤含水量相对山前戈壁优越造成这一现象(表1)。 图1不同生境条件下红砂种群年龄结构 表3不同生境条件下红砂种群龄级结构的动态变化指数 生境参数数值ZYL种群动态指数级V1V2V3V4VpiV'pi动态指数值/%-76.3525.6266.898033.850.68WWL种群动态指数级V1V2V3V4V5V6V7V8V9VpiV'pi动态指数值/%-60-62.53032.1457.8912.514.290-45.4512.530.21LZJ种群动态指数级V1V2V3V4V5V6V7V8VpiV'pi动态指数值/%-46.15-59.38-28.8931.1148.3943.7544.4420024.720.92 种群期望寿命反映的是x龄级个体的平均生存能力,从期望寿命来看,随着龄级的增加,个体间竞争加强,不同生境条件下红砂个体衰老导致期望寿命均逐渐降低。这种趋势符合种群的生物学特征,体现了目前天然红砂种群的自然生长状况相对较稳定。 种群的存活曲线更直观地描绘了种群的死亡过程,本文以标准化存活数lx为纵坐标,以龄级为横坐标绘制了不同生境条件下红砂种群存活曲线(图2A)。按Deevey的划分,一般有3种基本类型,Ⅰ型是凸曲线,属于该型的种群绝大多数都是能活到该物种年龄,早期死亡率较低,但当活到一定生理年龄时,短期内几乎全部死亡;Ⅱ型是直线,也称对角线型,属于该型的种群各年龄的死亡率基本相同;Ⅲ型是凹曲线,早期死亡率高,一旦活到某一年龄,死亡率就较低[2]。由图2A可以看出,不同生境条件下红砂种群的存活曲线为凸形,即Deevey Ⅰ型,说明红砂种群在早期死亡率低,种群数量增长快,而到一定的时间后,种群存活数量大幅度减少,种群开始衰减,最终趋于稳定。 从图2B—2C可以看出,随龄级增大,不同生境条件下红砂种群死亡率(qx)和消失率(Kx)曲线变化趋势基本一致,结合表4可以看出,随着红砂个体的成熟,植株对水分、光照、养分的竞争越来越激烈,导致死亡率逐渐上升。且第Ⅰ或Ⅱ,Ⅲ龄级出现死亡率为负的情况,说明该种群幼苗补充量较少。 2.3种群的空间分布格局分析 种群分布格局分析是对物种生物学特性、种间关系和生境条件等因素综合利用下的种群个体水平空间配置和分布状态做出定量描述。应用聚集度指标对不同生境条件下红砂种群空间分布格局进行分析,结果表明,不同生境红砂种群C均大于1,各项指数均表明红砂种群分布格局为聚集分布,只是聚集度大小不同(表5)。对每个个体来讲,m*值的大小依赖于现有的总个体数。武威老虎口荒漠与绿洲过渡带中的红砂聚块性指数最大(m*/m=2.839 2),其次是张掖龙首山山前戈壁(m*/m=2.313 2),红砂在兰州九州台梁峁坡地中的聚块性指数最小(m*/m=2.151 5)。聚块性指数考虑了空间格局本身的性质,并不涉及密度,其值越大,集聚性越强,因此,红砂在武威老虎口风沙地中的集聚性最强,在张掖龙首山山前戈壁中的聚集性次之,在兰州九州台梁峁坡地中的聚集性最弱。从K指标来判断,兰州九州台梁峁坡地K值最大,张掖龙首山戈壁山前次之,武威老虎口风沙区K值最小,这也说明,聚集性强度大小为:武威老虎口>张掖龙首山>兰州九州台。 图2不同生境条件下红砂种群存活曲线 表4不同生境条件下红砂种群静态生命表 生境龄级xax/株lx/株dx/株qxLx/株Tx/株exKxlnlxⅠ48.001000.00-3229.00-3.222614.009123.009.21-1.446.91Ⅱ203.004299.001084.000.263687.006508.001.540.308.35ZYLⅢ151.003145.002104.000.672093.002821.000.901.108.05Ⅳ50.001041.00833.000.80624.00728.000.701.616.95Ⅴ10.00208.00——104.00104.000.50—5.34Ⅰ6.001000.00-1500.00-1.501750.0023836.0023.84-0.916.91Ⅱ15.002500.00-4167.00-1.674584.0022086.008.83-0.987.82Ⅲ40.006667.002000.000.305667.0017502.002.630.358.80Ⅳ28.004667.001500.000.323917.0011835.002.540.398.45WWLⅤ19.003167.001834.000.582250.007918.002.500.868.06Ⅵ8.001333.00166.000.121250.005668.004.250.147.20Ⅶ7.001167.00167.000.141084.004418.003.970.157.06Ⅷ6.001000.000.000.001000.003334.003.330.006.91Ⅸ6.001000.00-833.00-0.831417.002334.002.33-0.606.91Ⅹ11.001833.00——917.00917.000.50—7.51Ⅰ7.001000.00-857.00-0.861429.0020716.0020.72-0.626.91Ⅱ13.001857.00-2714.00-1.463214.0019287.0010.39-0.907.53Ⅲ32.004571.00-1815.00-0.415500.0016073.003.52-0.348.43Ⅳ45.006429.002000.000.315429.0010573.001.640.378.77LZJⅤ31.004429.002143.000.483358.005144.001.160.678.40Ⅵ16.002286.001000.000.441786.003573.001.560.587.73Ⅶ9.001286.00572.000.441000.001787.001.390.577.16Ⅷ5.00714.00285.000.40572.00787.001.100.516.57Ⅸ3.00429.00——215.00215.000.50—6.06 注:x为年龄级;ax为在x年龄级内出现的个体数;lx为在x龄级开始时标准化存活数(以1 000为基数),lx=(ax/a0)×1000;dx为从x到x+1龄级期间标准化死亡数,dx=lx-lx+1;qx为从x到x+1龄级间隔期间的死亡率,qx=dx/lx;Lx为从x到x+1龄级间隔期间平均存活的个体数,Lx=(lx+lx+1)/2;Tx为从x龄级到超过x龄级的个体总数,Tx=Lx+Lx+1+Lx+2+…,即Tx=∑Lx;ex为进入x龄级个体的生命期望,ex=Tx/lx;Kx为消失率(损失度),Kx=lnlx-lnlx+1。ZYL代表张掖龙首山;WWL代表武威老虎口;LZJ代表兰州九州台。 表5不同生境条件下红砂种群聚集指标与空间分布格局类型 生境扩散系数CK指标平均拥挤度m*I指标Cassie指标m*/m指标空间分布格局ZYL1.03270.76154.81030.03271.31322.3132聚集WWL1.01840.54374.07590.01841.83922.8392聚集LZJ1.03160.83184.61380.03161.20222.1515聚集 2.4时间序列预测 红砂种群数量动态时间序列预测以红砂种群各龄级株数为原始数据,按照一次平均推移法预测出各龄级在未来2,4,6,8个龄级时间后的个体数。从表6可以看出,未来2,4个龄级时间后各生境条件下红砂种群个体数均呈现减少趋势。但从Ⅳ龄级后各生境条件下的红砂种群个体数均呈现出增加趋势。这说明各生境条件下红砂幼苗不足,幼龄红砂种群数量将有所衰减,由于缺乏可更新的幼龄个体,如不采取适当的人工自然更新措施,红砂种群未来必然趋于衰退。然而,在未来4,6,8 a,不同生境条件下红砂种群中龄级株数呈增长趋势,种群将呈稳定增长趋势。 表6不同生境条件下红砂种群动态变化的时间序列预测 生境龄级原始数据M(1)2M(1)4M(1)6M(1)8I48Ⅱ203126ZYLⅢ151177Ⅳ50101113V1030104I6Ⅱ1511Ⅲ4028Ⅳ283422WWLV192426Ⅵ8142419Ⅶ781620Ⅷ67101816Ⅸ6671216X11981016I7Ⅱ1310Ⅲ3223Ⅳ453924LZJV313830Ⅵ16243126Ⅶ9132524Ⅷ57152320Ⅸ3481820 3.1种群结构及动态 红砂作为我国西北生态脆弱区的建群种和优势种,其分布范围较广,生境类型多样。由于环境条件的差异以及种群发育历史的不同,不同生境条件下红砂种群特征存在较大差异。本研究发现,张掖龙首山山前戈壁红砂植株矮小,大于Ⅴ龄级的个体缺失,种群密度较大,这与吕朝燕等[3]对准噶尔盆地西北缘山前戈壁梭梭的研究结果一致,这可能与种群所处生境自然条件恶劣,种群整体长势较差以及种群发育历史较短有关。然而,在武威老虎口荒漠绿洲过渡带以及兰州九州台梁峁坡地不同龄级红砂个体均占一定的比例,不存在径级缺失现象,可见,该两种生境红砂种 群发育时间较长,种群处于稳定发展阶段。同时,研究发现各生境条件下红砂Ⅰ龄级个体所占比例较少(4.11%~10.39%),这说明红砂种群缺少幼苗(特别是以种子萌发的幼苗)输入,种群更新可能存在问题,种群已经有衰退的倾向。 从静态生命表和存活曲线来看,不同生境条件下红砂种群的存活曲线为凸形即DeeveyⅠ型,说明红砂种群在早期死亡率低,种群数量增长快,而到一定的时间后,种群开始衰减,最终趋于稳定。在种群静态生命表的编制过程中,由于是用同一时期收集的种群内部全部个体的数据编制而成,是对种群发展历程的一段特定时间的反映,而不是对整个种群生活史进行探讨分析[3],且在野外调查中系统抽样误差不可避免,因此在生命表中会出现负死亡率的情况。对于这一问题学者们的处理意见分为两派,有些学者认为应对该龄级数据波动进行匀滑处理[31];如林勇明等[32]对野生桂花种群研究时采用匀滑(Smooth out)技术进行处理,对ax进行匀滑修正,据此编制桂花种群静态生命表;李清河等[33]对荒漠珍稀灌木半日花种群静态生命表编制时也对其进行匀滑处理;保长虎等[34]在编制黄土丘陵沟壑区柠条种群静态生命表的过程中,也采取了匀滑技术。但是有些学者认为运用匀滑技术会隐藏某些生态学现象,如宋于洋等[35]在对石河子不同生境条件下的梭梭种群进行研究时没有对相关数据进行匀滑处理,将各龄级株数标准化后直接用于各参数计算效果良好;刘国军等[36]对准噶尔盆地东南缘梭梭种群进行调查时也未对原始数据进行匀滑处理。本研究并未对数据做任何匀滑处理,利用原始数据进行分析研究,以保证更加客观真实地反映种群的生态学现象。静态生命表、存活曲线均反映出不同生境红砂种群虽然现状稳定,但是种群内部幼苗数量储备不足,长此以往若是种群幼苗得不到及时补充,种群将会衰退,这与种群年龄结构研究结果相吻合。各生境条件下红砂种群表现为缺乏幼苗特别是以种子萌发幼苗缺失,这与其自身有性更新能力相对于无性生殖能力较弱、幼苗难以存活有密切的联系。可以预测,在种群发展若干年后,目前处于中龄级范围(70%~90%)的红砂个体将全部老龄化,如果没有足够的幼龄植株进行补充,势必影响种群的整体稳定性。 3.2种群空间分布格局 种群分布格局是指植物在环境、资源及群落内其他物种的影响下在空间上表现出来的具有一定规律的分布情况,受时间限制的同时又随着时间发生着变化[37]。研究种群分布格局的目的不仅是对种群的水平结构特征进行定量化的描述[38],更重要的是用以揭示格局的成因,阐述种群及群落的动态变化等,为种群保护措施的制定提供一定的科学依据[4]。本研究发现,不同生境条件下红砂种群整体的分布格局表现为聚集分布,这与何志斌等[22]对黑河中游甘肃省临泽县北部荒漠绿洲过渡带红砂种群的分布格局呈现出聚集分布的观点一致,同时与李昌龙等[23]的研究结果吻合。这可能是红砂自身的物种特性以及与当地的生态环境密不可分;红砂具有很强的无性繁殖能力,尤其在相对干旱的环境条件下,尤以无性繁殖为主。再者,聚集分布对于恶劣的环境抵抗能力较强,能很好地抵御风沙较大、降水少的干旱荒漠气候。 研究发现虽不同生境条件下红砂种群均表现为聚集分布,但聚集强度不同,表现为武威老虎口>张掖龙首山>兰州九州台。种群空间分布的格局强度是物种自身的特性与环境长期耦合作用的结果。研究发现,光照与土壤条件在不同生境间的异质性分布是克隆植物具有差异性的主要原因[39]。光照的增强有利于克隆植物产生更多的克隆子株[39],王洪义等[40]也指出在环境条件趋于严酷时,克隆植物的无性繁殖在种群更新中的相对重要性有所增加。红砂为克隆植物,在河西走廊(张掖、武威)光照资源明显优于黄土丘陵沟壑区(兰州九州台),且风沙频繁自然环境较恶劣,因此,河西走廊红砂种群聚集强度大于黄土丘陵梁峁坡地种群。总之,红砂种群表现为聚集分布的特点,一方面主要取决于红砂无性繁殖的生物学特性,同时又与该地区的水文水资源、气候、土壤和地形等异质性环境密切相关;另一方面也是红砂种群对异质性环境的具体响应[41]。 3.3甘肃中西部生态脆弱区红砂适应性保护和恢复对策 种群的更新是种群生态学关注的焦点。种群能否实现更新,是种群能否长久存在与发展的关键[3]。本研究发现,甘肃中西部不同生境条件下红砂幼苗较少特别是以种子萌发的幼苗几乎缺失,表明幼苗更新断层。如果这一情况长期存在并发展下去,红砂种群将在年龄上表现出断代现象,同时种群的扩张能力会弱化。因此,可以判定在甘肃中西部生态脆弱区红砂以种子开始的自然更新存在问题,如果不加以关注,随时间的推移,该种群会逐渐走向衰退直至灭亡。红砂在我国西北生态脆弱区分布,是这些自然分布带的建群种和优势种,因其抗逆性强,生态可塑性大,是该区重要的生物屏障,对维护荒漠地区生态环境有着重要作用。红砂能够通过无性繁殖形成沙堆效应并适应该地区干旱气候和瘠薄的土壤环境,进而成为荒漠地区的重要植被类型,说明红砂是西北干旱半干旱区适应力极强的超旱生灌木。然而,由于红砂分布地区多为生态脆弱地带,长期受自然和人类活动的影响,致使其分布面积缩小,种群数量减少,且以种子开始的自然更新存在断层,极大地影响了干旱生态系统的稳定性。因此,一方面,对于处于稳定发展阶段的红砂种群进行封育加以保护,避免放牧及人为干扰,滥采等不合理的利用,维持当地生态系统的稳定。另一方面,由于以种子开始的幼苗更新出现断层,然而红砂种子结实率多,因此在种子成熟期适时采收种子,在洪水季节引洪漫灌,提高自然条件下种子向幼苗的转化率,在人工辅助下实现以种子开始的自然更新过程。 [1]刘振学,任广鑫,王得祥,等.通天河国家森林公园锐齿栎群落物种组成及群落结构研究[J].水土保持研究,2011,18(2):86-93. [2]卢杰,郭其强,郑维列,等.藏东南高山松种群结构及动态特征[J].林业科学,2013,49(8):154-160. [3]吕朝燕,张希明,刘国军,等.准噶尔盆地西北缘梭梭种群结构和空间格局特征[J].中国沙漠,2012,32(2):380-387. [4]赵丽,王晓江,刘果厚,等.浑善达克沙地榆树种群结构、格局及动态研究[J].中国沙漠,2009,29(3):508-513. [5]李吉玫,徐海量,张青青,等.塔里木河下游荒漠河岸林不同退化区胡杨种群结构和空间分布格局研究[J].中国沙漠,2009,29(5):897-904. [6]李海东,沈渭寿,佘光辉,等.西藏砂生槐种群结构与点格局分析[J].中国沙漠,2011,31(6):1443-1448. [7]Harcombe P A. Tree life tables [J]. Bioscience,1987,37(8):557-568. [8]王卓,黄荣凤,王林和,等.毛乌素沙地天然臭柏种群生命表分析[J].中国沙漠,2009,29(1):118-124. [9]Díaz S, Mercado C, Alvarez-Cardenas S. Structure and population dynamics ofPinuslagunaeM.-F. Passini[J]. Forest Ecology and Management, 2000,134(1):249-256. [10]洪伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报.2004,15(6):1109-1112. [11]奇凯,张春雨,侯继华,等,赤峰市黑里河天然油松林空间分布格局研究[J].水土保持研究,2010,17(4):239-242. [12]徐学红,于明坚,胡正华,等.浙江古田山自然保护区甜槠种群结构与动态[J].生态学报,2005,25(3):645-653. [13]杨永川,达良俊,陈波.天童米槠—木荷群落主要树种的结构及空间格局[J].生态学报,2006,26(9):2927-2938. [14]赵哈林,苏永中,周瑞莲.我国北方沙区退化植被的恢复机理[J].中国沙漠,2006,26(3):323-328. [15]马剑英,陈发虎,夏敦胜,等.荒漠植物红砂叶片δ13C值与生理指标的关系[J].应用生态学报,2008,19(5):1166-1171. [16]潘晓玲,党荣理,伍光和.西北干旱荒漠植物区系地理与资源利用[M].北京:科学出版社,2003. [17]刘家琼,邱明新,蒲锦春,等.我国荒漠典型超旱生植物:红砂[J].植物学报,1982,24(5):485-488. [18]种培芳,李毅,苏世平,等.红砂3个地理种群的光合特性及其影响因素[J].生态学报,2010,30(4):914-922. [19]种培芳,李毅,苏世平.荒漠植物红砂叶绿素荧光参数日变化及其与环境因子的关系[J].中国沙漠,2010,30(3):539-545. [20]周海燕,谭会娟,张志山,等.红砂和珍珠对极端环境的生理响应与调节机制[J].中国沙漠,2012,32(1):24-32. [21]周生荟,刘玉冰,谭会娟,等.荒漠植物红砂在持续干旱胁迫下的光保护机制研究[J].中国沙漠,2010,30(1):69-73. [22]何志斌,赵文智.黑河流域荒漠绿洲过渡带两种优势植物种群空间格局特征[J].应用生态学报,2004,15(6):947-952. [23]李昌龙,马瑞君,王继和,等.甘肃民勤连古城自然保护区优势种种群结构和动态研究[J].西北植物学报,2005,25(8):1628-1636. [24]周资行,李毅,焦健.兰州市南北两山不同生境红砂种群数量动态研究[J].自然资源学报,2011,26(10):1726-1737. [25]王继和,靳虎甲,马全林,等.干旱区油蒿种群结构和分布格局分析[J].中国沙漠,2010,30(3):534-538. [26]宋于洋,楚光明,胡晓静.古尔班通古特沙漠梭梭种群径级与龄级关系的研究[J].西北植物学报,2011,31(4):808-814. [27]刘峰,陈伟烈,贺金生.神农架地区锐齿槲栎种群结构与更新的研究[J].植物生态学报,2000,24(4):396-401. [28]陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217. [29]周纪纶.植物种群生态学[M].北京:高等教育出版社,1992. [30]谢宗强,陈伟烈,路鹏,等.濒危植物银杉的种群统计与年龄结构[J].生态学报,1999,19(4):523-528. [31]段仁燕,黄敏毅,吴甘霖.黄山松种群结构与动态研究[J].生物学杂志,2009,26(1):34-37. [32]林勇明,洪滔,吴承祯,等.桂花野生种群生命表及生存分析[J].北京林业大学学报,2007,29(3):185-188. [33]李清河,高婷婷,刘建锋,等.荒漠珍稀灌木半日花种群的年龄结构与生命表分析[J].植物研究,2009,29(2):176-181. [34]保长虎,张文辉,何景峰.黄土高原丘陵沟壑区30年柠条人工种群动态研究[J].西北植物学报,2010,30(8):1681-1688. [35]宋于洋,刘长青,赵自玉.石河子地区不同生境梭梭种群数量动态分析[J].西北植物学报,2008,28(10):2118-2124. [36]刘国军,张希明,朱军涛,等.准噶尔盆地东南缘梭梭种群结构与动态研究[J].西北植物学报,2011,31(6):1250-1256. [37]宋萍,洪伟,吴承桢,等.珍稀濒危植物桫椤种群结构与动态研究[J].应用生态学报,2005,16(3):413-418. [38]李皓,姜景民,邵文豪,等.浙江九龙山自然保护区粉叶柿群落的结构特征[J].林业科学研究,2009,22(3):360-366. [39]罗学刚,董鸣.蛇莓克隆构型对光照强度的可塑性反映[J].植物生态学报,2001,25(4):494-497. [40]王洪义,王正文,李凌浩,等.不同生境中克隆植物的繁殖倾向[J].生态学杂志,2005,24(6):670-676. [41]郭华,王孝安,肖娅萍.秦岭太白红杉种群空间分布格局动态及分形特征研究[J].应用生态学报,2005,16(2):227-232. Population Structure and Saptial Pattern ofReaumuriasoongoricaPopulation in Midwest Region of Gansu DUAN Guifang, SHAN Lishan, LI Yi, DUAN Ya′nan, ZHANG Rong (CollegeofForestry,GansuAgriculturalUniversity,Lanzhou730070,China) In order to reveal the distribution characteristics ofReaumuriasoongoricapopulation in ecological fragile areas in midwestern Gansu, based on the crown structure and static life table and survival curve, age structure and dynamics ofReaumuriasoongoricapopulation under three typical habitat conditions in the central and western regions of Gansu were studied, besides, the theory of distribution model and aggregation intensity index were applied to analyze the population distribution pattern. The results showed thatReaumuriasoongoricapopulation structure was obviously different under different habitat conditions.Ⅰ(0 cm population structure; static life table; survival curve; time series analysis; spatial distribution 2014-11-17 2015-03-05 国家自然科学基金(41361100,31360205);国家国际科技合作专项(2012DFR30830);甘肃省科技支撑计划项目(1204NKCA084) 段桂芳(1989—),女,甘肃景泰人,硕士研究生,研究方向为西北抗旱灌木研究。E-mail:13659431519@163.com 李毅(1962—),男,甘肃兰州人,教授,主要从事荒漠植物林木遗传育种研究。E-mail:liyi@gsau.edu.cn S722 A 1005-3409(2016)01-0067-081 研究区概况与研究方法



2 结果与分析



3 讨论与结论
