伏牛山自然保护区森林生态系统植物功能群光合特性
2016-10-24范玉龙刘慧敏丁圣彦
范玉龙,刘慧敏,胡 楠 ,丁圣彦,*
1 河南大学环境与规划学院, 开封 475001 2 南阳理工学院, 南阳 473004
伏牛山自然保护区森林生态系统植物功能群光合特性
范玉龙1,2,刘慧敏1,胡楠2,丁圣彦1,*
1 河南大学环境与规划学院, 开封475001 2 南阳理工学院, 南阳473004
植物功能群(PFG)把一系列植物归为一个功能团体,可以作为植被随环境动态变化的基本单元,简化相关研究。植物功能群内不同物种的生理特性差异是功能群存在的基础,也影响着功能群整体的功能。采用种间联结法并结合优势种在海拔梯度上的变化,将伏牛山自然保护区森林生态系统各层优势种划分为4组植物功能群,研究每组植物功能群内不同物种的光合特性。对光合-光响应、最大净光合速率(Pn)、暗呼吸速率(Rd)、光补偿点(LCP)、光饱合点(LSP)和表观量子效率(AQY)等指标进行分析和比较,结果表明:植物功能群内不同物种的光补偿点和暗呼吸速率的变异系数最高分别达70.47%和74.29%,功能性状存在着显著的互补性,这是物种能长期共存的主要原因。乔木的最大净光合速率在7—8 μmol CO2m-2s-1之间,对高光强的适应能力较高。同一功能群内各层植物光饱合点在1200—1500μmol m-2s-1之间,差别不大。林下灌木、草本植物对弱光的利用效率较乔木高。同一功能群内物种功能性状的差异及不同植物功能群间整体性状的差异,影响着森林生态系统的结构、功能、过程和生态系统服务。
植物功能群(PFE);光合特性;伏牛山自然保护区;生态系统功能;生态系统服务
植物功能群(PFG)是具有确定功能特征的一系列植物的组合,可以看作是对环境有相同响应和对主要生态系统过程有相似作用的组合[1-2]。森林生态系统结构复杂,研究难度较大,植物功能群把一系列植物归为一个功能团体,使复杂的生态系统研究得以简单化,可以作为研究植被随环境动态变化的基本单元。PFG是与植物的外貌密切相关的,或多或少反映了植物在不同环境中的生存能力[3-4]。植物功能群将植物形态学、群落学和生态过程联系起来,为研究气候变化提供了一个非常好的工具[5],也为恢复生态系统提供更为科学、实用的物种组合。基于植物功能群的研究思路不仅简便,而且利于揭示生物多样性与生态系统服务的深层次机理[6]。
森林生态系统可以从结构和功能两个方面来描述,也可从结构和功能两方面进行分类[7]。优势种控制着植被的结构,能更好地体现出地区生态系统的状态,一个群落中的优势种,特别是演替顶极中的优势种,或者至少是特征种就比次要种能成为更有用的指示植物[8],以优势种为主体来划分植物功能群具有较强的代表性。植物功能群的划分依赖于植物实际的分布[9],也反映出功能相近的植物在时空分布过程中的互补性。伏牛山自然保护区森林生态系统植被可以分为3层,即乔木层、灌木层和草本层,在个别地带也存在藤本植物,但是种类和数量较少。以各层优势种为主体分别划分植物功能群(即乔木层、灌木层和草本层各自为主体划分各自的植物功能群)能反映出不同层次植物的功能特性[10-12]。随着研究尺度的扩大,将不同层次的植物划分在同一植物功能群更具实际意义,可以深化对森林生态系统的协同进化的理解,特别是对森林生态系统结构、功能和生态系统服务的研究提供思路[13],为森林生态系统立体化恢复提供参考[14-15]。
植物功能群功能的发挥依赖于群内不同植物的功能贡献,因此,功能群内植物的生理生态特性及其差异就成为此类研究的关键一环。无论何种划分方法,植物功能群研究必定要涉及到功能性状概念,植物功能性状是指影响物种生存、生长和繁殖等生物特征[16]。光是限制植物生长的最主要的生态因子,光合特性是植物最主要的生理生态特性,光合作用是能量流动的基础。在垂直结构明显的森林生态系统中,群落结构的复杂性及其对光辐射的吸收、反射和透射等作用,能强烈地改变光强、光质和光照时间[17],光对林下灌木层和草本层植物的发育就显得非常重要。伏牛山国家级自然保护区植物种类丰富,不同种类植物的形态及生理特性差异显著,表现出不同的需光特性。上层乔木接受直接光照,灌木层和草本层接受反射和透射光,光合特性的差异使它们能适应不同的光照强度,对光的充分利用使整体光合效率高于单一物种。这种功能互补正是植物功能群长期共存的生理生态基础[18]。
1 研究地概况
伏牛山自然保护区位于河南省西部,是秦岭东西方向上的余脉,地理坐标为东经110°30′—113°05′,北纬32°45′—34°00′;伏牛山区是黄河、淮河和长江三大水系的分水岭和一些支流的发源地;境内地势高差悬殊,在季风气候的背景条件下,根据中国气候区划的指标,伏牛山北坡属暖温带,南坡属北亚热带,具有明显的过渡带特点;伏牛山区的土壤复杂多样,可划分为3种土类(山地棕壤、山地黄棕壤和山地褐土);植被属暖温带落叶阔叶林向亚热带常绿阔叶林的过渡型[19]。北坡的洛阳白云山与南坡的南阳宝天曼自然环境随海拔变化差异显著,群落类型变化梯度大,优势种变化明显,为此类研究提供了一个良好场所(表1)。

表1 研究区自然地理概况
2 研究方法
在北坡的白云山和南坡的宝天曼,从海拔600—2 000 m的山体上,选择代表性地段,每隔海拔100 m确定至少1个20 m×20 m的样地,共设置样地66个,在样地的中心以及四个角分别设一个2 m×2 m灌木层样方和一个1 m×1 m草本层样方。其中包括乔木层样方66个,灌木层样方共330个,草本层样方共330个。调查指标包括:乔木层的盖度、种类、个体数量、树高、枝下高、冠高、冠幅、基径、胸径等;灌木层的盖度、种类、个体数量、树高、冠幅、基径;草本层的盖度、种类、个体数量、高度、冠幅、基径(丛径)。
由于各物种所处的层次不同,优势种确定所采用的方法也不同,乔木层是以重要值为指标,灌木和草本层以出现次数为指标。植物功能群划分采用优势种种间联结法[20],并结合其在海拔梯度上的变化[10-12]。这种划分方法的客观性在于结合了森林生态系统的结构、功能和过程[6]。
用Li- 6400对植物功能群内优势种进行光合日变化、光响应测定,时间从8:00到18:00,在相同高度位置,选择充分伸展的枝条上的第3—7片成熟叶片进行测量,每片叶片进行3个重复,循环测定,保证每2 h左右循环测定1次。灌木层和草本层采用连体叶片,测定方法同上。光合-光响应曲线测定在10: 00后进行,3次重复, 采用人工光源, 测定前使用1 000μmolm-2s-1的光强诱导20 min, 光合有效辐射(PAR)分别设定为2 000, 1 500, 1 000, 800, 500, 200, 100, 50, 30, 20, 10, 5, 0μmolm-2s-1等13个水平。通过拟合曲线得到最大净光合速率、光补偿点、光饱和点和表观量子效率等参数[21,22]。
3 结果与分析
通过对野外调查数据的整理、计算和分析,确定了伏牛山自然保护区森林生态系统各层的主要优势种(表2)。
根据优势种种间联结及其在海拔梯度上的变化,可以将乔木、灌木和草本作为整体划分为4组植物功能群。第1组:栓皮栎、胡枝子、栓皮栎(灌木)、连翘和羊胡子草;第2组:短柄枹、胡枝子、栓皮栎(灌木)、菅草、苍术和羊胡子草;第3组:锐齿栎、华山松、大芽南蛇藤、橐吾和宽叶苔草;第4组:华山松、青榨槭、杜鹃、橐吾和宽叶苔草。
光照是植物初级生产力的能量来源,在其他条件都合适时,光强是决定光合速率高低的决定因素。在森林生态系统中,植被分层现象明显,各层植物优势种在光合特性上存在显著差异,这种差异使它们在协同进化过程中形成了稳定的组合,也使整个森林生态系统具有更强的光转化能力。
3.1第1组植物功能群光合特性
第1组植物功能群主要分布在低海拔区域,特别是在800 m以下,人为影响较大,有些地方属于正处在发育中的幼林区,群落层次变化较大,结构相对单一,处在森林生态系统的演替初期[10]。栓皮栎的表观量子效率为0.0208,是灌木层的胡枝子和连翘的表观量子效率的1/2,也低于草本层的羊胡子草,说明本组植物功能群的乔木层对弱光的利用效率远低于灌木层和草本层。栓皮栎无论是处在乔木层还是灌木层,光补偿点相对较高,且暗呼吸速率也较高,说明栓皮栎对光强的要求较高。光补偿点最低的是连翘,它能生长在林下光强较弱的地方。植物功能群内物种的光饱和点除胡枝子外基本一致,胡枝子的各项指标均较高,这是它在灌木层优势度最大的主要原因。羊胡子草的表观量子效率较高,光补偿点比连翘高,在栓皮栎稠密的地段只有少量的羊胡子草存在,在乔木长势不好或稀疏的地段成片分布。各物种光补偿点的变异系数最高,对弱光的利用能力存在显著差异。这是物种生态位的分化、适应不同生存环境和长期共存的主要原因之一(表3)。
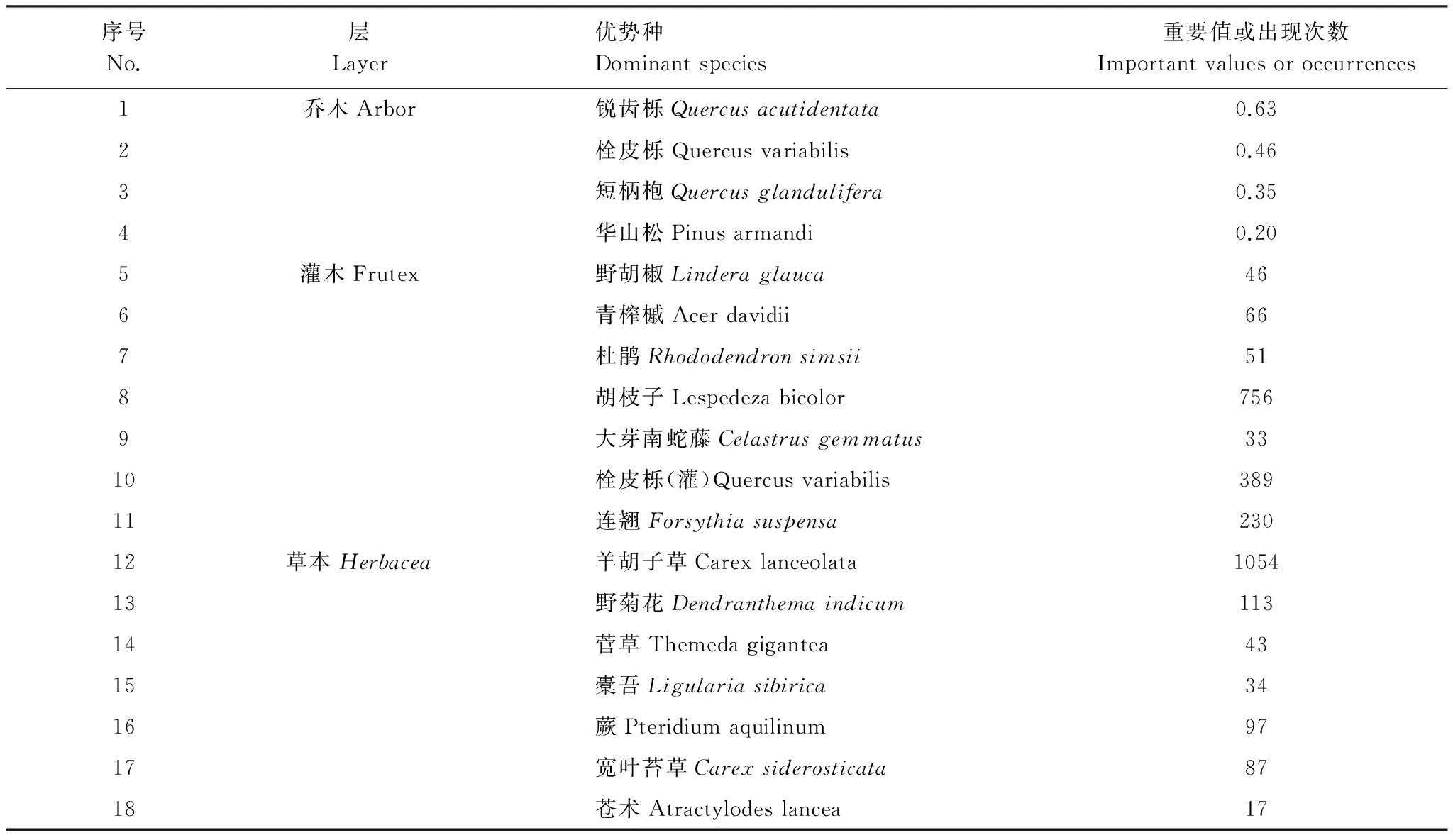
表2 研究区各层主要优势种
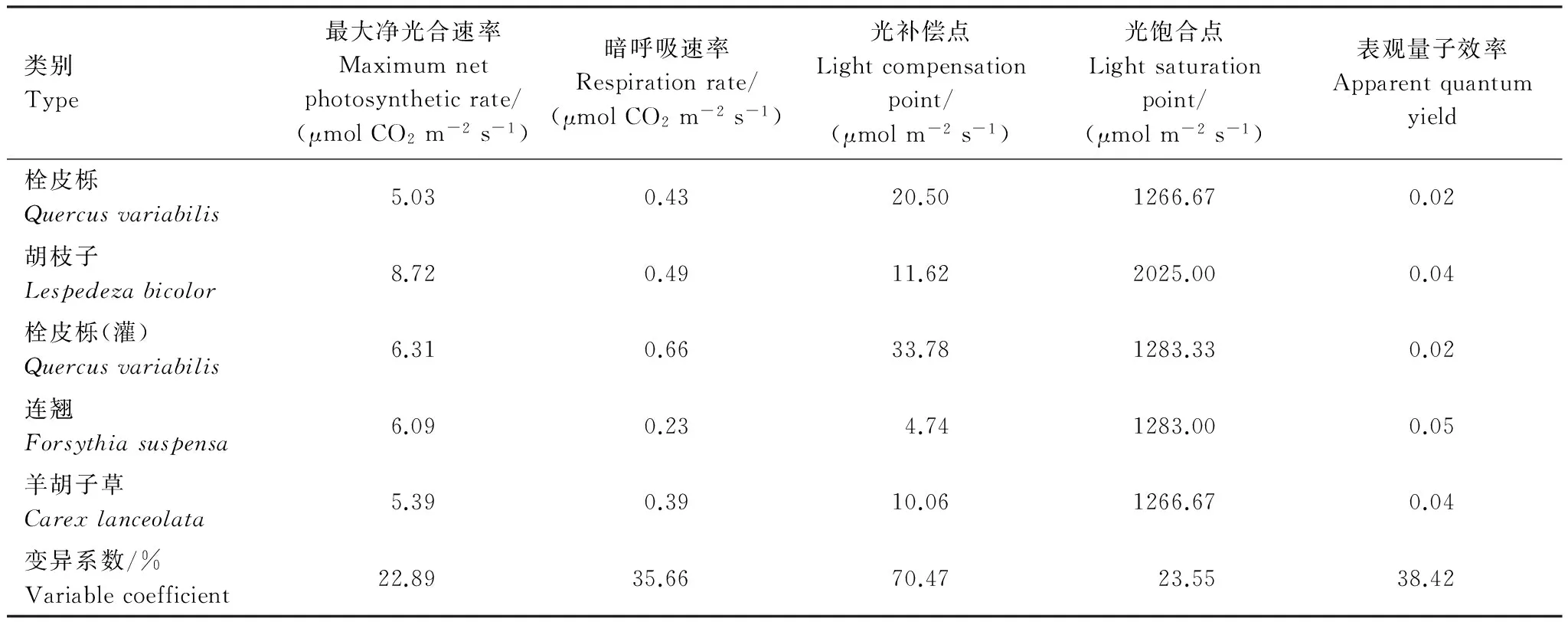
表3 第1组植物功能群光合特性
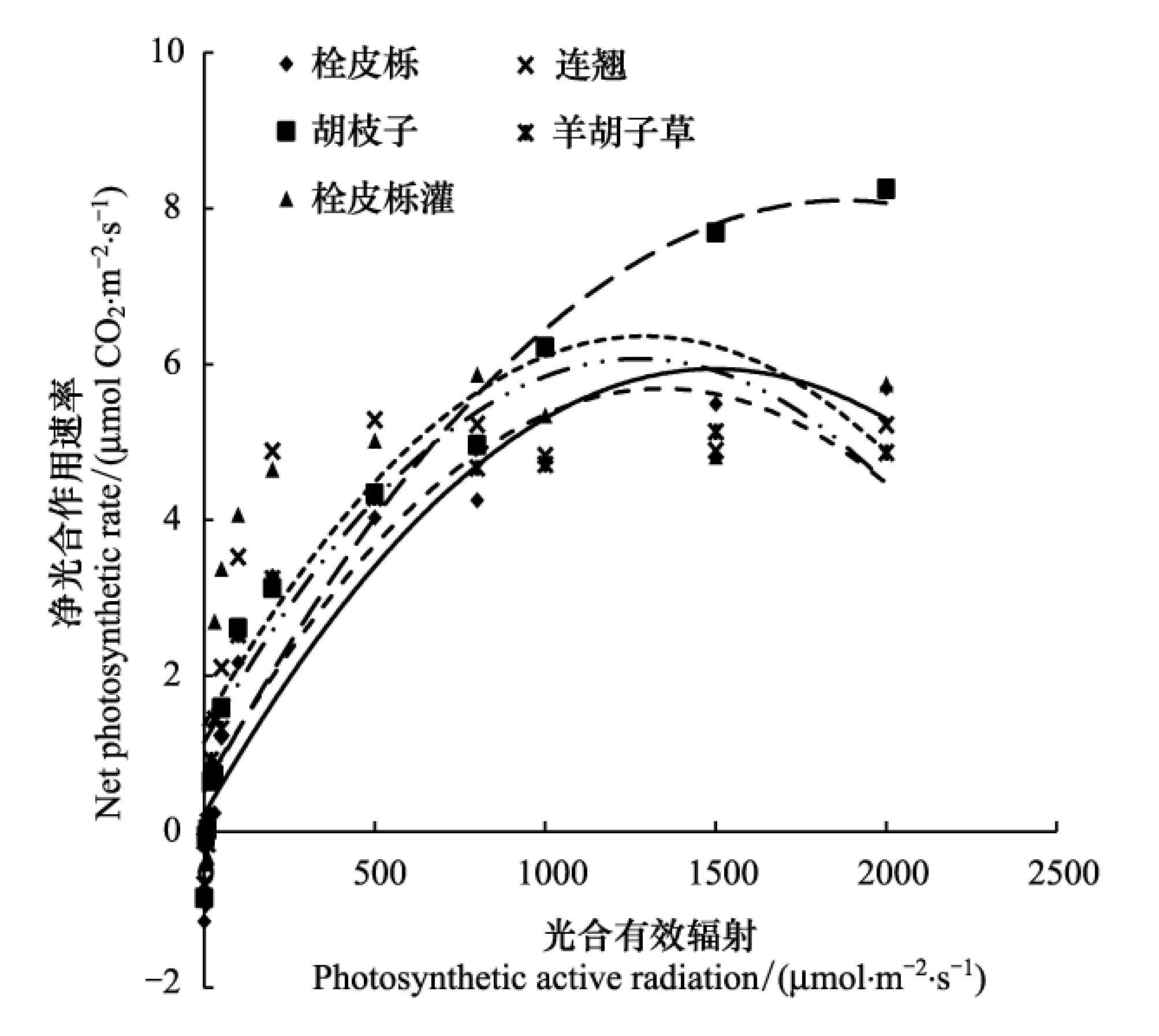
图1 第1组植物功能群光合-光响应曲线Fig.1 Photosynthesis-light response curve of the first PFG
在低光照强度下,灌木层和草本层植物的光合速率均较乔木层栓皮栎的高(图1),林下灌木和草本植物对低光强的适应性较强。在高光强条件下,乔木栓皮栎的光合速率又高于其它植物,这与其所处的上层位置相适应。胡枝子作为灌木层植物,对高光强也有较高的适应性,它一般分布在林下乔木冠层还没有闭合的地方,特别在林中的路边到处可见,其喜光的特性及对高光强的适应性使其能广泛的分布在整个伏牛山区。胡枝子可以作为良好的搭配物种在本区域的植被恢复过程中得到应用。
3.2第2组植物功能群光合特性
第2组植物功能群主要分布在海拔1 100—1 400m之间,属中低海拔到高海拔的过渡区,位于栓皮栎林与锐齿栎林的垂直过渡带,乔木层以短柄枹为建群种,栓皮栎逐渐减少,灌木层的胡枝子和栓皮栎仍是优势种[10]。短柄枹的表观量子效率和栓皮栎基本一样,但是最大光合速率达8.19μmol CO2m-2s-1,明显较栓皮栎高。菅草的表观量子效率达0.0585,在此海拔区域,分布着大量的菅草群丛,特别是在土壤比较贫瘠的地段更为显著,弥补了羊胡子草的不足。苍术的最大净光合速率为2.75μmol CO2m-2s-1,显著低于同组其他物种,它零星分布在林下 (表4)。
短柄枹是该地段的乔木层优势物种,在光照强度30 μmolm-2s-1以下,除苍术外,它的净光合速率最低,随着光强的增加,它的净光合速率逐渐超过其它物种,并且在光照强度达到1 000 μmolm-2s-1时成为光合速率最高的物种,这种光强也是通常天气条件下的光照强度。菅草和羊胡子草的光响应曲线非常接近,说明它们的光合特性接近,具有一定的可替代性,而其它条件(如土壤和水分)的差异就会影响它们分布。胡枝子的净光合速率在本组中最高,仍是灌木层的优势种(图2)。
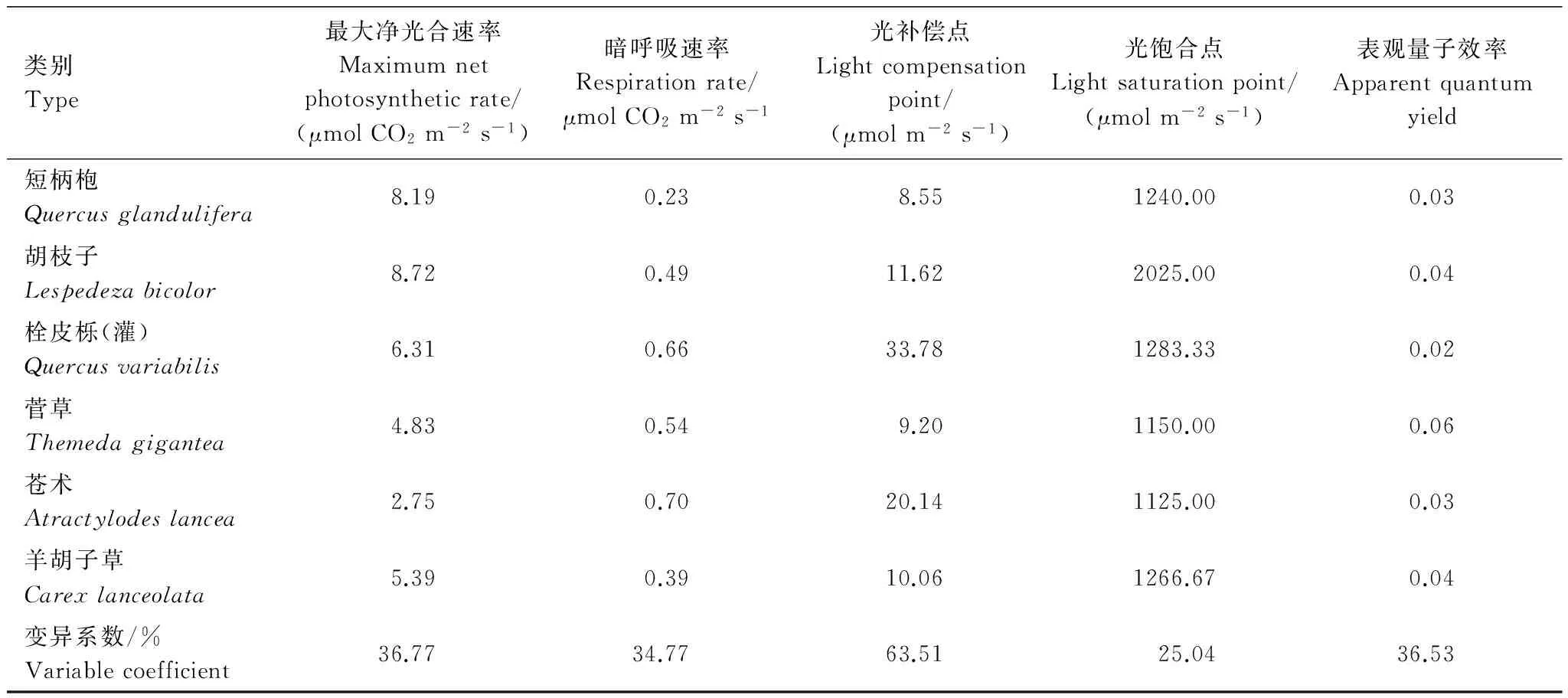
表4 第2组植物功能群光合特性
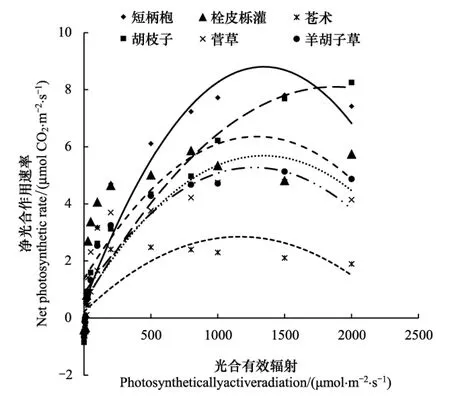
图2 第2组功植物能群光合-光响应曲线Fig.2 Photosynthesis-light response curve of the second PFG
3.3第3组植物功能群光合特性
第3组植物功能群主要分布在1 400—1 800 m的中高海拔区,以锐齿栎为建群种[10]。锐齿栎的表观量子效率为0.0243,和前两组中的乔木差距不大,与短柄枹更为接近,最大光合速率为7.63μmol CO2m-2s-1,稍低于华山松。大芽南蛇藤的净光合光合速率较低,灌木层植物长势较弱。橐吾是草本层的优势植物,表观量子效率在本组中最高,但是其暗呼吸速率较高,虽然生长旺盛,干物质积累比较缓慢,加上其光补偿点较高,一般分布在林窗或林隙等光线较强的地方。宽叶苔草最大光合速率较低,表观量子效率相对较高,暗呼吸速率也比较高,整体上其生长速度较快,但是叶片的活力周期短,难以积累太多干物质,只能作为地被物种存在(表5)。
从整个光响应曲线来看,锐齿栎和华山松有较高的重合度,它们对光的利用能力相似,叶片形状及数量的不同使它们的光合单位差异较大,影响其第一生产力,从而会影响到它们的优势度。橐吾的光合利用效率一直高于林下的其它植物,特别是在高光强的条件下更为明显,也就使它在光强较高的地段成为主要优势种,而大芽南蛇藤和宽叶苔草的净光合速率一直处在相对较低的水平(图3),限制了它们的生长,宽叶苔草在这个地段的分布较广,这是由其他生理生态特性决定的。
3.4第4组植物功能群光合特性
第4组植物功能群主要分布在1 800 m以上的高海拔区,此地带温度低、湿度大,群落稳定,分层现象明显,分布着以锐齿栎、坚桦和红桦组成山顶矮曲林群落[10]。华山松和橐吾的光补偿点、最大净光合速率都比较高,它们都具有强的喜光特性,同时,它们的暗呼吸速率比较高,代谢强度大,活力较强,比较适应这一地段的环境。受温度等条件的影响,青榨槭在这个地段一般发育成小乔木或灌木,这与它的低光补偿点相适应。杜鹃一般分布在山脊,光照条件好,它的光补偿点和光饱和点都较高,但其表观量子效率较低,生长比较缓慢,较多发育为小灌丛(表6)。
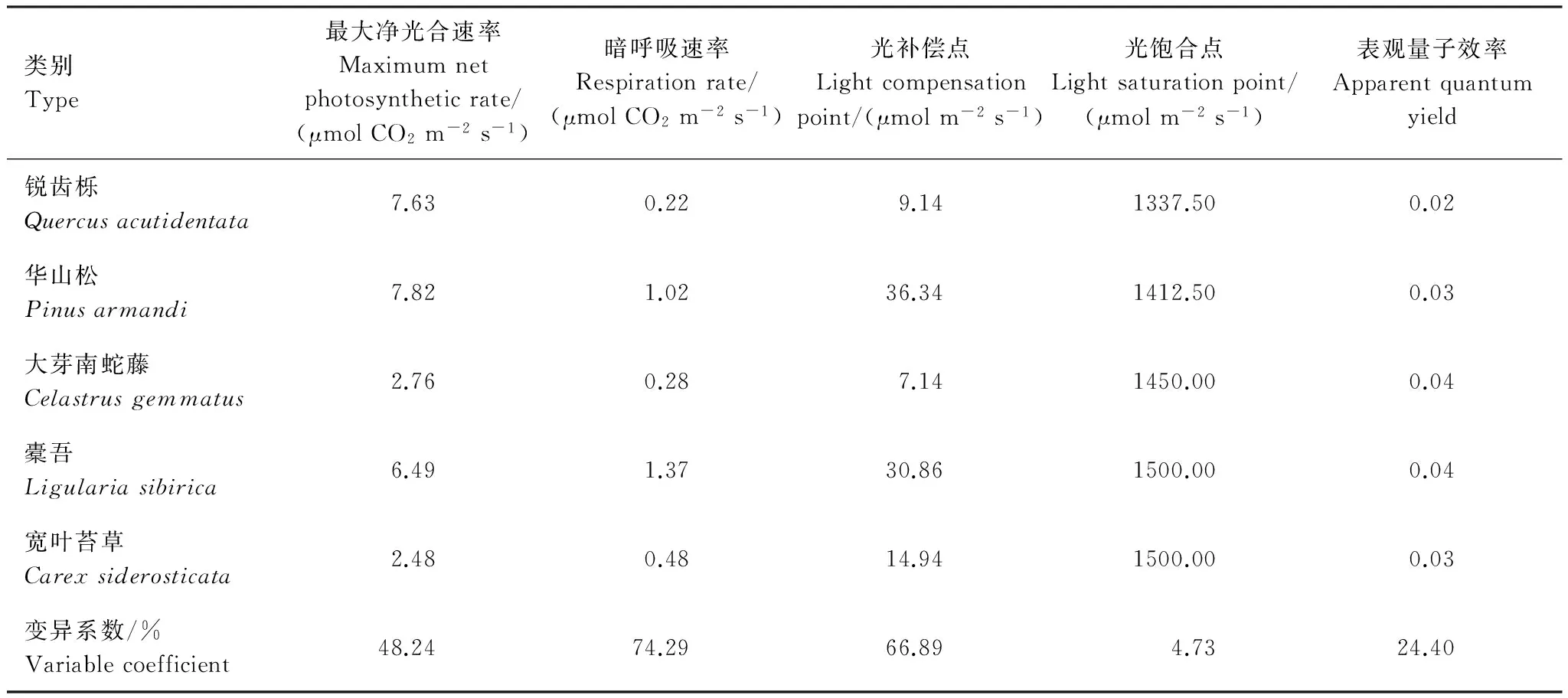
表5 第3组植物功能群光合特性
华山松在强光和弱光下都保持较高的净光合速率,表现出较强的喜光特性及光敏感性,在各个层次都发育良好,有较强的更新能力。草本层的橐吾也有较高的光合速率,加上林下光线较好,橐吾往往成片分布。青榨槭的光响应曲线处在本组的中间位置,发育为小乔木。山顶成熟林林下光线良好,灌木层的杜鹃成片出现,但其光合利用效率不高,大部分不能发育成小乔木,多以灌丛出现。宽叶苔草的光合利用效率最低,只在其它物种空缺的地方出现(图4)。
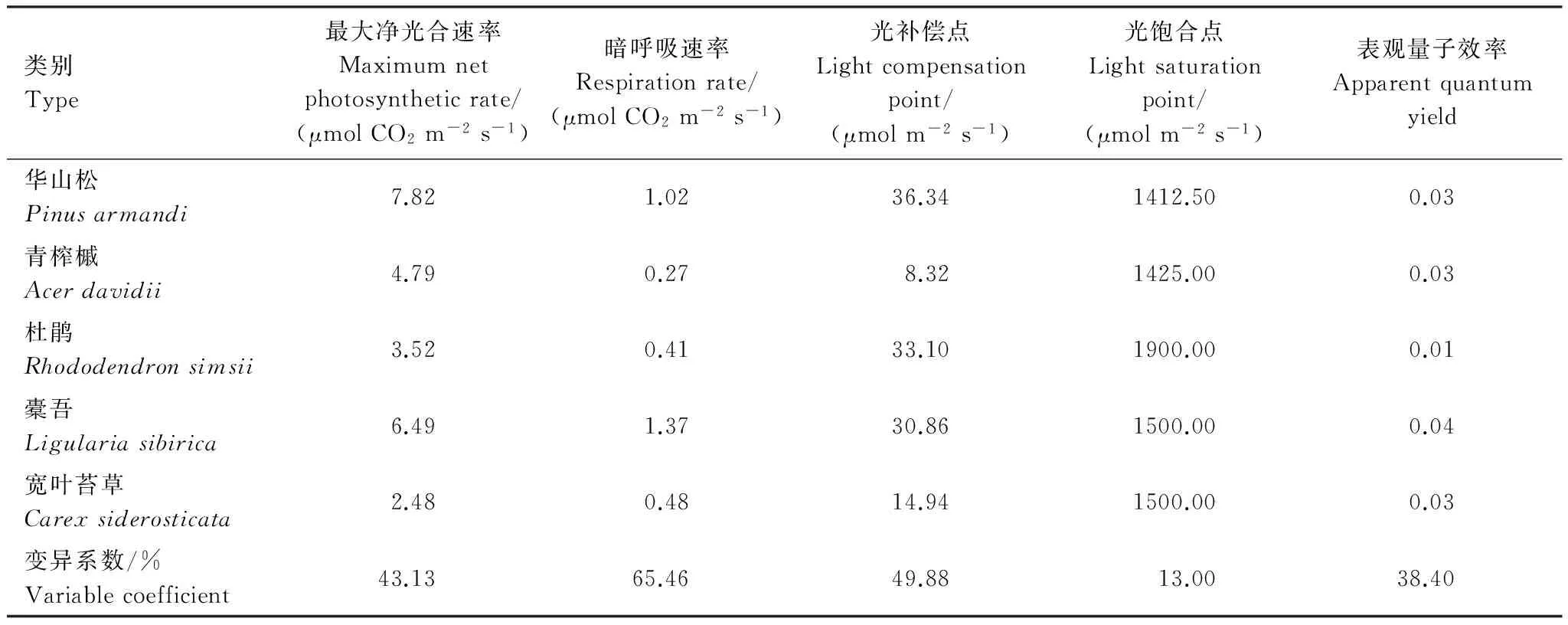
表6 第4组植物功能群光合特性

图3 第3组植物功能群光合-光响应曲线Fig.3 Photosynthesis-light response curve of the third PFG
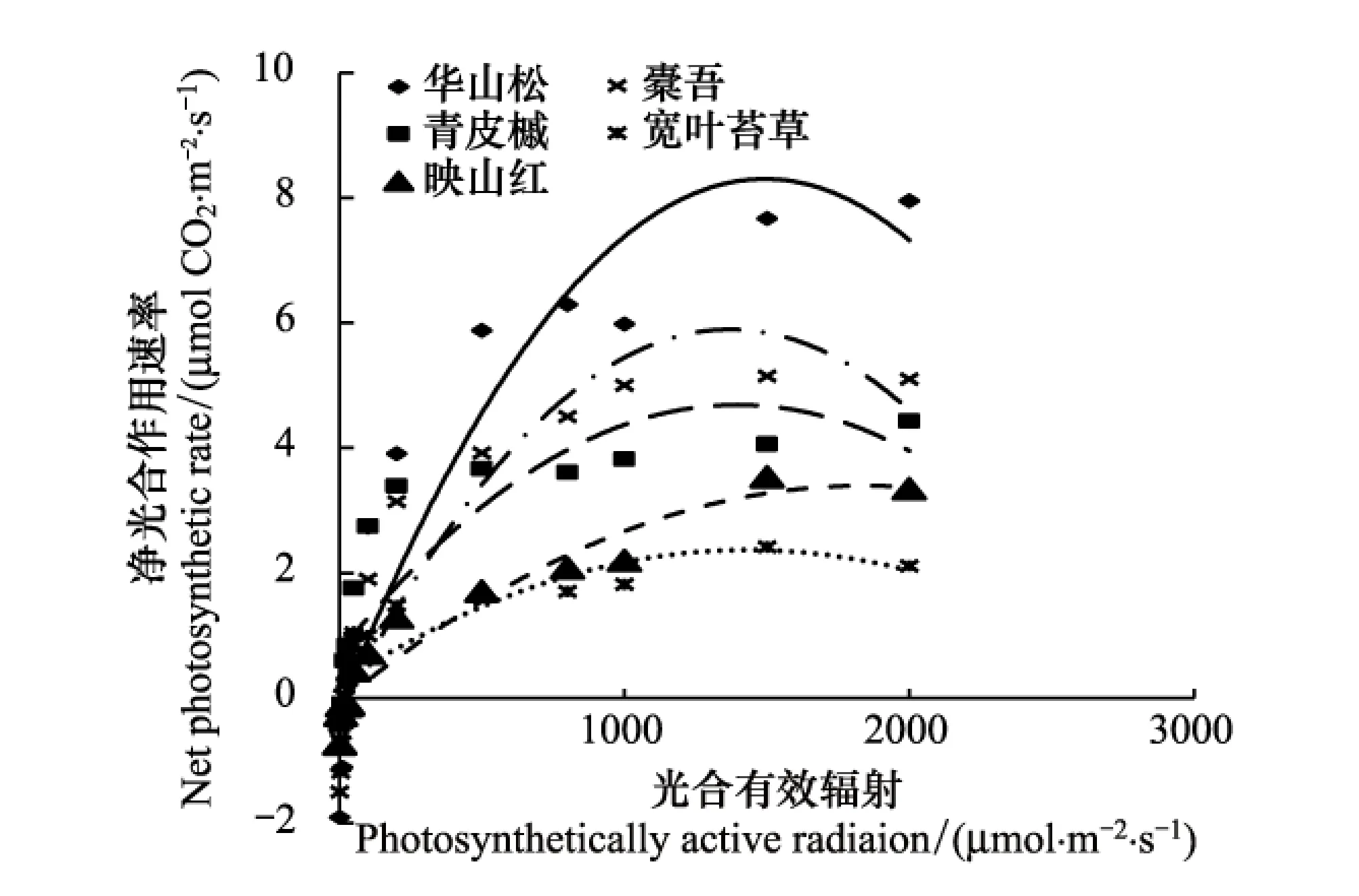
图4 第4组植物功能群光合-光响应曲线Fig.4 Photosynthesis-light response curve of the fourth PFG
总体来说,这4组植物功能群内的物种在光合特性上具有较强的互补性。它们所处分层位置与它们的光合特性联系密切。植物的光合作用受多种条件的影响,除光照强度外,温度和湿度也有较大影响。海拔、地形和坡向等因素都会影响到温度和湿度,功能群内的优势物种的光合特性具有强烈的地带性特征。也就是说,植物的光合特性依赖于其实际的分布位置,在其分布的上限或下限,光合特性会因为实际环境条件发生强烈变化,这也是整个生态系统存在植物多样性的主要原因。
4 讨论
在进行光合测定时,对不同的物种应该采取不同的最大光合有效辐射值,特别是不同层次的物种。光强值过大,会超出其平时的常见光强,不符合实际,造成数据的偏差。乔木应该选择在1 500μmol m-2s-1以内,灌草植物应该选择在100μmol m-2s-1以内。因为外界光照强度在1 000μmol m-2s-1左右,林下的光照强度在100μmol m-2s-1左右。叶龄、朝向、高度等因素也会对数值有一定的影响,所以在比较时尽可能多方位取值,以增加数据的可靠性。温度和湿度等外界因素也会影响光合速率测定数值,光强和温度经历了一个抛物线式的变动[23-24]。森林生态系统一般分布在较高的海拔,气温相对城市、裸地较低,加上森林生态系统存在分层现象,高的光强会给下层叶片提供更充足的光照,所以,从整体上来看,即使在夏季,并没出现明显的光合午休现象[25,26]。
要弄清森林生态系统的结构、功能及其生态系统服务,可以从植物功能群的组成、生境和功能特性等方面作为切入点。植物功能群内物种的联系看似松散,却是其长期协同进化的结果。植物功能群划分和其生存的自然环境有很强的相关性。在山地森林生态系统中,功能群划分与海拔直接相关,不同海拔的环境条件差异明显,形成了不同的植被分布。相应的,植物功能群组成及其功能性状就具有非常明显的地带性特征。功能群内不同物种的生理生态特性存在互补性,这种互补性是它们协同进化的基础,使植物功能群以一个整体对外界环境变动产生响应[27]。光合特性的差异与互补是其中的重要方面。光合效率是不同物种发育的基础,也是其竞争力的重要体现。在森林生态系统中,光照强度的分层使不同适应能力的乔木、灌木和草本植物聚集在一起,形成了对光的充分利用。在长期进化的过程中,不同层次的物种形成了对资源的独特利用方式,也形成了不同的植物功能群。正是这种多样性的功能群,才使得整个森林生态系统更加稳定和高效。这种差异与互补不仅存在于植物功能群内,也会在不同物种、不同尺度中广泛共存。
[1]Smith T M, Woodward F I, Shugart H H. Plant Function Types. Cambridge: Cambridge University Press, 1996.
[2]Woodward F I, Cramer W. Plant functional types and climatic change: introduction. Journal of Vegetation Science, 1996, 7(3): 306- 308.
[3]倪健. 区域尺度的中国植物功能型与生物群区. 植物学报, 2001, 43(4): 419- 425.
[4]Campbell C J, Johns C V, Nielsen D L. The value of plant functional groups in demonstrating and communicating vegetation responses to environmental flows. Freshwater Biology, 2014, 59(4): 858- 869.
[5]Laurent J M, Bar-Hen A, François L, Ghislain M, Cheddadi R. Refining vegetation simulation models: from plant functional types to bioclimatic affinity groups of plants. Journal of Vegetation Science, 2004, 15(6): 739- 746.
[6]臧润国, 张志东. 热带森林植物功能群及其动态研究进展. 生态学报, 2010, 30(12): 3289- 3296.
[7]刘增文, 王乃江, 李雅素, 吕月玲. 森林生态系统稳定性的养分原理. 西北农林科技大学学报: 自然科学版, 2006, 34(12): 129- 134.
[8]Box E O. Factors determining distributions of tree species and plant functional types. Vegetation, 1995, 121(1/2): 101- 116.
[9]Wana D, Beierkuhnlein C. Responses of plant functional types to environmental gradients in the south-west Ethiopian highlands. Journal of Tropical Ecology, 2011, 27(3): 289- 304.
[10]胡楠, 范玉龙, 丁圣彦, 卢训令. 伏牛山自然保护区森林生态系统乔木植物功能型分类. 植物生态学报, 2008, 32(5): 1104- 1115.
[11]胡楠, 范玉龙, 丁圣彦. 伏牛山森林生态系统灌木植物功能群分类. 生态学报, 2009, 29(8): 4017- 4025.
[12]范玉龙, 胡楠, 丁圣彦, 翟元杰, 柳静, 廖秉华, 卢训令. 伏牛山自然保护区森林生态系统草本植物功能群的分类. 生态学报, 2008, 28(7): 3092- 3101.
[13]McLaren J R, Turkington R. Ecosystem properties determined by plant functional group identity. Journal of Ecology, 2010, 98(2): 459- 469.
[14]Bermejo L A, de Nascimento L, Mata J, Fernández-Lugo S, Camacho A, Arévalo J R. Responses of plant functional groups in grazed and abandoned areas of a Natural Protected Area. Basic and Applied Ecology, 2012, 13(4): 312- 318.
[16]Ackerly D D. Community assembly, niche conservatism, and adaptive evolution in changing environments. International Journal of Plant Sciences, 2003, 164(S3): S165-S184.
[17]林鹏. 植物群落学. 上海: 上海科学技术出版社, 1986: 123- 128.
[18]Fry E L, Power S A, Manning P. Trait-based classification and manipulation of plant functional groups for biodiversity-ecosystem function experiments. Journal of Vegetation Science, 2014, 25(1): 248- 261.
[19]尚富德, 王正德. 伏牛山南北自然过渡地带植物多样性的特征及其成因分析. 河南大学学报: 自然科学版, 1998, 28(1): 54- 60.
[20]王伯荪, 彭少麟. 南亚热带常绿阔叶林种间联结测定技术研究—Ⅰ. 种间联结测式的探讨与修正. 植物生态学与地植物学丛刊, 1985, 9(4): 274- 285.
[21]中国科学院上海植物生理研究所, 上海市植物生理学会. 现代植物生理学实验指南. 北京: 科学出版社, 1999: 89- 95.
[22]郭志华, 张宏达, 李志安, 胡宏伟. 鹅掌楸(Liriodendronchinense)苗期光合特性的研究. 生态学报, 1999, 19(2): 164- 169.
[23]杜占池, 杨宗贵. 羊草和大针茅光合作用午间降低与生态因子关系的研究. 自然资源学报, 1990, 5(2): 177- 188.
[24]王德利, 王正文, 张喜军. 羊草两个趋异类型的光合生理生态特性比较的初步研究. 生态学报, 1999, 19(6): 837- 843.
[25]卢训令, 胡楠, 丁圣彦, 廖秉华, 房会普. 伏牛山自然保护区锐齿栎光合生理生态特性研究. 河南大学学报: 自然科学版, 2010, 40(6): 617- 621.
[26]胡楠, 范玉龙, 丁圣彦, 卢训令, 翟元杰, 柳静, 廖秉华, 王晓辉. 伏牛山自然保护区北坡主要乔木优势种光合作用特性研究. 河南大学学报: 自然科学版, 2009, 39(4): 392- 396.
[27]Eisenhauera N, Sabaisb A C W, Scheuc S. Collembola species composition and diversity effects on ecosystem functioning vary with plant functional group identity. Soil Biology and Biochemistry, 2011, 43(8): 1697- 1704.
Photosynthetic characteristics of plant functional groups in forest ecosystem at the national natural reserve of FuNiu Mountain
FAN Yulong1,2, LIU Huimin1, HU Nan2, DING Shengyan1,*
1TheCollegeofEnvironmentandPlanning,HenanUniversity,Kaifeng475001,China2NanyangInstituteofTechnology,Nanyang473004,China
A Plant Functional Group (PFG) is composed of a set of plants co-existing within a community and can serve as a basic unit of vegetation dynamic change within an environment and can act to simplify related research. The physiological characteristics of different species provide the basis for PFG classification and also affect the overall function of PFGs. The forest ecosystem has structural integrity in the National Natural Reserve of FuNiu Mountain. Through species′ interspecific associations and their dynamic changes along the elevation gradient, we classified the dominant species into four major PFGs. We then analyzed the photosynthetic characteristics of each plant of the four PFGs. Using the photosynthesis system, we measured the photosynthetic characteristics of different plants of PFGs and analyzed and compared the results which show: that the maximum Coefficient of Variation of different species′ LCP(light compensation point) and Rd(respiration rate) was 70.47% and 74.29% respectively within the same PFG; these are complementary to each other among functional traits, which is the main reason for the long-term coexistence of the PFG. Trees′ Pn(maximum net photosynthetic rate) was between 7 and 8 CO2m-2s-1which suggests that they have the ability to capture high-intensity light LSP(Light saturated point) within the same PFG was mainly between 1200—1500μmol m-2s-1, which differs very little. Herbaceous understory shrubs are more sensitive to low-intensity light than the trees. There are functional characteristic differences among the same PFG and between different PFGs, which affects the structure, function, processes, and ecosystem services of the forest ecosystem.
plant functional group (PFG); photosynthetic characteristics; FuNiu Mountain Natural Reserve; ecosystem function; ecosystem service
国家自然科学基金资助项目(41071118,40671175)
2014- 10- 16;
2016- 04- 17
Corresponding author.E-mail: syding@henu.edu.cn
10.5846/stxb201410162040
范玉龙,刘慧敏, 胡楠, 丁圣彦.伏牛山自然保护区森林生态系统植物功能群光合特性.生态学报,2016,36(15):4609- 4616.
Fan Y L, Liu H M, Hu N, Ding S Y.Photosynthetic characteristics of plant functional groups in forest ecosystem at the national natural reserve of FuNiu Mountain.Acta Ecologica Sinica,2016,36(15):4609- 4616.
