异氟烷对新生小鼠神经元细胞外信号调节激酶和cAMP反应元件结合蛋白磷酸化水平的影响
2016-10-22张斌
张斌
辽宁省锦州市中心医院麻醉科,辽宁锦州 121001
异氟烷对新生小鼠神经元细胞外信号调节激酶和cAMP反应元件结合蛋白磷酸化水平的影响
张斌
辽宁省锦州市中心医院麻醉科,辽宁锦州121001
目的观察异氟烷对小鼠神经元细胞外信号调节激酶和环磷酸腺苷(cAMP)反应元件结合蛋白磷酸化水平的影响,探究异氟烷诱导神经元损伤和学习记忆障碍的可能机制。方法原代培养小鼠神经元,分为对照组、0.96mmol/L异氟烷处理12 h及24 h组。采用MTS方法检测各组细胞活力;Western blot方法检测p-ERK1/2和p-CREB表达情况;逆转录定量PCR(RT-qPCR)方法检测ERK和CREBmRNA表达。 结果 与对照组比较,0.96 mmol/L异氟烷处理12 h及24 h后神经元活力明显下降(P<0.05);Western blot实验结果显示:与对照组比较,异氟烷处理组小鼠神经元中p-ERK1/2和p-CREB表达水平均明显降低(P<0.05);随着异氟烷处理时间的增加,p-ERK1/2和p-CREB表达逐渐降低,处理12 h与24 h比较差异有统计学意义(P<0.05)。RT-qPCR实验显示:与对照组比较,异氟烷处理组小鼠神经元中ERK和CREBmRNA表达水平比较,差异无统计学意义(P>0.05)。 结论一定浓度的异氟烷可以降低原代培养的小鼠神经元活力和ERK1/2、CREB的磷酸化水平。
异氟烷;神经元;细胞外信号调节激酶;cAMP反应元件结合蛋白
[Abstract]Objective To investigate Isoflurane-induced neuronal damage and possiblemechanisms involved in learning and memory impairments.M ethods Cerebral neurons separated from fetal mice were cultured and divided into three groups,one control group,two experimental groups were treated with 0.96 mmol/L Isoflurane for 12 h and 24 h,respectively.MTSmethod was used to determine the cell viability;Western blotmethod was used to detect p-ERK1/2 and p-CREB expressions;the reverse transcription qPCR(RT-qPCR)method was used to detectmRNA expressions of ERK and CREB.Results Compared with the control group,the viability of neuronal cells decreased significantly(P<0.05);the phosphorylation of ERK1/2 and CREB also significantly decreased(P<0.05),and with the prolonging of Isoflurane treatment duration,the expression of p-ERK1/2 and p-CREB decreased gradually,and the difference between 12 and 24 h after treatmentwas statistically significant(P<0.05);there were no statistically significant difference inmRNA expressions of the ERK and CREB(P>0.05).Conclusion A certain concentration of Isofluranemay directly inhibitneuronal cell viability and also inhibits the activities of ERK1/2 and CREB.
[Key words]Isoflurane;Neuron;ERKs;CREB
全麻药异氟烷因其价格便宜、毒副作用小、诱导快,是临床中应用广泛的吸入麻醉药之一。近年来研究表明,异氟烷急性暴露可诱发神经细胞凋亡,引起新生大鼠长期学习记忆能力下降[1-2];异氟烷还可以引发围术期成年患者术后认知功能障碍(Post-operative cognitive dysfunction,POCD)并对神经系统发育、学习及记忆产生影响[3]。细胞外信号调节激酶(Extracellular signal regulated kinases,ERKs)是丝裂原活化蛋白激酶家族成员之一,介导细胞增殖、分化及存活。在中枢神经系统中,ERK可影响神经细胞增殖分化、突触可塑性、轴突生长等,与脑内长时程增强(Long-term potentiation,LTP)的形成以及学习记忆功能密切相关[4-5]。研究证明,许多形式的突触可塑性都需要ERK参与,同时动物实验证明,抑制神经细胞中ERK活性可导致动物学习功能的降低[6]。
环磷酸腺苷反应元件结合蛋白(cAMP response element binding protein,CREB)在突触可塑性中具有重要的作用,激活CREB可促进多种与突触可塑性相关的转录因子的转录[7],从而诱导产生一些与学习记忆十分相关的蛋白分子。
本实验采用神经细胞原代培养的方法,从细胞水平观察异氟烷对神经元ERK和CREB的影响,探讨异氟烷的神经毒性以及影响学习记忆能力的潜在机制。
1 对象与方法
1.1主要材料
取24 h内新生的SPF级小鼠30只,雌雄不限,由辽宁医学院实验动物中心提供[动物许可证号:SYXK(辽)2009-0004]。DMEM培养基(美国Hyclone)、胎牛血清(FBS,浙江天杭生物科技有限公司)、神经元基本培养基(美国Introvigen)、p-CREB抗体(Cell Signal公司)、p-ERK1/2抗体 (美国SANTACruz公司)、β肌动蛋白抗体和辣根过氧化物酶(HRP)标记的二抗(北京中杉金桥生物技术有限公司)、ECL发光试剂(美国Sigma公司)、Cell Titer-BlueTM Cell Viabiliy Assay试剂盒(美国Promega公司)。
1.2小鼠大脑皮层神经元分离、培养及细胞分组
设定路径 S1={5,4,10,6,9,16,26,20,19},路径 S2={5,7,8,10,12,14,16,23,20,19},为被选中父体实施杂交,S1,S2括号里的数字为路径的节点。如去除首末节点后,相同节点有2个以上时,则将头2个相同节点来杂交,杂交点间的节点交换,生成杂交段。从S1,S2路径可以看出,除起始节点和结束节点相同外,相同节点为10、16和20,再选择起始于终点实施杂交,最终得到交换后的路径表达如下所示。
根据文献报道[8],选用出生24 h内的昆明小鼠,无菌条件下,断头处死小鼠取脑,于预冷的D-Hanks液中,分离大脑皮层后,显微镜下去除脑膜和血管;D-Hanks液洗2~3次,剪成小块。37℃下0.25%胰蛋白酶消化20 min后取出吹打,用含10%胎牛血清的DMEM终止反应。4℃下1000 r/min离心5min,2次,弃上清液。DMEM调节细胞悬液浓度为5×105/mL,接种于培养瓶中,37℃恒温的CO2培养箱内培养。半量换液,1次/2~3 d。细胞培养7 d,经神经元特异性烯醇化酶(Neuron specific enolase,NSE)免疫细胞化学法鉴定,神经细胞占全部细胞数的90%以上,取培养7 d的神经细胞进行试验。
培养细胞分为,对照组:常规培养;异氟烷暴露组:采用终浓度为0.96mmol/L异氟烷的培养基培养细胞,分别培养12 h及24 h,共三组。每组分别取10只动物,细胞培养每组设10个复孔。
1.3异氟烷溶液的配制
根据文献[8]的方法,将130μL液体异氟烷加入103mL平衡盐溶液(Balanced salt solution,BSS)中,密封后摇床上摇匀24 h,充分溶解异氟烷,配制10mmol/L储备液。使用前迅速将30 mL溶液倒入50 mL聚丙烯离心管中,震荡5~10 s,加入培养基(终浓度为0.96mmol/L)后用于细胞处理。
1.4MTS方法测定细胞活力
将细胞悬液稀释至1×105/mL,以每孔1×104个细胞接种于96孔板,每孔100μL。将培养板放入CO2培养箱内37℃培养过夜后,更换含有异氟烷的培养液继续培养24 h。根据说明书,每孔加入20μLMTS试剂,37℃下孵育4 h,592 nm处测定各组细胞吸光度值A。根据公式计算生存率,细胞生存率=(A各组-A空白孔)/(A对照组-A空白孔)×100%。
1.5免疫印迹检测神经元ERK和CREB蛋白表达
1.6 RT-qPCR检测神经元ERK和CREB mRNA表达
Trizol法提取总RNA。SYBR GreenⅡ进行检测,热循环条件:95℃3 min,94℃ 15 s,60℃ 60 s,40循环。PCR扩增反应体积10μL:RNase Free dH2O23μL,SYBRRPrimeScriptTM(2×)5μL,ERK、CREB、18Sr-RNA上下游引物 (10μmol/L)1μL,DNA模板1μL,每个样品3个复孔。采用比较Ct值(2-ΔΔCt法)相对定量法,改变的倍数(fold change)=2-ΔΔCt;ΔCt=Ct靶基因-Ct内参,ΔΔCt=ΔCt处理组-ΔCt未处理组。以18SrRNA为内对照。以2-ΔΔCt表示实验组目的基因的表达相对于对照组的变化倍数。PCR引物序列见表1。

表1 ERK、CREB、18SrRNA引物序列
1.7统计学方法
采用SPSS 18.0统计学软件进行数据分析,计量资料数据用均数±标准差(x±s)表示,组间计量资料比较采用单因素方差分析(ANOVA)检验,组间两两比较采用LSD-t检验,计数资料用率表示,组间比较采用χ2检验,以P<0.05为差异有统计学意义。
2 结果
2.1异氟烷对小鼠神经元生存率的影响
与对照组比较,12 h和24 h异氟烷处理组的生存率降低,生存率从对照组的100%分别降至(86.58±8.96)%和(78.63±8.23)%,差异均有统计学意义(P<0.05)。见图1。
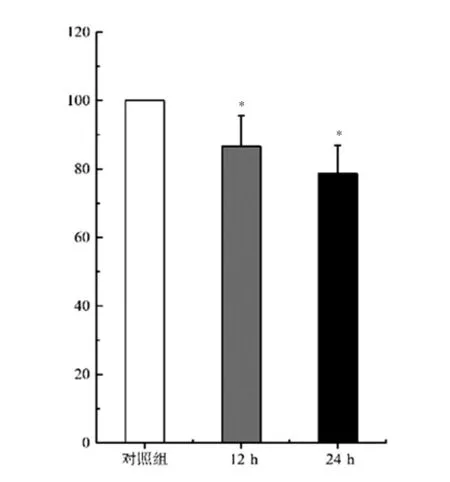
图1 异氟烷处理不同时间后细胞生存率变化(n=10)
2.2异氟烷对小鼠神经元ERK1/2和CREB活性的影响
Western blot方法检测小鼠神经元 ERK1/2和CREB活性,结果显示,与对照组比较,异氟烷处理组小鼠神经元中p-ERK1/2和p-CREB表达水平均明显降低 (P<0.05);随着异氟烷处理时间的增加,p-ERK1/2和p-CREB表达逐渐降低,处理12 h与24 h比较差异有统计学意义(P<0.05)。见图2。
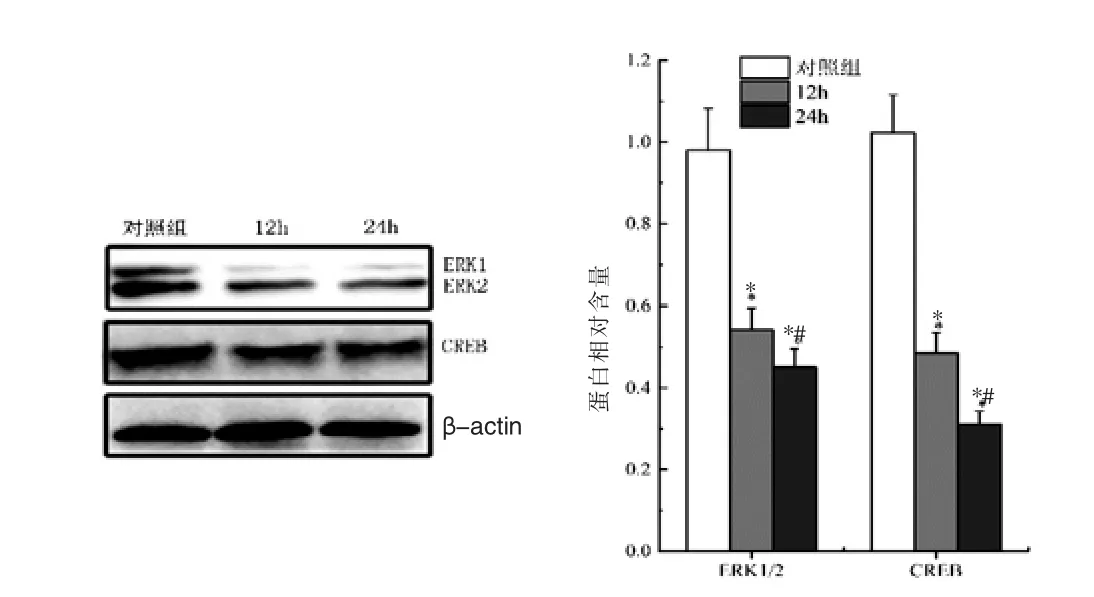
图2 异氟烷对小鼠神经元ERK1/2和CREB活性的影响(n=10)
2.3异氟烷对小鼠神经元ERK1/2和CREB mRNA表达的影响
RT-qPCR方法检测了小鼠神经元ERK和CREB的mRNA表达。与对照组比较,异氟烷处理组小鼠神经元中ERK和CREB表达差异无统计学意义(P>0.05)。见图3。
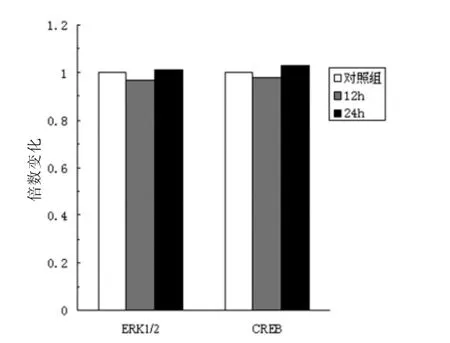
图3 异氟烷对小鼠神经元ERK1/2和CREBm RNA表达的影响(n=10)
3 讨论
全身麻醉药目前已广泛应用于临床麻醉中,而其对学习记忆功能的影响也受到重视,有研究证实,吸入性麻醉药异氟烷可以诱导产生细胞毒性,造成大鼠中枢神经系统神经元凋亡[9-10],引起未成年小鼠大量脑神经细胞凋亡,产生术后认知功能或持续性学习功能障碍[11]。本实验从细胞水平探讨了异氟烷的神经毒性以及影响学习记忆能力的潜在机制。
本研究结果显示:与对照组比较,0.96 mmol/L异氟烷处理后小鼠神经元活力明显下降(P<0.05);磷酸化ERK1/2和CREB显著降低(P<0.05)。结果提示,一定浓度的异氟烷可以降低原代培养的小鼠神经元活力和ERK1/2、CREB的磷酸化水平,可能与异氟烷诱导学习记忆损伤及术后POCD相关。
ERK是一类丝/苏氨酸蛋白激酶,在神经系统中,ERK分布广泛,参与突触可塑性长时程增强电位诱导过程[12],也参与神经元的凋亡,在记忆过程中发挥重要作用。研究证明,将ERK通路阻断后,可抑制LTP的形成[13],而影响学习记忆功能。Mawh inney等[14]发现,异氟烷可降低海马和皮质的磷酸化ERK1/2,导致空间认知功能障碍。Liu等[15]发现,低浓度及短时间应用异氟烷可以提高pERK1/2水平,可能改善认知功能,而高浓度应用异氟烷超过4 h则会降低pERK1/2水平,而损害认知功能。本研究也发现,0.96 mmol/L的异氟烷溶液作用能对小鼠神经元产生明显的毒性作用,且随处理时间延长,p-ERK1/2的表达逐渐降低(P<0.05)。
CREB是一种分子量为43 kD的核转录因子蛋白,参与了神经干细胞增殖、细胞周期调控、学习记忆等正常生理活动[16]。CREB上Ser-133的磷酸化位点是许多蛋白激酶的作用靶点,一旦被磷酸化,CREB介导的转录启动并激发长时程增强电位相关蛋白的合成,对于神经突触可塑性长时间改变有重要作用[17]。有结果表明在脊椎动物CREB基因被破坏后,其长时记忆形成受到影响。施小娇等[18]发现:高浓度七氟烷可以引起大鼠麻醉后早期认知功能障碍,并呈剂量依赖性,CREB蛋白磷酸化的抑制以及记忆相关蛋白合成的减少可能是机制之一。本研究中异氟烷处理组小鼠神经元中 p-CREB表达水平均明显降低 (P<0.05);并随处理时间延长而逐渐降低(P<0.05)。
依文献报道[19],0.28mmol/L异氟烷相当于临床上1个最小肺泡浓度(Minimal alveolar concentration,MAC)的血药浓度,一般临床常用1.3 MAC。本研究将神经元暴露于0.96mmol/L的异氟烷浓度中,明显高于临床常用量,随着处理时间的延长,神经元活力逐渐下降,与学习与记忆有关的蛋白表达受到抑制。结果提示临床中降低用药浓度,减少使用时间将有助于减少认知障碍及学习记忆能力下降的发生。
综上所述,本研究结果显示,异氟烷的神经毒性可能是由于异氟烷直接造成了神经细胞活力的下降,同时也显著抑制了神经元内学习记忆相关的重要蛋白ERK和CREB的活性。这些现象可能与异氟烷引发围术期成年患者术后POCD相关。可能机制包括线粒体功能障碍、氧化应激、抑制海马神经元脑源性神经营养因子前体的水解,促进神经元凋亡等[20]。还需要更多的工作来深入研究异氟烷与ERK/CREB通路间复杂的相互作用,以明确异氟烷在神经损伤中的确切作用。
[1]Jevtovic-Todorovic V,Hartman RE,IzumiY,etal.Early exposure to common anesthetic agents causeswidespread neurodegeneration in the developing rat brain and persistent learning deficits[J].JNeurosci,2003,23(3):876-882.
[2]余梓浦,王佩芳,李纯,等.七氟烷和异氟烷急性暴露对SD仔鼠海马的神经毒性作用[J].中国药理学与毒理学杂志,2011,25(1):72-76.
[3]RammesG,Starker LK,HasenederR,etal.Isofluraneanaesthesia reversibly improves cognitive function and longterm potentiation(LTP)via an up-regulation in NMDA receptor2B subunitexpression[J].Neuropharmacology,2009,56(3):626-636.
[4]Di Cristo G,Berardi N,Cancedda L,et al.Requirement of ERK activation for visual cortical plasticity[J].Science,2001,292(5525):2337-2340.
[5]Lee SE,Simons SB,Heldt SA,et al.RGS14 is a natural suppressor of both synaptic plasticity in CA2 neurons and hippocampal-based learning and memory[J].Proc Natl Acad Sci USA,2010,107(39):16994-16998.
[6]Villarreal JS,Barea-Rodriguez EJ.ERK phosphorylation is required for retention of trace fearmemory[J].Neurobiol Learn Mem,2006,85(1):44-57.
[7]Porte Y,BuhotMC,Mons NE.Spatialmemory in the Morris water maze and activation of cyclic AMP response element-binding(CREB)protein within themouse hippocampus[J].Learn Mem,2008,15(12):885-894.
[8]王丛丛,朱正华,吕文英,等.抑制CaMKII对异氟烷神经毒性的影响[J].中华神经外科疾病研究杂志,2011,10(5):420-423.
[9]Istaphanous GK,Howard J,Nan X,et al.Comparison of the neuroapoptotic properties of equipotent anesthetic concentrations of desflurane,isoflurane,or sevoflurane in neonatalmice[J].Anesthesiology,2011,114(3):578-587.
[10]Zhou ZW,Ma DQ.Anaesthetics-induced neuro?toxicity in developing brain:an update on preclinicalevidence[J]. Brain Sci,2014,4(1):136-149.
[11]Lunardi N,Ori C,Erisir A,et al.General anesthesia causes long-lasting disturbances in the ultrastructural properties of developing synapses in young rats[J].Neurotox Res,2010,17(2):179-188.
[12]Evans PR,Dudek SM,Hepler JR.Regulator of G Protein Signaling 14:A Molecular Brake on Synaptic Plasticity Linked to Learning and Memory[J].Prog Mol Biol Transl Sci,2015,133:69-206.
[13]Yang MT,Chien WL,Lu DH,et al.Acetazolamide impairs fear memory consolidation in rodents[J].Neuropharmacology,2013,67(4):412-418.
[14]Mawhinney LJ,de Rivero Vaccari JP,Alonso OF,et al. Isoflurane/nitrous oxide anesthesia induces increases in NMDA receptor subunit NR2B protein expression in the aged rat brain[J].Brain Res,2012,1431(1):23-34.
[15]Liu J,Wang P,Zhang X,et al.Effects of different concentration and duration time of isoflurane on acute and long-term neurocognitive function of young adult C57BL/ 6mouse[J].Int JClin Pathol,2014,15,7(9):5828-5836.
[16]宋海龙,郑焱,王晓民.CREB信号通路在神经系统中的调控及功能[J].中国生物化学与分子生物学报,2013,29(9):797-804.
[17]Paramanik V,Thakur MK.Role of CREB signaling in aging brain[J].Arch Ital Biol,2013,151(1):33-42.
[18]施小娇,金梦,张晓暄,等.不同浓度七氟烷对大鼠认知功能及海马内磷酸化cAMP反应元件结合蛋白表达的影响[J].中国老年杂志,2014,34(20):5807-5809.
[19]Boban M,Stowe DF,Buljubasic N,etal.Directcomparative effects of isoflurane and in isolated guinea pig hearts[J]. Anesthesiology,1992,76(5):775-780.
[20]饶婉宜,鲁惠顺.异氟烷诱导术后认知功能障碍的机制及预防的研究进展[J].中国药理学与毒理学杂志,30(2):178-181.
Effect of Isoflurane exposure on expressions of ERKs and CREB in neuron of newborn mice
ZHANG Bin
Department of Anesthesiology,Jinzhou Central Hospital,Liaoning Province,Jinzhou121001,China
R614.2
A
1673-7210(2016)09(b)-0022-04
2016-06-10本文编辑:任念)
张斌(1977.10-),男,副主任医师;研究方向:神经阻滞、体外循环。
