内蒙古羊草草原凋落物分解过程中营养元素的动态
2016-10-20母悦耿元波
母悦,耿元波*
内蒙古羊草草原凋落物分解过程中营养元素的动态
母悦1,2,耿元波1*
1. 中国科学院地理科学与资源研究所,北京 100101;2. 中国科学院大学,北京 100049
凋落物分解是生态系统养分循环的关键环节,作为植物体向土壤归还营养物质的主要途径,对维持生态系统的平衡具有重要的意义。以内蒙古锡林河流域的羊草草原优势种羊草(Leymus chinensis)和冰草(Agropyron cristatum)、羊草和冰草双物种、多物种混合凋落物为研究对象,采用凋落物袋法研究了4类凋落物的分解特征及营养元素动态变化过程。结果表明:多物种、双物种、羊草和冰草凋落物的分解常数分别为1.04、0.98、1.01和0.95;4类凋落物各元素表现为不同的动态变化特征,C、P、K、Mg、Ca、Cu为净释放,平均释放率在16.37%~87.43%,Fe为累积模式,累积率为140%~339%;羊草、冰草、多物种凋落物的Mn为净累积,累积率为35.07%~57.28%,而双物种凋落物的Mn为净释放,释放率仅为0.9%;羊草凋落物的Zn为净累积,累积率为70.76%,其它凋落物表现为净释放,释放率为4.09%~22.32%。混合凋落物分解存在非加性效应,在0~305 d内,羊草和冰草凋落物之间存在着抑制作用。羊草和冰草凋落物混合对C、Cu残留率起抑制作用,对Mn、Zn、Ca、Fe残留率起促进作用,而对N残留率先起促进作用后起抑制作用,对P、Mg、K的残留率无显著影响。
羊草草原;凋落物;营养元素;混合效应
引用格式:母悦, 耿元波. 内蒙古羊草草原凋落物分解过程中营养元素的动态[J]. 生态环境学报, 2016, 25(7): 1154-1163.
MU Yue, GENG Yuanbo. The Dynam ics of Nutrients during Litter Decom position in Leymus chinensis Grassland in Inner Mongolia,China [J]. Ecology and Environmental Sciences, 2016, 25(7): 1154-1163.
凋落物分解是植物体将营养物质归还土壤的主要途径(张建利等,2008),也是生态系统土壤有机碳输入的主要来源,在陆地生态系统营养元素循环中占据重要地位(Aerts,2006;Rovira et al.,2010),就草原生态系统而言,凋落物的分解影响着草原植物萌发、群落结构和植被演替,在土壤理化性状改善、生态系统能量流动和营养物质循环等方面都起着重要作用(Facell et al.,1991;郭继勋等,2000)。因此,凋落物分解的研究对我们了解陆地生态系统的功能十分重要。凋落物分解是一个复杂的过程,受生物因素和非生物因素的影响,凋落物的化学性质(Voříšková et al.,2011),气候和土壤性质(Aerts,1997;A raujo et al.,2012),分解者群落(Chapman et al.,2013)等都影响凋落物的分解速率。C/N比值、P以及木质素的含量是判断凋落物分解速率的重要指标(Palm et al.,1991;Kemp et al.,2003),这些因素因空间尺度的不同对凋落物分解的影响程度不同。凋落物物种的组成变化通过不同物种之间的相互作用改变凋落物分解并影响生态系统的养分循环(Vivanco et al., 2006)。Gartner et al.(2004)研究发现,76%的混合凋落物在分解过程中表现出了非加性效应,混合凋落物的养分含量有可能上升也有可能下降,比基于组分凋落物计算的期望值偏离25%~100%。
自然状态下,牧草营养物质的外部来源主要是凋落物分解,研究优势种、混合物种凋落物中营养元素的动态变化十分必要,尤其是Ca、Mg、Fe、Mn、Cu、Zn等对植物和动物都十分重要的营养元素。然而,近些年来有关草原凋落物植物营养元素的研究中(赵吉等,1992;张彩虹等,2011;邵玉琴等,2004),有关C、N、P、K的研究比较多,而Ca、Mg、Fe、Mn、Cu、Zn等的研究较少,在草原日益退化的背景下,亟待加强此方面的研究工作。本文以羊草草原4类凋落物为研究对象,分析C、N、P、K、Ca、Mg、Fe、Mn、Cu和Zn等植物营养元素以及纤维素、木质素在凋落物分解过程中的变化规律,研究凋落物分解特征以及多物种凋落物的非加性效应。
1 研究区概况
锡林郭勒草原位于内蒙古高原中部,北纬43°16′~44°339′,东经1115°32′~117°12′,属于国际地圈-生物圈计划(IGBP)全球变化研究典型中国东北陆地样带(NECT),是内蒙古高原半干旱草原区的主体部分。锡林河流域属于中温带亚干旱大区,其气候类型属于温带半干旱草原气候,年平均气温为(0.3±0.11)℃,全年光照为2603.8 h,多年平均降水量为(3347.0±79.6)mm,降水量年际变化较大,主要集中在6—9月份,约占全年降水量的80%(陈佐忠,19888)。
本研究选择的实验地点(图1)位于内蒙古锡林郭勒草原锡林河流域中国科学院内蒙古草原生态系统定位研究站附近的羊草99样地(43°32′N,116°40′E),该样地自1999年起进行围栏封育。典型草原是锡林河流域面积最大、分布最广的草原,羊草(Leymmus chinensiss)草原是最具代表性的典型草原之一。实验样地内的建群种为根茎禾草羊草,优势种包括冰草(Agroopyron cristatum)和大针茅(Stipa granddis)等密丛禾草。牧草在4月中下旬返青,生长期在150~160d(李博等,1988)。主要的土壤类型为钙积干润均腐土,土壤质地为砂壤质(汪久文等,1988)。
2 研究方法
2.1实验设置
2014年8月下旬在羊草99样地选取植物群落均一且植物种较为接近的地段,平行设置5组1 m×1 m的样方,每组5个,共25个1 m×1 m样方,分物种齐地面剪取植物地上部分活体,将各样方内采集的植株按种均匀混合,根据样地所调查的物种数(图2),进行单物种、双物种、多物种凋落物的选择、配比和制备。

图1 研究样点的位置Fig. 1 Location of stu dy site
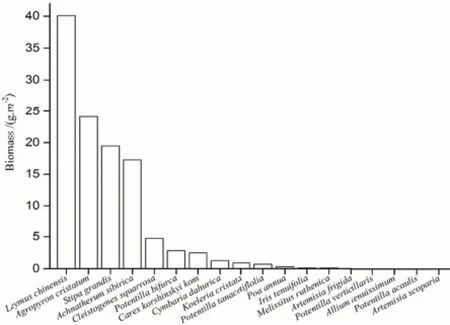
图2 羊草草原样方中各植物活体生物量Fig. 2 Abovveground biomassin Leymus chinenssis steppe
选择羊草(LC处理)和冰草(ACC处理)作为单一凋落物供试种;根据所调查两种物种生物量所占比例进行配比作为双物种凋落物(LA处理),配比为5∶3;多物种凋落物(MT处理)选择相应样地样方中的全部物种(图2包含的全部物种),按照样地内生物量比例配置,均匀混合。将供试种剪为8 cm左右的小段,装入20 cm×30 cm、孔径为2 mm×2 mm的尼龙网袋,每袋约20 g,每种凋落物30袋,4个处理共120袋,利用尼龙网袋法进行凋落物的分解研究(刘增文等,2006)。
2014年8月底,选取地势平坦的地段将凋落物袋放回羊草样地,紧贴地面,并用竹筷固定,每个尼龙网袋间隔50 cm。分别在2015年5月(返青期)、7(结实期)、9月(果后营养期)收集凋落物袋(廖仰南等,1994),4种凋落物各10袋(重复样),每次取回40袋。采集的凋落物先去除表面附着的泥沙和其他杂质,再用清水洗净,转移到信封中,置于65 ℃鼓风干燥烘箱,烘干至恒重,称量余重。凋落物称重后粉碎,进行元素测定(任来阳等,2013)。
2.2样品分析
凋落物C、N含量用元素分析仪(Flash 2000)测定,待测的凋落物样品经体积比为5∶1的混酸(HNO3-HClO4)消解后用ICP-OES(Optima 5300DV)测定K、P、Ca、Mg、Cu、Fe、Zn、Mn含量,以植物成分分析标准物质GSV-2作质量控制,测定值在标准范围内,平行样相对标准偏差小于5%,纤维素和木质素的测定采用重量法测其相对含量(王玉万等,1987;薛惠琴等,2001),样品测试结果满足研究需要。
2.3数据处理
2.3.1残余重
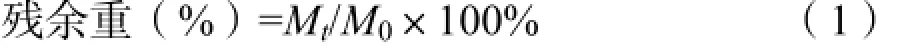
式中,Mt表示分解t天后凋落物残留的重量(g),M0表示分解凋落物的初始重量(g)。
2.3.2分解速率常数
分解速率常数k按照下列指数方程计算(Olson,1963):

式中,t为分解时间(a),k为分解常数,a为拟合参数。
2.3.3半衰期和周转期
由k值估算凋落物分解的半衰期和周转期:

2.3.4元素残留率

式中,C0为凋落物的初始养分浓度,Ct为t时间后凋落物的浓度。
2.3.5混合凋落物的期望残留重和元素期望残留率(Hoorens et al.,2003)
式中,MA为凋落物单独分解时的实测值,X为凋落物A在混合凋落物的比重(%);MB为凋落物单独分解时的实测值,Y为凋落物B在混合凋落物的比重(%)。混合凋落物重量及元素残留率期望值与实测值的比较采用t检验。根据t检验的结果判断组分凋落物是否存在相互作用:若期望值与实测值无明显差异(P>0.05),则组分之间无相互作用;若期望值显著(P<0.05)或极显著(P<0.01)低于实测值,则组分凋落物之间存在抑制作用;若期望值显著(P<0.05)或极显著(P<0.01)高于实测值,则组分凋落物之间存在促进作用。
采用单因素方差分析法检验不同时期凋落物之间的化学计量比、不同种类之间的凋落物质量残留率和元素残留率是否存在显著性差异,并利用Duncan法进行多重比较,采用独立样本t检验对期望残余重和实测残余重、期望元素残余率和实测残余率之间的差异进行显著性检验,采用回归分析对分解速率常数进行指数拟合。所有的统计分析都在SPSS Statistics 19中进行,利用OriginPro 9.1作图。
3 结果分析
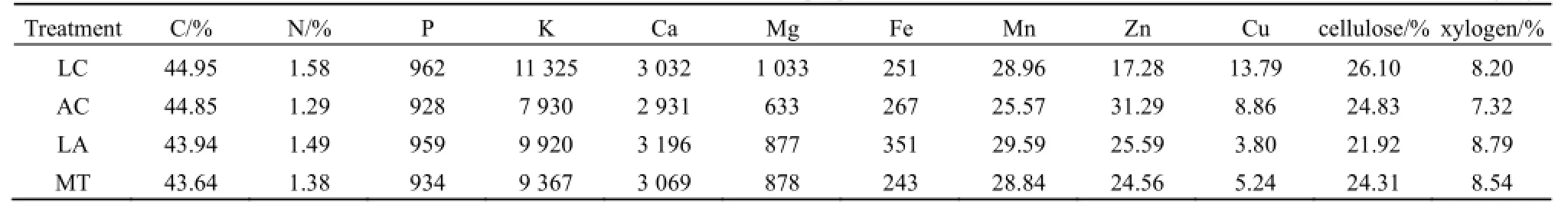
表1 凋落物的初始化学组成Table 1 Initial chemical properities of litters mg·kg-1
3.1凋落物分解营养元素的动态变化
3.1.1凋落物的初始化学组成
由表1可以看出,4类凋落物的初始化学组成各不相同,LC的C、N、P、K、Mg、Cu和纤维素的含量较高,AC的Zn含量较高,LA的Ca、Fe、Mn以及木质素含量较高。
3.1.2凋落物的质量残留率变化
4类凋落物质量残留率在不同分解阶段,其质量损失各不相同(图3)。前244 d,AC的质量损失最快,其次是LC,分解最慢的是MT,其中LC和AC的质量残留率有显著差异;在244~305 d,质量损失最快的是LC,其次是AC,MT分解速率最慢;分解305 d后,AC、LA、MT之间质量残留率有显著差异;305~367 d,MT质量损失速率最快,其次是LA。总体上,各类凋落物分解速率或快或慢,但在367 d后质量残留率无显著差异,维持在59%左右。4类凋落物在分解的初期(244 d),质量损失为16.24%~20.11%,负指数衰减模型能够较好地预测各凋落物的分解过程,且R2值的范围为0.899~0.999(表2)。凋落物在367 d的分解过程中以多物种凋落物分解常数最大,分解最快;其次是羊草,分解常数最小的是冰草。4类凋落物分解95%所需时间是2.88~3.15 a。
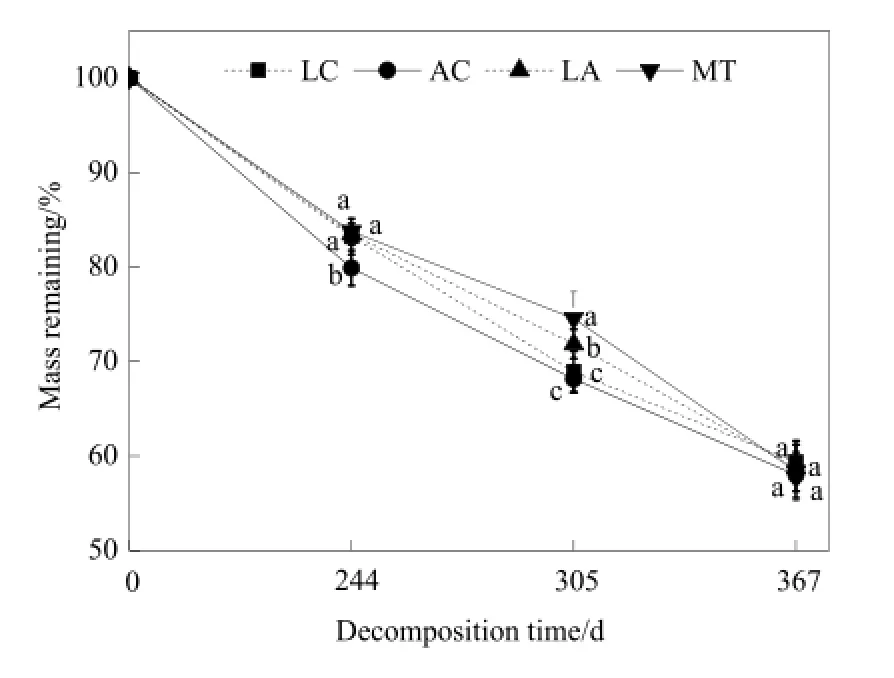
图3 凋落物的质量残留率变化Fig. 3 Litter decomposition of different plant species

表2 凋落物分解的速率(k)、决定系数(R2)、半衰期(t0.5)及分解95%所需时间(t0.95)Table 2 Litter decomposition rate (k), the coefficient of determ ination(R2), the half-life (t0.5) and the time of decompose 95% (t0.95)
3.1.3凋落物的元素残留率变化
由图4可知,在244、305 d时,LA和MT凋落物的C元素残留率无显著差异,367 d时,除LC,其它3类凋落物的C的净释放率在35.12%~41.02%,无显著差异(P>0.05)。LC的N均表现为净释放,AC的N先释放再累积,LA和MT凋落物N表现为先释放后累积再释放,经过367 d的分解,4类凋落物的N净释放率为26.73%~40.84%。4类凋落物的P、K、Mg在0~305 d,均表现为释放模式;在305~367 d,MT持续释放,其它类型开始累积。367 d时,4类凋落物的P净释放率为47.84%~51.92%,无显著差异(P>0.05),K净释放率为85.62%~ 89.66%,Mg净释放率为17.74%~31.41%。MT的Ca在0~305 d,均为缓慢释放模式,后有小幅度累积,其它3类凋落物则先累积后释放再累积,367 d时,4类凋落物总的Ca净释放率为11.94%~21.49%。4类凋落物的Fe在0~244 d先累积;244~305 d,LC的Fe开始释放,其它凋落物则持续累积;在305~367 d,MT的Fe开始释放,其它凋落物为累积模式;367 d时,4类凋落物累积率为140%~339%,LC、AC在367 d达到最大净累积量,另外两类凋落物的净累积量在305 d达到最大。4类凋落物的Mn在0~244 d为累积模式;244~367 d,MT的Mn先累积再释放,其它凋落物的Mn累积,累积率为35.07%~57.28%,LA则与之相反,为先释放再累积;367 d后,LC、AC、MT状态几乎与初始元素相同,释放率仅为0.9%。AC的Zn相反,为先释放再累积,367 d后,LC、AC、MT状态几乎与初始元素相同,释放率仅为0.9%。AC的Zn在整个分解阶段先持续释放再开始累积,其它凋落物Zn则是先累积后释放再累积,最后LC的Zn为净累积,累积率为70.76%,其它凋落物为净释放,释放率为4.09%~22.32%。LC、AC的Cu在整个分解阶段先持续释放,在305 d开始累积;LA先累积,在244 d开始持续释放;MT在初始阶段先释放,244 d后开始累积,305 d后为释放模式;各凋落物最终表现为释放模式,释放率为22.36%~72.05%。
3.2凋落物的化学计量特征变化
C/N是表征有机物质组成的重要指标,能反应有机物的无机化程度,C/N比值越小,无机化的程度越高,微生物活性增大,有利于凋落物的分解。LC的初始C/N、木质素/N最小,分别为28.45、5.19;MT的初始C/N、木质素/N最大,分别为31.58、6.18;LA的初始C/P、纤维素/N最小,分别为458.32、14.72;AC的初始C/P、纤维素/N最大,分别为483.36、19.30。由图5可知,MT、LA的C/N分别在305、367 d值最低,且在305~367 d,这两类凋落物的木质素/N迅速降低,木质素开始分解,它们的质量损失率在这一阶段变化最快。
3.3混合凋落物的分解
3.3.1凋落物分解的混合效应
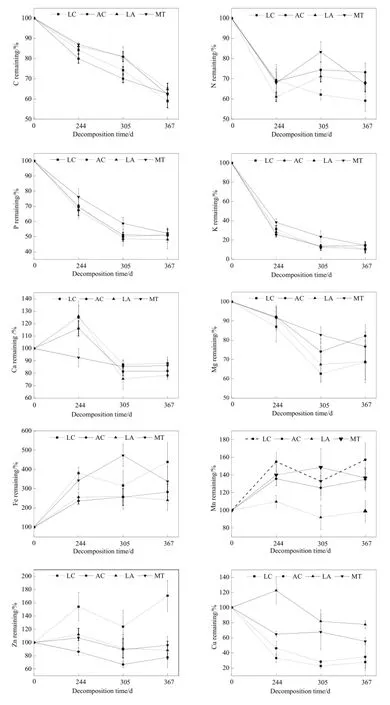
图4 凋落物分解元素残留率的变化Fig. 4 Element remaining of different plant species
从羊草和冰草混合分解的各阶段残留率来看(图6),在分解的244 d,实际测量的残余重量显著高于期望残余重量(F=1.495,P=0.017),在分解的305 d,实际残余重量极显著高于期望残余重量(F=0.18,P=0),说明在分解的305 d内,组分凋落物之间存在着抑制作用;而在分解的367 d,期望残余重量和实际测量的残余重量无显著差异,则说明在分解的305 d后,这种抑制作用开始减小。
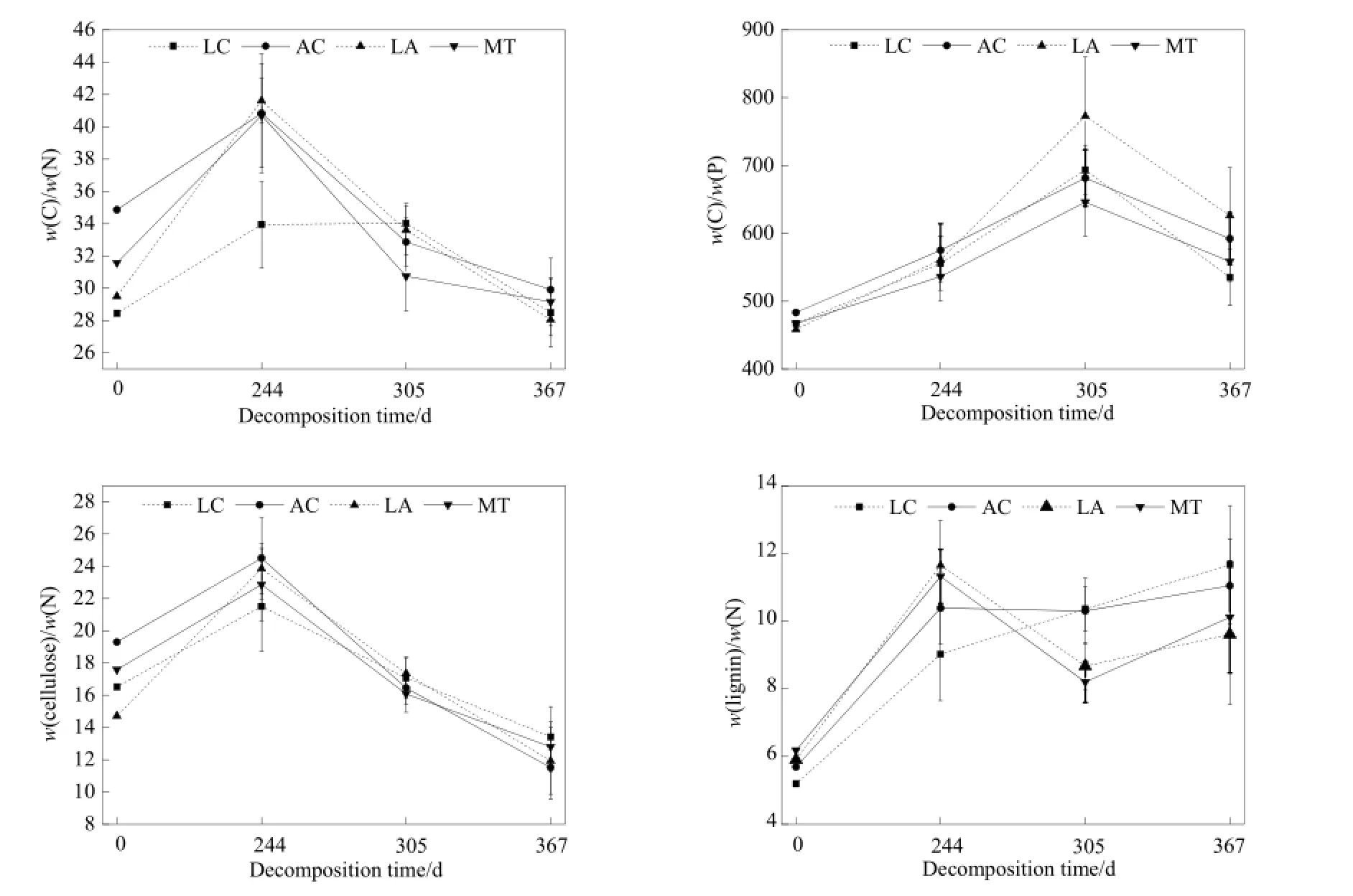
图5 凋落物分解C/N、C/P、纤维素/N、木质素/N的变化Fig. 5 C/N, C/P, cellulose/N, lignin/N of different plant species
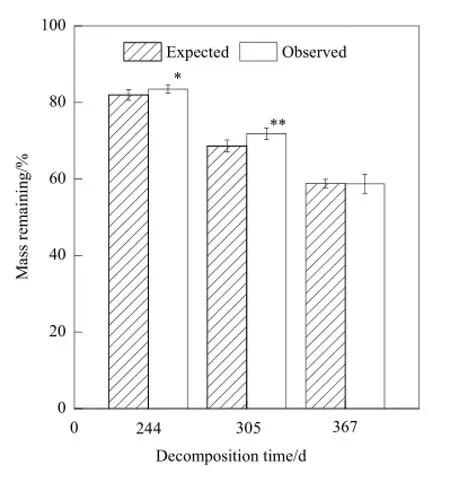
图6 期望残余重量和实测残余重量的比较Fig. 6 The difference between the Expected and Observed mass remaining
3.3.2凋落物混合后对营养元素动态的影响
将混合后凋落物元素残留率的实际测量值与期望元素残留率值进行比较(如图7)。在整个分解阶段,C、Cu的期望残留率极显著小于实测C、Cu残留率,Mn、Zn的期望残留率显著大于实测Mn、Zn残留率,羊草和冰草凋落物的混合对C、Cu残留率起抑制作用,对Mn、Zn残留率起促进作用。整个分解研究阶段,N的期望残留率与实测残留率有显著差异,在分解的0~244 d,N的期望残留率大于实测残留率,而在244~305 d,N的期望残留率开始下降,而N实测残留率呈上升趋势,N的期望残留率先大于实测残留率,后又小于实测残留率;分解的305~367 d,N期望残留率都小于实测残留率,羊草和冰草凋落物的混合对N残留率先起促进作用,随着分解的进行,这种促进作用开始消失,最后抑制凋落物的N残留率。在整个分解阶段,P、Mg期望残留率先小于实测残留率,随着分解的进行,又大于实测残留率,但两者之间无显著差异。在0~305 d,K的期望残留率和和实测残留率无显著差异,在305~367 d,K期望残留率逐渐升高,显著大于实测值,因此,羊草和冰草凋落物的混合对K残留率有一定的促进作用。0~244 d,Ca的期望残留率小于实测残留率,但无显著差异,305 d后,Ca的期望残留率极显著大于实测残留率。整个分解阶段,Fe的期望残留率大于实测残留率,但在分解244和367 d时达到极显著水平,羊草和冰草凋落物的混合对Ca、Fe残留率起促进作用。

图7 元素的期望残余率与实测残余率的比较Fig. 7 The difference between the Expected and Observed element remaining
4 讨论
4.1凋落物分解营养元素的变化特征
在本研究中,羊草的N均处于释放状态,冰草N先释放后累积,双物种、多物种凋落物N表现为释放-累积-释放模式,367 d的分解,4类凋落物的N净释放率为26.73%~40.84%。分解过程中元素的释放累积状态与初始元素含量有某种相关关系,初始N含量较低的凋落物容易发生累积现象(涂利华等,2011)。4类凋落物中羊草的初始N最高,处于释放状态,其它物种均出现累积。N累积是微生物分解的一个重要特征,N累积和释放的模式受初始N含量的影响,当凋落物初始N含量不能满足微生物的分解时,凋落物的微生物将外源N转化为自身生物量或是分解酶,导致N累积(Frey et al.,2000;Parton et al.,2007)。N是生物体的组成成分,是生物因子主导型,主要以微生物作用分解为主,元素含量随有机物分解升高或波动(杜占池等,2003),元素累积后又出现释放状态。4类凋落物的P、K、Mg在0~305 d内均为释放模式,而在305~367 d,多物种凋落物持续释放,其它凋落物开始累积,367 d时,4类凋落物P净释放率为47.84%~51.92%,无显著差异(P> 0.05),K净释放率为85.62%~89.66%,Mg净释放率为17.74%~31.41%。P、M g作为植物的重要组成部分,Mg以离子态和螯合态两种状态存在于植物细胞内,离子态的Mg很容易释放,分解过程中,螯合态Mg随螯合物缓慢解体并释放到土壤(马志良等,2015)。Mg动态变化受物理和生物作用双重主导,在分解过程中容易被微生物固定(Berg,1981)。所以,244 d内,离子态Mg大量释放,4类凋落物均为释放模式,随着螯合物的解体,Mg释放速率降低,凋落物出现了累积现象,这与凋落物本身的性质以及微生物作用有关。K是水溶性物质,常以离子状态存在,主要靠淋溶作用进行释放,各营养元素中以K释放的量最多(Bockheim et al.,1986;黄建辉等,1998),且在244 d内因为冻融和融冻的交替,加速凋落物的淋溶作用,K在这阶段释放率最快。多物种凋落物的Ca在0~305 d,均为缓慢释放模式,后有小幅度累积,其它3类凋落物则为累积-释放-累积模式;367 d时,4类凋落物Ca净释放率为11.94%~21.49%。Ca主要以螯合物的形式存在于植物体内,分解前期,螯合物大分子逐渐解体,Ca游离出来,在305~367 d,温度和降雨量的升高,使得Ca从凋落物中释放出来,物种的差异会导致分解速率对温度和降雨产生不同的响应(Whitford et al.,1995),不同物种在各阶段表现出不同的养分释放模式。Mn是木质素降解酶的重要组成部分,对凋落物分解起调节作用(Berg et al.,2010),较高的Mn含量可刺激微生物活动进行木质素的降解(Taylor et al.,1989)。4类凋落物的Mn在0~244 d均为累积模式;244~367 d,多物种凋落物的M n为先累积再释放,其它凋落物Mn先释放再累积;367 d时,羊草、冰草、多物种凋落物为净累积,累积率为35.07%~57.28%,双物种凋落物几乎与初始状态相同,释放量仅为0.9%。4类凋落物Fe在0~244 d均为累积模式,在244~305 d,羊草凋落物的Fe开始释放,其它凋落物则持续累积,在305~367 d,多物种凋落物Fe开始释放,其它凋落物为累积模式,4类凋落物总的累积率为140%~339%,羊草、冰草在367 d达到最大净累积量,另外两类凋落物的净累积量在305 d达到最大。Fe、Mn作为在难分解有机物的组分,微生物很难利用,在初期会先进行累积(王谨等,2001)。Zn、Cu元素作为重要的微量元素,其含量变异较大,各物种在各阶段的表现模式各不相同,这与元素在各凋落物组织中不同的存在形式有关。
4.2基质质量对凋落物分解的影响
在特定区域,凋落物基质质量是影响凋落物分解的主要因子(Zhang et al.,2008)。分解初期的凋落物分解速率与初始的N、P含量以及C/N、C/P等显著相关(Berg,2000),凋落物分解速率与凋落物初始N、P、K、Mg含量呈显著正相关,与凋落物初始C、木质素含量以及木质素/N、木质素/P呈显著负相关(陈法霖等,2011)。木质素是难分解的有机物,木质素含量较高的凋落物分解会受到一定的抑制作用。羊草的初始C/N、木质素/N相对较小,C、N、P、K、Mg、Cu的含量较高,而混合物凋落物初始C/N、木质素/N最大,所以羊草在初始阶段的分解速率较快,混合凋落物的分解速率最慢。其他元素含量在各凋落物间的差异不明显,所以在分解的367 d,各凋落物的质量损失没有显著差异,这也证实初始化学组成对凋落物分解的影响作用。从244 d开始,4类凋落物的C/N、纤维素/N持续下降,原因可能是244 d(5月)开始,温度和降水量明显升高,凋落物开始快速分解。
4.3混合凋落物的非加性效应
物种的非加性效应有多种解释机制。可能是凋落物的形态、大小差异创造多样化的土壤生境,增加食物链的复杂程度和土壤动物的数量,混合凋落物增加了微生物的多样性。同时,不同物种间的化学组成差异使高质量凋落物向低质量凋落物分解的转移(Liu et al.,2007;李强等,2014),缓和分解中营养元素对微生物的限制,而另一方面,凋落物混合对分解环境的改变比凋落物的组成影响更加显著(Hector et al.,2000)。本研究发现,在305 d时,羊草和冰草的混合对质量残留率产生负效应,而后这种负效应减弱,原因可能是羊草和冰草混合的负效应与微环境的变化或者抑制性物质的产生有关。羊草和冰草的混合对P、K、Mg的残留率无显著影响,原因在于水分和温度等其他因素的改变掩盖了混合效应对凋落物分解的影响(陈瑾等,2011)。
5 结论
(1)4类凋落物的初始化学组成各不相同,羊草的C、N、P、K、Mg、Cu和纤维素的含量较高,冰草的Zn含量较高,双物种凋落物的Ca、Fe、Mn、木质素含量较高。羊草的初始C/N、木质素/N最小,分别为28.45、5.19;多物种凋落物初始C/N、木质素/N最大,分别为31.58、6.18。双物种凋落物的初始C/P、纤维素/N最小,分别为458.32、14.72;冰草凋落物的初始C/P、纤维素/N最大,分别为483.36、19.30。4类凋落物分解的质量残留率各不相同,凋落物分解244 d,冰草的质量损失最快,其次是羊草,分解最慢的是多物种凋落物,其中羊草和冰草的质量残留率有显著差异(P<0.05)。在244~305 d的分解过程中,损失最快的是羊草,其次是冰草,多物种分解速率最慢,分解305 d后,冰草、双物种、多物种凋落物之间质量残留率有显著差异(P<0.05)。305~367 d,多物种凋落物质量损失速率最快,其次是双物种凋落物。凋落物在分解的367 d内,分解速率或快或慢,在367 d时质量残留率无显著差异,质量残留率在59%左右。4类凋落物在分解的初期(244 d),质量损失在16.24%~20.11%。
(2)利用负指数衰减模型预测各凋落物的分解过程,0~367 d,分解常数由大到小依次为多物种、羊草、双物种、冰草凋落物。4类凋落物分解95%所需时间是2.88~3.15a。0~307 d,4类凋落物各元素表现为不同的累积释放模式,最终,C、P、K、Mg、Ca、Cu元素为净释放,平均释放率维持在16.37%~87.43%,K、Cu释放率较大,超过50%,C、N的释放率较小,仅在20%左右;Fe为累积模式,累积率为140%~339%。不同凋落物Mn、Zn释放-累积模式不同,羊草、冰草、多物种凋落物的M n为净累积,累积率为35.07%~57.28%,双物种凋落物为净释放,释放率仅为0.9%。羊草凋落物的Zn为净累积,累积率为70.76%,其它凋落物表现为净释放,释放率为4.09%~22.32%。
(3)凋落物的混合存在非加性效应。分解的305 d内,羊草和冰草凋落物之间存在着抑制作用。羊草和冰草凋落物的混合对C、Cu残留率起抑制作用,对Mn、Zn、Ca、Fe残留率起促进作用,对N残留率先起促进作用而后抑制凋落物的N残留率,对P、Mg、K的残留率无显著影响。
AERTS R. 1997. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship [J]. Oikos,79(3):439-449.
AERTS R. 2006. The freezer defrosting: global warming and litter decomposition rates in cold biomes [J]. Journal of Ecology, 94(4):713-724.
ARAUJO P I, YAHDJIAN L, AUSTIN A T. 2012. Do soil organisms affect aboveground litter decomposition in the semiarid Patagonian steppe,Argentina? [J]. Oecologia, 168(1): 221-230.
BERG B, DAVEY M P, DE MARCO A, et al. 2010. Factors influencing limit values for pine needle litter decomposition: a synthesis for boreal and temperate pine forest systems [J]. Biogeochemistry, 100(1-3):57-73.
BERG B. 1981. Leaching, accumulation and release of nitrogen in decomposing forest litter [J]. Ecological Bulletin, 33: 163-178.
BERG B. 2000. Litter decomposition and organic matter turnover in northern forest soils [J]. Forest ecology and Management, 133(1): 13-22.
BOCKHEIM J G, LEIDE J E. 1986. Litter and forest-floor dynam ics in a Pinus resinosa plantation in Wisconsin [J]. Plant and soil, 96(3):393-406.
CHAPMAN S K, NEWMAN G S, HART S C, et al. 2013. Leaf litter m ixtures alter m icrobial community development: m echanisms for non-additive effects in litter decomposition [J]. PLoS One, 8(4):e62671.
FACELLI J M, PICKETTS T A. 1991. Plant litter: its dynamics and effects on plant community structure [J]. The Botanical Review, 57(1): 1-32.
FREY S D, ELLIOTT E T, PAUSTIAN K, et al. 2000. Fungal translocation as a mechanism for soil nitrogen inputs to surface residue decomposition in a no-tillage agroecosystem [J]. Soil Biology and Biochem istry, 32(5): 689-698.
GARTNER T B, CARDON Z G. 2004. Decomposition dynamics in mixed-species leaf litter[J]. Oikos, 104(2): 230-246.
HECTOR A, BEALE AJ, M INNS A, et al. 2000. Consequences of the reduction of plant diversity for litter decomposition: effects through litter quality and microenvironment [J]. Oikos, 90(2): 357-371.
HOORENS B, AERTS R, STROETENGA M. 2003. Does initial litter chemistry explain litter mixture effects on decomposition? [J]. Oecologia, 137(4): 578-586.
KEMP P R, REYNOLDS J F, VIRGINIA R A, et al. 2003. Decomposition of leaf and root litter of Chihuahuan desert shrubs: effects of three years of summer drought [J]. Journal of Arid Environments, 53(1):21-39.
LIU P, SUN O J, HUANG J, et al. 2007. Nonadditive effects of litter mixtures on decomposition and correlation with initial litter N and P concentrations in grassland plant species of northern China [J]. Biology and Fertility of Soils, 44(1): 211-216.
OLSON J S. 1963. Energy storage and the balance of producers and decomposers in ecological systems [J]. Ecology, 44(2): 322-331.
PALM C A, SANCHEZ P A. 1991. Nitrogen release from the leaves of some tropical legumes as affected by their lignin and polyphenolic contents [J]. Soil biology and biochem istry, 23(1): 83-88.
PARTON W, SILVER W L, BURKE I C, et al. 2007. Global-scale sim ilarities in nitrogen release patterns during long-term decomposition[J]. Science, 315(5810): 361-364.
ROVIRA P, ROVIRA R. 2010. Fitting litter decomposition datasets to mathematical curves: towards a generalised exponential approach [J]. Geoderma, 155(3): 329-343.
TAYLOR B R, PARKINSON D, PARSONS W F J. 1989. Nitrogen and lignin content as predictors of litter decay rates: a m icrocosm test [J]. Ecology, 70(1): 97-104.
VIVANCO L, AUSTIN A T. 2006. Intrinsic effects of species on leaf litter and root decomposition: a comparison of temperate grasses from North and South America [J]. Oecologia, 150(1): 97-107.
VOŘÍŠKOVÁ J, DOBIÁŠOVÁ P, ŠNAJDR J, et al. 2011. Chemical composition of litter affects the growth and enzyme production by the saprotrophic basidiomycete Hypholoma fasciculare [J]. Fungal Ecology,4(6): 417-426.
WHITFORD W G, MARTINEZ-TURANZAS G, MARTINEZ-MEZA E. 1995. Persistence of desertified ecosystems: explanations and implications [J]. Environmental Monitoring and Assessment, 37(1-3):319-332.
ZHANG D, HUI D, LUO Y, et al. 2008. Rates of litter decomposition in terrestrial ecosystems: global patterns and controlling factors [J]. Journal of Plant Ecology, 1(2): 85-93.
陈法霖, 江波, 张凯, 等. 2011. 退化红壤丘陵区森林凋落物初始化学组成与分解速率的关系[J]. 应用生态学报, 22(3): 565-570.
陈瑾, 李扬, 黄建辉. 2011. 内蒙古典型草原4种优势植物凋落物的混合分解研究[J]. 植物生态学报, 35(1): 9-16.
陈佐忠. 1988. 锡林河流域地形与气候概况[C]//陈佐忠. 草原生态系统
研究(第3集). 北京: 科学出版社: 13-22.
杜占池, 樊江文, 钟华平. 2003. 营养元素在红三叶叶片分解过程中的释放动态[J]. 草业科学, 20(7): 12-15.
郭继勋, 孙刚, 姜世成. 2000. 松嫩平原羊草草原凋落物层群落学作用的研究[J]. 植物生态学报, 24(4): 473-476.
黄建辉, 陈灵芝, 韩兴国. 1998. 辽东栎枝条分解过程中几种主要营养元素的变化[J]. 植物生态学报, 22(5): 398-402.
李博, 雍世鹏, 李忠厚. 1988. 锡林河流域植被及其利用[C]//陈佐忠. 草原生态系统研究(第3集). 北京: 科学出版社: 84-183.
李强, 周道玮, 陈笑莹. 2014. 地上枯落物的累积、分解及其在陆地生态系统中的作用[J]. 生态学报, 34(14): 3807-3819.
廖仰南, 赵吉, 刘宝音, 等. 1994. 羊草和大针茅不同物候期植株残体及凋落物分解酶活性的研究[J]. 内蒙古大学学报, 25(1) :81-87.
刘增文, 高文俊, 潘开文, 等. 2006. 枯落物分解研究方法和模型讨论[J].生态学报, 26(6): 1993-2000.
马志良, 高顺, 杨万勤, 等. 2015. 亚热带常绿阔叶林区凋落叶木质素和纤维素在不同雨热季节的降解特征[J]. 生态学杂志, 34(1): 122-129.任来阳, 于澎涛, 刘霞, 等. 2013. 重庆酸雨区马尾松与木荷的叶凋落物分解特征[J].生态环境学报, 22(2): 246-250.
邵玉琴, 赵吉, 杨劼. 2004. 内蒙古皇甫川流域凋落物分解过程中营养元素的变化特征[J]. 水土保持学报, 18(3): 81-84.
涂利华, 胡庭兴, 张健, 等. 2011. 模拟氮沉降对两种竹林不同凋落物组分分解过程养分释放的影响[J]. 生态学报, 31(6): 1547-1557.
汪久文, 蔡蔚祺. 1988. 锡林河流域土壤的发生类型及其性质的研究[C]//陈佐忠. 草原生态系统研究(第3集). 北京: 科学出版社.王谨, 黄建辉. 2001. 暖温带地区主要树种叶片凋落物分解过程中主要元素释放的比较[J]. 植物生态学报, 25(3): 375-380.
王玉万, 徐文玉. 1987. 木质纤维素固体基质发酵物中半纤维素、纤维素和木素的定量分析程序[J]. 微生物学通报, 14(2): 81-84.
薛惠琴, 杭怡琼, 陈谊. 2001. 稻草秸秆中木质素、纤维素测定方法的研讨[J]. 上海畜牧兽医通讯, (2): 15-15.
张彩虹, 张雷明, 刘杏认, 等. 2011. 呼伦贝尔草甸草原优势种贝加尔针茅根系组织和地上部分凋落物的分解[J]. 植物生态学报, 35(11):1156-1166.
张建利, 张文, 高玲苹, 等. 2008. 云南马龙县山地封育草地凋落物分解与氮释放的研究[J]. 草业科学, 25(7): 77-82
赵吉, 廖仰南, 张桂枝. 1992. 羊草和大针茅不同物候期植株的分解及其主要营养元素的转化[[C]//陈佐忠. 草原生态系统研究(第4集).北京: 科学出版社: 161-169.
The Dynamics of Nutrients during Litter Decomposition in Leymus chinensis Grassland in Inner Mongolia, China
MU Yue1,2, GENG Yuanbo1*
1. Insitute of Geographic Sciences and Natural Resources Research, CAS, Beijing 100101, China;2. University of Chinese Academy of Sciences, Beijing 100049, China
Litter decomposition is a key link in the process of nutrient cycling in ecosystem. As the main way to return nutrients to the soil from plants, litter decomposition has great significance on maintaining the balance of ecosystem. The decomposition characteristics and nutrient elements dynamic changes of Leymus chinensis (LC), Agropyron cristatum (AC), Leymus chinensis-Agropyron cristatum (LA), and Multi-species (MT) were studied by using the litterbag method in the Xilin River Basin ,Inner-Mongolia, China. The results were shown as follows: The decomposition constant of MT, LA, LC and AC was 1.04, 0.98, 1.01,0.95 respectively. The elemental dynamic changes of 4 litters were different. C, P, K, Mg, Ca, Cu of all four litter types were net release with 16.37%~87.43% mean release amount, and Fe of all four litter types was accumulated with 140%~339% accumulative amount. Mn of LC, AC, and MT litters were net accumulation, and the accumulative amount was 35.07%~57.28%. The LA litter was net release, and its net release amount was only 0.9%. Zn of the LC litter was net accumulation and the accumulative rate was 70.76%. In addition, the Zn of other litters were net release with 4.09%~22.32% release rate. Non-additive effect existed in mixed litter. From 0 to 305 days, there was a inhibitory effect between the LC litter and AC litter. Mixed litter of the LC and AC litter had inhibitory effects on the residual rate of C and Cu, while had positive effects on Mn, Zn, Ca, Fe remaining. Litter mixing could firstly promote N remaining, and then inhibit it. It had no significant influence on the residul rate of P, Mg, K.
Leymus chinensis grassland; litter decomposition; nutrient elements; non-additive effect
10.16258/j.cnki.1674-5906.2016.07.009
Q945.1; X173
A
1674-5906(2016)07-1154-10
国家自然科学基金项目(41071138)
母悦(1990年生),女(羌族),硕士研究生,研究方向为环境生物地球化学。E-mail: muy.13s@igsnrr.ac.cn *通信作者:耿元波,副研究员,研究方向为环境生物地球化学。E-mail: gengyb@igsnrr.ac.cn
2016-05-02
