Magnetospira sp. QH-2 MamK蛋白的结构和系统进化分析
2016-10-20滕兆洁张文燕陈一然潘红苗杜海舰王明玲
滕兆洁,张文燕,刘 佳,董 逸,陈一然,潘红苗,杜海舰,王明玲,徐 丛,肖 天
sp. QH-2 MamK蛋白的结构和系统进化分析
滕兆洁1, 2, 3,张文燕1, 3,刘 佳1, 2, 3,董 逸1, 3,陈一然1, 3,潘红苗1, 3,杜海舰1, 3,王明玲1, 3,徐 丛1, 2, 3,肖 天1, 3
(1. 中国科学院海洋研究所, 海洋生态与环境科学重点实验室,山东青岛 266071; 2. 中国科学院大学, 北京 100049; 3. 青岛海洋科学与技术国家实验室海洋生态与环境科学功能实验室,山东青岛 266071)
sp. QH-2是一株分离自黄海潮间带的海洋趋磁螺菌, 该菌磁小体排列不整齐, 并且磁小体链内某些区域排列疏松, 推测与其磁小体岛上的编码的MamK蛋白相关。开放阅读框大小为1086 bp, 编码361个氨基酸, MamK蛋白属于NBD_sugar-kinase_HSP70_actin超家族, 为非跨膜蛋白; 此外, 还利用生物信息学工具预测MamK蛋白二级和三级结构。系统进化分析显示MamK蛋白的系统进化和16S rRNA基因的系统进化有一定差异, 推测磁小体岛的获得可能与菌种的进化是独立的两个过程; 淡水与海水中的趋磁螺菌虽然形态相似, 但系统进化地位有一定差异, 可能是两者不同生境下适应性演化的结果。
MamK; 系统进化; 16S rRNA基因; 磁小体岛; 结构
趋磁细菌(Magnetotactic bacteria, MTB)是一类能够沿着磁力线运动的特殊细菌, 最早由意大利学者Bellini在淡水中发现[1-3], 但直到美国学者Blakemore在Science上报道了在海泥中发现的趋磁细菌后, 该类细菌才引起科学界广泛关注[4]。趋磁细菌形态多样, 常见有球形、杆状、弧形、螺旋形及多细胞聚集体形式等[5-8]。目前, 已知的趋磁细菌隶属于α-变形菌纲(Alpha-Proteobacteria)、γ-变形菌纲(Gamma-Proteobacteria)、δ-变形菌纲(Delta-Proteobacteria)、硝化螺菌门(Nitrospirae)、candidate phylum Omnitrophica[9-11]和candidate phylum[12]。
趋磁细菌体内均含有生物膜包裹的磁小体(magnetosome)。趋磁细菌通过磁小体的导向作用, 借助鞭毛进行趋磁运动[13], 使其更有效地找到最适生存环境, 即有氧-无氧过渡区(oxic-anoxic transition zone, OATZ)[14-16]。趋磁细菌的磁小体大小为25~120 nm, 处于稳定的单磁畴晶体范围内[13, 17-18], 由双层磷脂膜包被。磁小体有立方八面体、平行六面体、子弹头形、片或齿状、不规则形状等多种形态[19-21]。大部分趋磁细菌的磁小体在细胞内呈链状排列, 有序排列的磁小体犹如一个微型的生物指南针, 使得该类微生物能够有效感受到外界磁场。磁小体按成分可分为铁氧型(magnetite, Fe3O4)和铁硫型(gregite, Fe3S4)[22], 有的趋磁细菌可同时含有两种不同成分的磁小体。磁小体的特征, 如形态、排列、成分等均具有种属特异性[23]。
磁小体的合成过程受到严格的生物调控, 与磁小体合成相关的基因都位于一个特定的区域, 称为磁小体岛(magnetosome island, MAI), 主要包括、、和四个操纵子[24], 其中是控制磁小体合成的核心操纵子[25-27]。
本文研究的基因位于操纵子。MamK蛋白在细胞内形成丝状的细胞骨架结构(Magnetosome filament, MF), 磁小体沿着此结构紧密排列成链并锚定在细胞中[28]。MamK蛋白在磁小体链的排列与稳定功能中扮演重要角色[29-30]。
sp. QH-2 (QH-2)[31]是本实验室分离自黄海潮间带的一株海洋趋磁螺菌, 菌体长约3 μm, 宽约0.8 μm左右。其体内含有铁氧型(Fe3O4)磁小体, 7~28个磁小体排列成一条链。QH-2的磁小体大小不均一, 与其他趋磁螺菌相比, 其磁小体链中某些区域间隙较大, 且菌体运动较慢, 速度为30~50 μm/s。QH-2基因组包括4.0 Mb的拟核和31.0 Kb的质粒[32]。该菌属于α-变形菌纲的趋磁螺菌属, 与海水分离的趋磁螺菌MMS-1 (MMS-1)差异性仅为2.8%[33], 而与淡水分离的趋磁螺菌AMB-1 (AMB-1)[34-35]和MSR-1 (MSR-1)序列差异性超过11%[34]。
研究表明MamK蛋白与磁小体链的排列和稳定密切相关[28-30], 因此QH-2的磁小体排列疏松、运动速度较慢等特性可能与该蛋白有关。基于已经获得的基因组数据, 本研究将通过分析QH-2的MamK的理化性质、二级结构和三级结构并在三级结构的基础上进行空间结构的同源建模, 预测在QH-2磁小体合成和排列中可能的功能与作用, 为进一步研究在QH-2中的作用奠定基础; 同时分析、、16Sr RNA基因的系统进化地位, 以探索磁小体岛的进化和起源。
1 分析方法
1.1 MamK蛋白的理化性质分析
将获得的趋磁螺菌QH-2的基因组序列, 在MaGe (https: //www.genoscope.cns.fr)平台进行同源比对, 找到基因, 用于MamK蛋白的理化性质和结构进行预测分析。利用蛋白分析专家系统(Expert Protein Analysis System, ExPASy)的ProtParam tool (http: //ca.expasy.org/tools/protparam/ html)以及ProtScale((http: //web.expasy.org/protscale/)分析MamK蛋白的氨基酸组成、相对分子质量、等电点、不稳定性和亲/疏水性等理化性质。
1.2 MamK蛋白的高级结构分析及同源建模
利用GOR4(https: //npsa-prabi.ibcp.fr/cgi-bin/ npsa_automat.pl?page=/NPSA/npsa_gor4.html)分析MamK蛋白的二级结构, 包括α-螺旋、β-转角、无规则卷曲及延伸链等; MamK蛋白的跨膜区域分析由HMMTOP(http: //hmmtop.enzim.hu/)系统完成; MamK蛋白三级结构分析通过Phyre2 (http: // www.sbg.bio. ic.ac.uk/phyre2/html/page.cgi?Id=index)进行, 序列提交后, 用同源建模的方法获得MamK蛋白的三级结构, 并进行评价。
1.3 QH-2 MamK蛋白系统进化分析
利用NCBI BLAST工具中blastp程序(http: // blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp& PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)对MamK蛋白进行同源比对, 同时在NCBI保守功能域数据库(Conserved domains Database)v.3.11中寻找该蛋白结构中的超家族关系及保守区域。
在NCBI GenBank (http: //www.ncbi.nlm.nih.gov/ genbank/)数据库中检索相关物种的16Sr RNA基因。利用Clustal W对QH-2 MamK蛋白及同源序列、QH-2的16S rRNA基因序列及相关物种16S rRNA基因序列进行比对, 比对后利用MEGA 6.0软件, 采用Neighbor-joining的方法构建系统进化树, Bootstrap为1000, 分析两者的系统进化关系。
2 结果分析
2.1 MamK蛋白理化性质分析
基因大小为1086 bp, 编码的MamK蛋白由361个氨基酸组成, 原子总数为5462个, 分子式为C170H2763N465O520S13, 分子质量为38.4652 kD, 理论等电点为4.95。在所有氨基酸中, 丙氨酸(Ala)含量最高为11.6%, 缬氨酸(Val)次之为10%, 氨基酸组成见图1。该蛋白包含36个碱性氨基酸, 50个酸性氨基酸, 145个疏水性氨基酸和62个极性氨基酸。带正电的氨基酸残基(Arg + Lys)共45个, 负电残基(Asp + Glu)共49个, 不稳定系数(instability index, II)为27.61, 总的疏水性平均值(Grand average of hydropathicity, GRAVY)为0.057。Protscale分析显示QH-2的MamK蛋白为疏水性蛋白, 分析结果见图2。
2.2 QH-2 MamK蛋白高级结构分析
利用GOR4对QH-2 MamK蛋白进行二级结构的预测, 结果(见图3)表明, α螺旋(Hh, 41%)和不规则卷曲(Cc, 44.6%)是MamK蛋白的主要组成结构元件, 有利于稳定蛋白质的结构, 延伸链(Ee)结构占14.4%, 且该蛋白无β-转角结构。HMMTOP对该蛋白进行跨膜结构域的预测结果显示, MamK蛋白不存在跨膜区。
利用Phyr2在线工具, 以c1o1f4为模型建模预测的蛋白三级结构见图4。
2.3 QH-2 MamK蛋白系统进化分析
Blast结果(表1)显示QH-2的MamK蛋白在QH-2中还存在一个MamK-like蛋白, 同源性100%, 但序列覆盖率仅为89%。除此之外, MamK蛋白与actin-like蛋白相似性较高, 与其他趋磁细菌的MamK蛋白同源性最高达66%, 而与细胞形状维持蛋白MreB相似性最高为41%。同时在NCBI保守功能域数据库的检索结果显示, QH-2 MamK蛋白是MreB-like蛋白, 属于NBD_sugar-kinase_HSP70_ actin超蛋白家族。
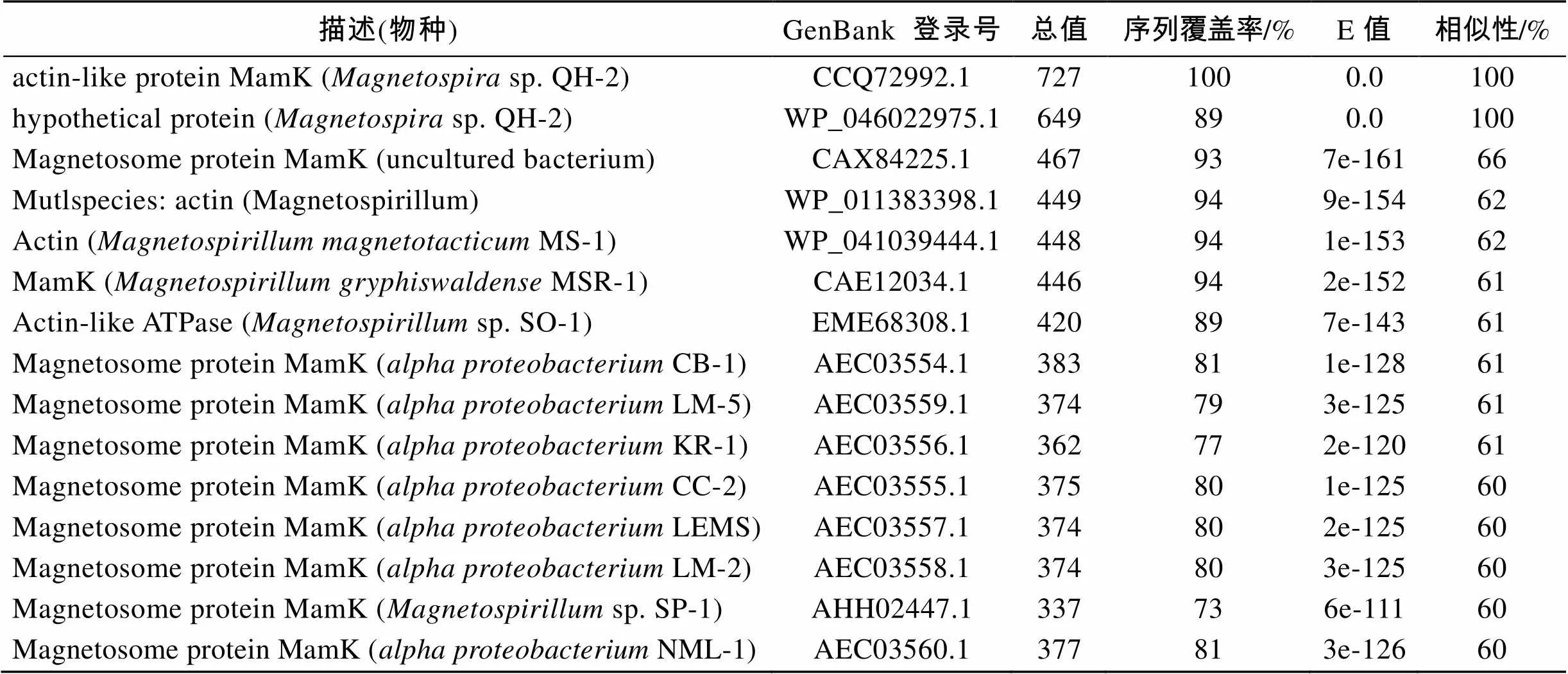
表1 QH-2 MamK蛋白Blast结果
选择MamK蛋白同源性较高的序列27条, 同时检索相关物种的16S rRNA基因序列23条, 序列比对后分别构建系统进化树。MamK蛋白的系统进化分析(见图5)显示, 这些物种的MamK蛋白分为明显的两大支, 其中来源于α-变形菌纲和γ-变形菌纲的趋磁细菌MamK蛋白聚为一类, 而δ-变形菌纲、硝化螺菌门以及candidate phylum Omnitrophica的趋磁细菌MamK蛋白聚在一起。MamK蛋白的系统进化和16S rRNA基因的系统进化(见图6)有差异, 尤其是δ-变形菌纲、硝化螺菌门以及candidate phylum The numbers in parenthesis are the GenBank accession numbers, and the MamKprotein sequences of QH-2 are shown in boldOmnitrophica, 两者差异明显; 而MamK蛋白和16S rRNA基因的系统进化在α-变形菌纲中保持高度的一致性。QH-2与strain MV-1 (MV-1)中MamK蛋白的序列最为接近, 同16S rRNA基因的系统进化一致。MamK和16S rRNA基因序列的系统进化均显示, QH-2代表的海洋趋磁螺菌属与淡水趋磁螺菌属的分支距离较远, 同源性较低, 而与MV-1代表的海洋趋磁弧菌属更为接近。
括号内为GenBank登录号, 加粗字体显示QH-2 16S rRNA基因序列
The numbers in parenthesis are the GenBank accession numbers, and the 16S rRNA gene sequences of QH-2 are shown in bold
括号内为GenBank登录号, 加粗字体显示QH-2 Mamk蛋白序列
The numbers in parenthesis are the GenBank accession numbers, and the MamKprotein sequences of QH-2 are shown in bold
3 讨论
趋磁细菌磁小体岛含有多个基因可能与磁小体多链结构以及磁小体复杂排列有关(除趋磁螺菌和MC-1)[36]。与QH-2 MamK蛋白同源性最高的蛋白来自于QH-2本身, 序列覆盖率为89%, 同源性100%, QH-2中可能存在MamK的同源蛋白MamK- like蛋白, 但它在QH-2菌体内行使何种功能, 是否与MamK蛋白功能一致, 两个MamK的存在是否与QH-2大小不一磁小体的排列复杂有关, 这些问题还有待于进一步实验确认。Arash Komeili等曾报道在AMB-1中发现了一种MamK同源蛋白MamK-like, 它能与MamK直接作用, 调节相关ATPase活性, 从而影响磁小体链合成进程[37]。AMB-1为一株淡水趋磁螺菌, 其磁小体颗粒平均大小为50 nm, 15个以上磁小体排列成一条链, 但与之相比, QH-2磁小体链排列更为疏松[38]。本文原拟通过分析QH-2 MamK蛋白的理化性质解释磁小体排列疏松、菌体运动速度较慢等特性, 但结果显示QH-2的这些特性可能不仅与MamK蛋白相关, 也与其他磁小体岛基因的表达调控有关。以上推测仅由分析获得, 具体原因机理将通过后续实验加以验证。
MamK蛋白与MreB蛋白同源, 均为actin-like蛋白。MreB蛋白在细菌中普遍存在, 其主要功能与细胞形状决定、细胞极性构建, 以及染色体分离有关[39];而趋磁细菌的MamK蛋白主要在磁小体链的组装, 成链以及空间定位的过程中起作用, 但两者功能的行使都有赖于丝状结构的形成[25]。此外, MamK蛋白可能与MreB蛋白一样行使分子马达功能[40], 在磁小体链排列过程中, 将新合成的磁小体晶体转运到正在生长的磁小体链的末端。
MamK蛋白的系统进化分析显示, 来源于α-变形菌纲和γ-变形菌纲的趋磁细菌MamK蛋白与δ-变形菌纲、硝化螺菌门以及candidate division OP3的趋磁细菌MamK蛋白形成区分明显的两大支, 两者的MamK蛋白可能具有不同的起源, 暗示磁小体岛的起源可能不同; 同时, MamK蛋白的系统进化和16S rRNA基因的系统进化有差异, 尤其是δ-变形菌纲、硝化螺菌门以及candidate division OP3, 两者差异明显, 在这些分类单元16S rRNA基因和MamK蛋白的进化并不一致, 由此推测磁小体岛的获得可能与菌种的进化是独立的两个过程, 这些进化地位不同的趋磁细菌获得了相同或相近来源的磁小体岛, 从而获得趋磁性, 磁小体岛的获得很可能是在系统进化之后。而MamK蛋白和16S rRNA基因的系统进化在α-变形菌纲中保持高度的一致性, 因此这些趋磁细菌可能是由具有磁小体岛的同一祖先进化而来。与QH-2中Mamk蛋白的同源性最近的MreB蛋白来源于浮霉菌门的Phycisphaera mikurensis, 同源性为41%, 远低于与趋磁细菌MamK蛋白的同源性(66%)。QH-2 MamK蛋白与MreB蛋白序列的相似性仅为24.9% (CCQ72944.1和WP_046020642.1), 两者同源性很低, 表明两者相互进化而来的可能性极低。与淡水趋磁螺菌属趋磁细菌相比, QH-2与MV-1(海洋趋磁弧菌)更为接近。淡水的趋磁螺菌属和海洋的趋磁螺菌属虽然形态类似, 但生境不同, 在α-变形菌纲中系统进化地位也不同, 两者不同的生境造就了它们之间的差异, 这是趋磁细菌长期适应性演化的结果, 也可能意味着海洋和淡水的螺菌有着不同的来源。
[1] Bellini S. On a unique behavior of freshwater bacteria[J]. Chin J Oceanol Limnol, 2009, 27: 3-5.
[2] Bellini S. Further studies on “magnetosensitive bacteria”[J]. Chin J Oceanol Limnol, 2009, 27: 6-12.
[3] Frankel RB. The discovery of magnetotactic/magnetosensitive bacteria[J]. Chin J Oceanol Limnol, 2009, 27: 1-2.
[4] Blakemore R. Magnetotactic bacteria[J]. Science, 1975, 190: 377-379.
[5] Lefevre C T, Bernadac A, Yu-Zhang K, et al. Isolation and characterization of a magnetotactic bacterial culture from the mediterranean sea[J]. Environmental Microbiology, 2009, 11: 1646-1657.
[6] Maratea D, Blakemore R P. Aquaspirillum-magnetotacticum sp-nov, a magnetic spirillum[J]. International Journal Of Systematic Bacteriology, 1981, 31: 452-455.
[7] Sakaguchi T, Arakaki A, Matsunaga T. Desulfovibrio magneticus sp nov., a novel sulfate-reducing bacterium that produces intracellular single-domain-sized magnetite particles[J]. International Journal Of Systematic And Evolutionary Microbiology, 2002, 52: 215-221.
[8] Wenter R, Wanner G, Schueler D, et al. Ultrastructure, tactic behaviour and potential for sulfate reduction of a novel multicellular magnetotactic prokaryote from north sea sediments[J]. Environmental Microbiology, 2009, 11: 1493-1505.
[9] Kolinko S, Wanner G, Katzmann E, et al. Clone libraries and single cell genome amplification reveal extended diversity of uncultivated magnetotactic bacteria from marine and freshwater environments[J]. Environmental Microbiology, 2013, 15: 1290-1301.
[10] Lefevre C T, Bazylinski D A. Ecology, diversity, and evolution of magnetotactic bacteria[J]. Microbiology And Molecular Biology Reviews, 2013, 77: 497-526.
[11] Kolinko S, Richter M, Glöckner F-O, et al. Single-cell genomics of uncultivated deep-branching magnetotactic bacteria reveals a conserved set of magnetosome genes. Environmental microbiology[J], 2016, 18: 21-37.
[12] Lin W, Pan Y. A putative greigite-type magnetosome gene cluster from the candidate phylum latescibacteria[J]. Environmental Microbiology Reports, 2015, 7: 237-242.
[13] Bazylinski D A, Frankel R B. Magnetosome formation in prokaryotes[J]. Nature Reviews Microbiology, 2004, 2: 217-230.
[14] Frankel R B, Bazylinski D A. Magnetosomes and magneto-aerotaxis[J]. Contributions to microbiology, 2009, 16: 182-193.
[15] Frankel R B, Bazylinski D A, Johnson M S, et al. Magneto-aerotaxis in marine coccoid bacteria[J]. Biophysical Journal, 1997, 73: 994-1000.
[16] Simmons S L, Sievert S M, Frankel R B, et al. Spatiotemporal distribution of marine magnetotactic bacteria in a seasonally stratified coastal salt pond[J]. Applied And Environmental Microbiology, 2004, 70: 6230- 6239.
[17] Farina M, Debarros H L, Esquivel D M S, et al. Ultrastructure of a magnetotactic microorganism[J]. Biology Of the Cell, 1983, 48: 85-88.
[18] Bazylinski D A, Moskowitz B M. Microbial biomineralization of magnetic iron minerals: microbiology, magnetism and environmental significance[J]. Rev Mineral Geochem, 1997, 35: 181-223.
[19] Bazylinski D A, Moskowitz B M. Microbial biomineralization of magnetic iron minerals; microbiology, magnetism and environmental significance[J]. Reviews in Mineralogy and Geochemistry, 1997, 35: 181-223.
[20] Schüler D. Formation of magnetosomes in magnetotactic bacteria[J]. Journal of Molecular Microbiology and Biotechnology, 1999, 1(1): 79-86.
[21] 高峻, 潘红苗, 吴龙飞, 等. 黄海冷水团附近沉积物中的趋磁细菌及磁小体的特性研究[J]. 海洋科学, 2006, 30: 11-16. Gao Jun, Pan Hongmiao, Wu Longfei , et al. 黄海冷水团附近沉积物中的趋磁细菌及磁小体的特性研究[J]. Marine Sciences, 2006, 30: 11-16.
[22] Bazylinski D A, Lefèvre C T, Schüler D. Magnetotactic bacteria[M] . Rosenberg E, DeLong E F, Lory S, Stackebrandt E, Thompson F eds. The Prokaryotes. Heidelberg: Springer, 2013: 842-862.
[23] Benzerara K, Menguy N. Looking for traces of life in minerals[J]. Comptes Rendus Palevol, 2009, 8: 617- 628.
[24] Yoshino T, Matsunaga T. Development of efficient expression system for protein display on bacterial magnetic particles[J]. Biochemical And Biophysical Research Communications, 2005, 338: 1678-1681.
[25] Komeili A, Li Z, Newman D K, et al. Magnetosomes are cell membrane invaginations organized by the actin-like protein mamk[J]. Science, 2006, 311: 242-245.
[26] Komeili A, Vali H, Beveridge T J, et al. Magnetosome vesicles are present before magnetite formation, and mama is required for their activation[J]. Proceedings Of the National Academy Of Sciences Of the United States Of America, 2004, 101: 3839-3844.
[27] Scheffel A, Gruska M, Faivre D, et al. An acidic protein aligns magnetosomes along a filamentous structure in magnetotactic bacteria[J]. Nature, 2006, 440: 110-114.
[28] Frankel R B, Bazylinski D A. How magnetotactic bacteria make magnetosomes queue up[J]. Trends In Microbiology, 2006, 14: 329-331.
[29] Schuler D. Molecular analysis of a subcellular compartment: The magnetosome membrane in magnetospirillum gryphiswaldense[J]. Archives Of Microbiology, 2004, 181: 1-7.
[30] Pradel N, Santini C L, Bernadac A, et al. Biogenesis of actin-like bacterial cytoskeletal filaments destined for positioning prokaryotic magnetic organelles[J]. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103: 17485-17489.
[31] Zhu K, Pan H, Li J, et al. Isolation and characterization of a marine magnetotactic spirillum axenic culture qh-2 from an intertidal zone of the china sea[J]. Research In Microbiology, 2010, 161: 276-283.
[32] Ji B, Zhang S D, Arnoux P, et al. Comparative genomic analysis provides insights into the evolution and niche adaptation of marine magnetospira sp qh-2 strain[J]. Environmental Microbiology, 2014, 16: 525-544.
[33] Williams T J, Lefevre C T, Zhao W D, et al. Magnetospira thiophila gen. Nov., sp nov., a marine magnetotactic bacterium that represents a novel lineage within the rhodospirillaceae (alphaproteobacteria)[J]. International Journal of Systematic and Evolutionary Microbiology, 2012, 62: 2443-2450.
[34] Wang X, Wang Q, Zhang W, et al. Complete genome sequence of magnetospirillum gryphiswaldense msr-1[J]. Genome announcements, 2014, 2: e00171-14.
[35] Matsunaga T, Okamura Y, Fukuda Y, et al. Complete genome sequence of the facultative anaerobic magnetotactic bacterium magnetospirillum sp strain amb-1[J]. DNA Research, 2005, 12: 157-166.
[36] Jogler C, Lin W, Meyerdierks A, et al. Toward cloning of the magnetotactic metagenome: Identification of magnetosome island gene clusters in uncultivated magnetotactic bacteria from different aquatic sediments[J]. Applied and Environmental Microbiology, 2009, 75: 3972-3979.
[37] Abreu N, Mannoubi S, Ozyamak E, et al. Interplay between two bacterial actin homologs, mamk and mamk-like, is required for the alignment of magnetosome organelles in magnetospirillum magneticum amb-1[J]. Journal of Bacteriology, 2014, 196: 3111- 3121.
[38] Matsunaga T, Sakaguchi T, Tadokoro F. Magnetite formation by a magnetic bacterium capable of growing aerobically[J]. Appl Microbiol Biotechnol, 1991, 35: 651-655.
[39] Gitai Z. Diversification and specialization of the bacterial cytoskeleton[J]. Current Opinion in Cell Biology, 2007, 19: 5-12.
[40] Thanbichler M, Shapiro L. Getting organized - how bacterial cells move proteins and DNA[J]. Nature Reviews Microbiology, 2008, 6: 28-40.
Insight into the structure and evolution of the actin-like protein MamK insp. QH-2
TENG Zhao-jie1, 2, 3, ZHANG Wen-yan1, 3, LIU Jia1, 2, 3, DONG Yi1, 3, CHEN Yi-yan1, 3,PAN Hong-miao1, 3, DU Hai-jian1, 3, WANG Ming-ling1, 3, XU Cong1, 2, 3, XIAO Tian1, 3
(1. Key Laboratory of Marine Ecology & Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Laboratory of Marine Ecology and Environmental Science, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266071, China)Received:Nov. 3, 2015
MamK; phylogenesis; 16S rRNA gene; Magnetosome Island; structure
sp. QH-2 is a marine magnetotactic spirillum, isolated from the intertidal zone of the Yellow Sea, China. The trait that its magnetosome chains display a relatively large size distribution is predicted to be related to the function of the mamK-encoded protein, namely, MamK. We find that mamK gene codes for 1086bp and MamK, which belong to the NBD_sugar-kinase_HSP70_actin superfamily, are composed of 361 amino acids and do not contain any trans-membrane domain. Based on the differences of the phylogenetic trees, we determine that the acquisition of magnetosome island and bacteria evolution are possibly two independent processes. While the morphology feature of magnetotactic spirillum both in fresh water and marine is similar, its phylogenetic status is different to some extent, which may indicate its adaptive evolutions.
Q811.4
A
1000-3096(2016)07-0001-07
10.11759/hykx20151103001
2015-11-03;
2015-12-31;
国家自然科学基金(41206150, 41276170, 41330962)
[Foundation: National Natural Science Foundation of China, No. 41206150, 41276170, 41330962]
滕兆洁(1991-), 女, 山东青岛人, 硕士研究生, 从事海洋微生物生态学研究, 电话: 0532-82898584, E-mail: tengpaper@163.com;肖天,通信作者, 研究员, 博士生导师, 电话: 0532-82898586, E-mail: txiao@qdio.ac.cn
(本文编辑: 康亦兼)
