粤西近岸海域浮游植物群落的生态特征
2016-10-20林华剑徐姗楠龚玉艳李纯厚
林华剑, 徐姗楠, 刘 永, 龚玉艳, 李纯厚
粤西近岸海域浮游植物群落的生态特征
林华剑, 徐姗楠, 刘 永, 龚玉艳, 李纯厚
(中国水产科学研究院南海水产研究所, 农业部南海渔业资源开发利用重点实验室, 广东省渔业生态环境重点实验室, 广州 510300)
作者基于2011年9月(丰水期)和2012年1月(枯水期)粤西近岸30米以浅海域的调查资料, 分析该海域浮游植物的种类组成、优势种、丰度及其群落结构多样性。结果表明, 调查海域共鉴定浮游植物4门41属116种, 硅藻为最主要优势类群。丰水期与枯水期共有种类40种, 物种相似性指数为34.48%, 表明2个调查季节种类组成变化较大, 存在明显的季节性差异。枯水期优势种仅1种, 为金藻门()的球形棕囊藻(), 优势度高达0.496; 丰水期优势种有4种, 其中以硅藻门的旋链角毛藻()优势度较高(0.249)。浮游植物丰度表现出时间和空间异质性, 丰度总体呈现近岸站位高于离岸站位, 此外, 丰水期浮游植物丰度湛江沿岸站位高于其他调查站位, 枯水期浮游植物丰度则由西南部向东北部递减, 总体而言, 枯水期浮游植物丰度比丰水期高。群落结构多样性为枯水期西南部海域明显低于东北部海域, 丰水期站位间多样性无明显变化规律, 相比之下, 丰水期群落物种多样性水平相对较高。相关分析表明: 浮游植物丰度与pH、悬浮物呈正相关, 与丰水期水温和枯水期盐度呈负相关。
粤西近岸海域; 浮游植物; 丰度; 群落结构
海洋浮游植物作为海洋动物, 尤其是海洋动物幼体的直接或间接饵料是海洋生物生产力的基础, 其种类组成和数量变化会影响整个食物链的物质循环、能量流动和渔业资源的波动, 从而引起海洋生态系统的变化。浮游植物对水体条件变化响应灵敏, 是水质监测的重要生物类群, 其种类组成、数量分布及密度的季节变化是其群落结构动态的重要特征, 也是判断水体富营养状态的关键指标之一[1-3]。研究表明, 海洋浮游植物群落不仅可以迅速而灵敏地反映环境的变化[4-6], 而且浮游植物群落结构、生物量及生产力的改变会在碳通量、云反照率和海水光通量与热通量上改变着全球气候[7]。因此, 研究海洋浮游植物群落结构特征是研究海区其他生态和环境问题的基础, 具有重要意义。
粤西海域地处热带、亚热带, 是多种经济鱼类产卵、索饵、繁殖和洄游的重要场所, 粤西及海南岛东北部渔场就坐落于此。粤西沿岸海域地处陆源众多江河入海口, 深受人类养殖业、工业排污等活动影响,是海洋冲淡水的主要影响区域, 其水域生态环境变化, 尤其是作为海洋第一生产力的浮游植物群落结构的变化与人类关系密切, 因此, 有必要对该海域浮游植物进行系统研究。近几十年来, 虽然在南海北部海域进行过多次大规模的海洋勘测和调查[8-10], 但对该海域尤其是粤西海域浮游植物的系统性调查资料甚少。因此, 作者通过2011年9月(丰水期)和2012年1月(枯水期)的调查资料, 对粤西沿岸30 m以浅海域浮游植物的种类组成、密度变化及多样性水平进行分析, 为深入研究该海域的生产力水平、生态系统结构和环境质量状况提供基础资料。
1 材料与方法
分别于2011年9月3~5日(丰水期)和2012年1月23~25日(枯水期)在粤西近岸海域30 m以浅(南至琼州海峡, 北至阳江以南近岸海域; 110°2′0″~ 111°18′0″E, 20°12′0″~21°25′0″N)乘“北渔60011”渔船进行浮游植物样品采集, 调查海域共设12个调查站点(图1)。样品的采集、固定、计数和数据处理均按《海洋调查规范》[11]执行, 采用浅水Ⅲ型浮游生物网(网长280 cm, 筛绢JP80, 网口内径37 cm, 网目孔径0.077 mm, 网口面积0.1 m2)自海底0.5 m至海面垂直拖网。样品用甲醛溶液固定和保存, 浓度为5%。固定样品带至实验室后水样静置48 h, 吸去上清液, 根据细胞丰度大小浓缩至50或100 mL, 取0.1 mL均匀样品在光学显微镜下进行种类鉴定和细胞计数, 每次计数3次, 取其平均值。同时采集表层和底层水样, 测定其温度、盐度、pH、溶解氧、CODMn、BOD5、溶解无机氮、活性磷酸盐、活性硅酸盐、总悬浮物, 并测定水深和透明度。分析时环境数据均采用表层和底层的平均值。
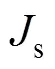
﹦
物种多样性指数的计算采用Shannon-Wiener多样性指数[14]和Margalef指数[15]的计算公式:
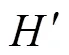
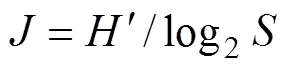
通过系统聚类法分析浮游植物群落结构, 聚类分析以站位间浮游植物Bray-Curtis相异测度系数[17-18]为距离指标, 采用非加权平均配对法(UPGA)在Statistics统计软件上完成[19]。采用SPSS13.0统计分析软件对浮游植物丰度与环境因子进行Pearson相关分析。
2 结果
2.1 种类组成及季节变化
本次调查共鉴定出硅藻门(Bacillariophyta)、甲藻门(Pyrrophyta)、蓝藻门(Cyanophyta)和金藻门(Chrysophyta)4大门类, 共41属116种(包括变种和变形), 其中, 硅藻最多, 有25属86种, 占总种数的74.14%; 甲藻次之, 有13属25种, 占总种数的21.55%; 蓝藻有2属4种, 占总种数的3.44%, 金藻有1属1种, 占总种数的0.86%。硅藻门以角毛藻属()种类最多(26种), 占硅藻总种数的30.23%, 其次为圆筛藻属()(14种)和根管藻属()(12种), 分别占硅藻总种数的16.24%和13.95%。甲藻门以角藻属()为主(10种), 占甲藻总种数的40.00%。在丰水期和枯水期, 浮游植物分别有34属84种和24属72种, 丰水期种数比枯水期多, 其中共有物种40种, 物种相似性指数为34.48%, 表明2个调查季节种类组成变化较大, 存在明显的季节性差异。丰水期和枯水期均表现为硅藻门种类最多, 甲藻门次之, 蓝藻门和金藻门最少。浮游植物种类组成及分布分别见图2和表1。
a. 丰水期; b. 枯水期
a. wet season; b. dry season
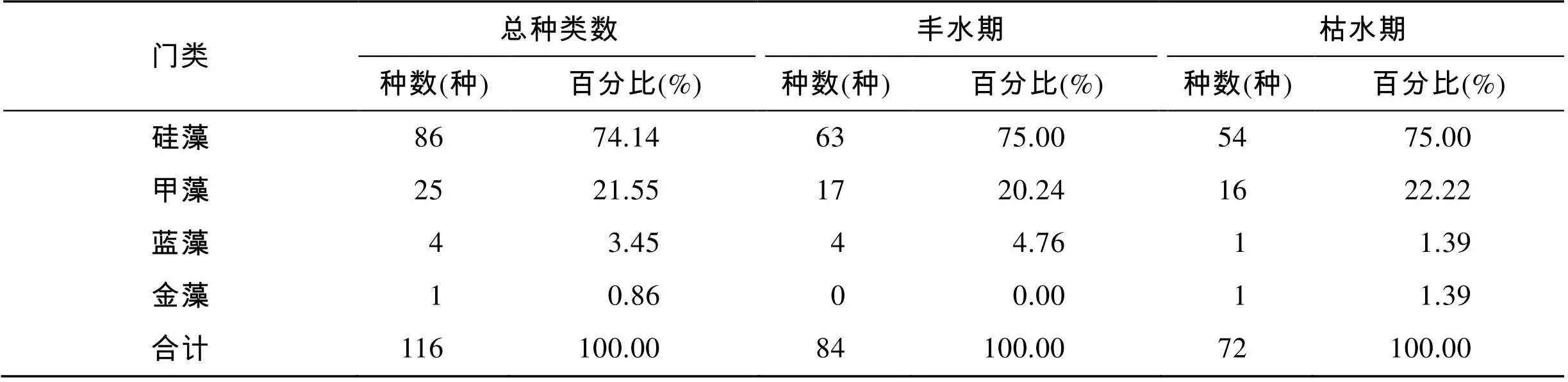
表1 粤西近岸海域浮游植物种类组成
2.2 浮游植物丰度分布
丰水期浮游植物丰度范围为(0.05×104~35.20× 104)个/L, 平均为7.14×104个/L; 主要密集分布于湛江湾附近海域, 以S3站最高, S12站最低, 丰度变化呈现近岸站位高于离岸站位, 浅水站位高于深水站位(图3a)。枯水期浮游植物细胞丰度范围为(0.52× 104~3 540.00×104)个/L, 平均丰度为315.62×104个/L;最高值出现在S2站, 最低值出现在S9站, 丰度由调查海域西南部向东北部递减。部分调查站位(S1~S5, S7)球形棕囊藻大量繁殖, 导致浮游植物丰度急剧增大, 尤其是S2站位, 球形棕囊藻细胞丰度高达3532.50×104个/L(图3b), 已造成球形棕壤藻赤潮。除个别站位(S3、S9)外, 枯水期浮游植物丰度明显高于丰水期。
a. 丰水期; b. 枯水期
a. wet season; b. dry season
2.3 优势种组成及季节变化
调查海域浮游植物优势种表现出明显的季节差异和平面分布差异(表2)。丰水期优势种有4种, 分别为旋链角毛藻柔弱菱形藻()、透明辐杆藻()和中肋角毛藻(), 其中以硅藻门的旋链角毛藻优势度较高(0.249)。丰水期优势种平面分布差异明显, 湛江湾站位(S1~S6)以旋链角毛藻和柔弱菱形藻作为优势种, 优势度相对较高; 到茂名市近岸站位(S7~S12), 旋链角毛藻和柔弱菱形藻数量减少, 透明辐杆藻数量增多, 并成为该海域的优势种。枯水期优势种仅1种, 为球形棕囊藻, 优势度高达0.496, 主要分布在南部调查站位(S1~S5, S7), 丰度比例高达99.238%; 北部调查站位(S6, S9~S12)并未出现球形棕囊藻, 而主要以硅藻门种类为主, 但未见明显的优势种类。
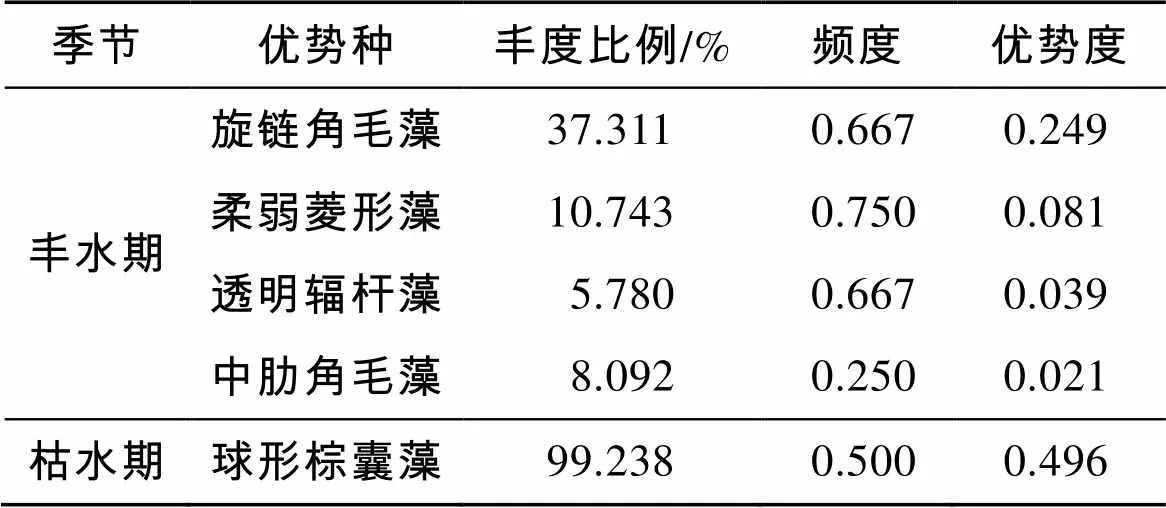
表2 粤西近岸海域浮游植物优势种组成及优势度
2.4 群落结构
聚类分析结果(图4)显示, 调查海域不同季节间浮游植物群落结构差异明显。丰水期分为4个生态区, 湛江湾西南中部生态区(S3、S4)、茂名湛江湾东北近岸生态区(S5、S7、S9)、由北面站位(S8、S10~S12)构成的茂名阳江生态区和西南面站位(S1、S2)组成的湛江湾西南部生态区(图4a)。枯水期研究海域亦表现出明显的区域生态现象, 研究海域受区域影响较大, 整体研究海域可分为3个生态区, 由S1、S2站位构成的湛江湾西南生态区, 由S3~S4、S5、S7站位构成的湛江湾中部生态区, 由S6、S8~S12站位构成的茂名阳江生态区(图4b)。聚类分析结果与浮游植物丰度空间分布(图3)对应较好: 密度高值区和密度低值区分别处于不同生态区, 相对差异性较大; 高值区与低值区之间的过渡区构成新的生态区。
2.5 群落多样性
粤西近岸海域丰水期的香浓维纳(Shannon- wiener) 指数、Margalef指数、Pielou均匀度指数变化范围分别为1.734~3.956、18.994~50.634、0.484~ 0.904, 均值分别为2.964、31.591、0.712; 枯水期的Shannon-wiener指数、Margalef指数、Pielou均匀度指数变幅分别为0.020~3.707、10.842~26.126、0.005~0.788, 均值分别为1.574、19.248、0.372(图5)。与枯水期相比较, 相同站位(除S9和S10的Margalef指数外)丰水期的群落物种多样性水平较高; 枯水期群落物种多样性西南部站位明显低于东北部站位。
2.6 浮游植物丰度与环境因子的关系
将各站浮游植物丰度与水温、盐度、溶解氧、pH、化学需氧量、生化需氧量、活性磷酸盐、活性硅酸盐、无机氮和悬浮物含量进行了Pearson相关回归系数分析(表3)。结果表明, 丰水期浮游植物丰度与水温呈负相关(<0.05), 与pH和悬浮物呈正相关(<0.05); 枯水期浮游植物丰度与pH(<0.01)、悬浮物(<0.05)呈正相关, 而与盐度(<0.05)呈负相关。
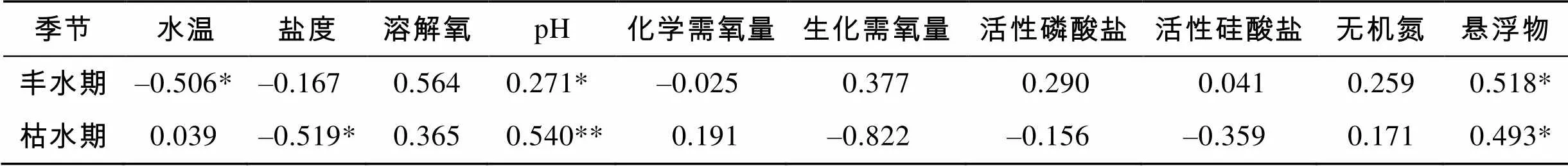
表3 粤西近岸海域浮游植物丰度与环境因子的相关性
注: * .<0.05; **.<0.01
3 讨论
3.1 粤西海域浮游植物的分布及其与环境因子的关系
由于浮游植物无自主活动能力, 基本上随波逐流, 因此其丰度变动主要受到温度、盐度、pH、沿岸流、风生流、冲淡水、海水营养盐、叶绿素等的影响[20-23], 其中, 本次调查粤西海域浮游植物丰度变化与pH和悬浮物呈正相关, 丰水期与水温呈负相关, 枯水期与盐度呈负相关。粤西海域浮游植物丰度表现为近岸站位高于离岸站位, 丰水期浮游植物丰度有湛江沿岸站位高于其他调查站位, 枯水期浮游植物丰度则由西南站位向东北站位递减。夏季粤西近岸各海域浮游植物受光强、温度和摄食压力的影响差别不大, 浮游植物丰度主要取决于外源性营养盐的输入[24-26], 大陆径流和入海河流携带大量营养物质流入海中并不断向外海扩散, 近岸浮游植物因营养盐充足大量繁殖形成密集区, 随着营养物质不断向外海稀释、减少, 离岸浮游植物也不断减少, 最终出现近岸站位丰度高于离岸站位的现象。湛江市的经济活动中心主要分布在湛江港湾沿岸, 丰水期降雨量大, 各种工农业废水和生活污水的大量排入以及港湾内大面积的水产养殖给该海域带来丰富的营养物质, 为浮游植物的增殖提供充足的营养盐, 从而形成浮游植物高值区。此结果与龚玉艳等[24]在2010年的调查结果一致。枯水期粤西海域浮游植物丰度呈现由西南部向西北部逐渐减少的趋势, 主要是球形棕囊藻在南面站位大量爆发与繁殖造成的结果。吴迪生等[27]的研究发现持续温暖天气、降水少、风速小、相对湿度偏高的气候条件有利于赤潮藻类的繁殖与生长, 且赤潮期间极少的降水是维持赤潮的一个重要因素。冬季的低温是制约浮游植物大量繁殖的一个重要因素, 粤西不同纬度地区的温度会有一定差别, 湛江港冬季的水温相对较高(15~21℃), 这一水温与广东东部海域1997年暴发球形棕壤藻赤潮时海域表层水温(18~24℃)相近, 随着调查海域纬度升高、水温下降, 不适宜球形棕壤藻生长, 印证了郭瑾等[28]的研究理论: 温度是球形棕壤藻生长的显著因子, 在30℃达到最大生长速率, 在20~25℃达到更高藻丰度。同时水产养殖业的大力发展, 繁忙的海上交通和陆地工业经济的发展给湛江港湾水域带来不少污染物, 为浮游植物的生长繁殖提供了营养物质, 相比较于北面调查站位更利于藻类的大量繁殖与生长, 造成枯水期调查海域南部湛江港湾调查站位浮游植物丰度明显高于北部茂名阳江调查站位。
3.2 浮游植物群落结构特征及其与历史资料的比较
3.2.1 浮游植物群落结构特征
粤西近岸浮游植物群落结构的Shannon-wiener指数、Margalef指数和Pielou均匀度指数表现出明显的季节变化和区域特点。总体呈现丰水期浮游植物多样性水平高于枯水期。稳定而适宜的温度和盐度条件是藻类生长繁殖的基础, 资料显示, 在世界各地的海洋中, 棕囊藻赤潮一般于春季有规律地暴发[29], 受粤西热带气候影响, 粤西海域冬季水温偏高, 本次调查平均水温可达13℃, 且枯水期降雨量的减少和养殖海域无机氮(特别是NO3-N)的增加给粤西海域棕囊藻赤潮的暴发创造了有利条件[30]。调查发现赤潮期间伴随藻类主要为海链藻()和辐杆藻等大型、成链的藻类, 显示了球形棕囊藻和大型硅藻占据相同的生态位, 且在竞争中占明显优势。赤潮藻类的暴发抑制了其他藻类的生长, 造成生态环境的不平衡(S1-S6和S7-S12的Pielou均匀度指数范围和平均值分别为0.005~0.325、0.234~0.788; 0.129、0.615), 导致粤西海域枯水期群落结构整体水平降低。粤西近岸浮游植物群落具有明显的区域性分布特征, 在丰水期和枯水期, 分别形成了4个和3个不同浮游植物生态区, 生境复杂, 这一调查结果与龚玉艳等[24]在2010年的调查结果一致。
3.2.2 粤西近岸海域浮游植物群落与历史资料的比较
粤西近岸海域浮游植物的历史资料对比表明(表4), 相对于2000年之前的调查结果, 本次调查的浮游植物物种种数降低、浮游植物丰度显著升高, 这一结果与龚玉艳等[24]在2010年的调查结果一致。与1998~1999年的调查结果比较, 本次调查粤西海域秋季和冬季的浮游植物物种组成分别由126种和139种下降到84和72种, 而丰度由0.15×104和0.42×104个/L分别上升到7.14×104和315.62×104个/L。比较于2010年雷州半岛东部海岸带的调查结果, 本次调查物种组成和细胞丰度差异不显著, 物种组成分别由71种和76种过度为84种和72种, 丰度由370.08×104和23.45×104个L演变为7.14×104和315.62×104个/L。
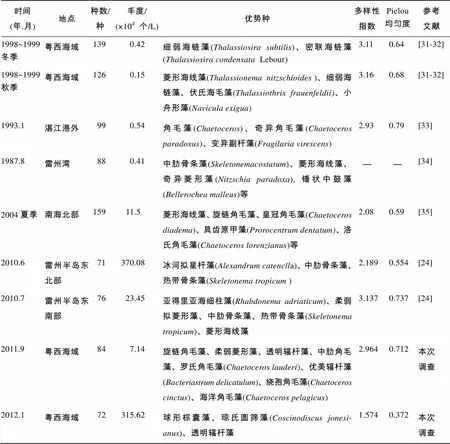
表4 研究海域浮游植物历史资料对比
与历史资料相比较, 粤西海域优势种虽有一定演替, 但基本为该海域常见种类, 由近岸、近海低盐性种类和外海广布性种类构成。粤西海域1998~1999年航次调查以外海广布性种类的海链藻属为优势种; 雷州半岛东北部附近海域1987年和2010年的调查结果显示该海域的优势种主要为暖温带近岸种类的骨条藻属()和热带近岸性种类的海线藻属(Thalassionema); 而1993年湛江港外的优势种调查结果与本次调查结果较为相近, 均主要以近岸、近海低盐性的角毛藻属()作为主要优势种。
粤西海域浮游植物多样性历史数据对比表明, 本次调查丰水期多样性水平变化不大; 枯水期多样性水平呈下降状态, 群落结构稳定性下降。
4 结论
粤西海域浮游植物种类组成以硅藻为主, 甲藻次之, 蓝藻和金藻较少。不同季节浮游植物种类组成及其群落结构变化较大, 存在明显的季节性差异。
丰水期浮游植物丰度平均为7.14×104个/L, 枯水期浮游植物丰度平均为315.62×104个/L, 枯水期浮游植物丰度总体比丰水期高; 浮游植物丰度大体呈现近岸站位高于离岸站位, 而丰水期浮游植物丰度有湛江沿岸站位高于其他调查站位, 枯水期浮游植物丰度则由西南部向东北部递减。
粤西海域浮游植物优势种表现出明显季节差异和平面分布差异。丰水期优势种有4种, 枯水期优势种仅1种, 以金藻门的球形棕囊藻占绝对优势。
粤西海域浮游植物物种多样性水平相对较高, 不同季节不同区域物种多样性水平差异明显。群落结构多样性为枯水期西南部海域明显低于东北部海域, 丰水期站位间多样性无明显变化规律, 相比之下, 丰水期群落物种多样性水平相对较高。
丰水期浮游植物丰度与水温呈负相关(<0.05), 与pH和悬浮物呈正相关(<0.05); 枯水期浮游植物丰度与pH(<0.01)、悬浮物(<0.05)呈正相关, 而与盐度(<0.05)呈负相关。
[1] 张婷, 李林, 宋立荣. 熊河水库浮游植物群落结构的周年变化[J]. 生态学报, 2009, 29(6): 2971-2979. Zhang Ting, Li Lin, Song Lirong. Annual dynamics of phytoplankton abundance and community structure in the Xionghe Reservoir[J]. Acta Ecologica Sinica, 2009, 29(6): 2971-2979.
[2] 王晓伟, 李纯厚, 戴明. 大亚湾湾口海域冬季浮游植物生长限制性元素研究[J]. 南方水产, 2007, 13(4): 26-31. Wang Xiaowei, Li Chunhou, Dai Ming. Studies on limited nutrient factors in outer of Daya Bay in winter[J]. South China Fisheries Science, 2007, 13(4): 26-31.
[3] 袁华荣, 陈丕茂, 贾晓平, 等. 北部湾东北部游泳生物资源现状[J]. 南方水产科学, 2011, 7(3): 31-38. Yuan Huarong, Chen Pimao, Jia Xiaoping, et al. Nekton resources in northeast Beibu Gulf[J]. South China Fisheries Science, 2011, 7(3): 31-38.
[4] 吴玉霖, 孙松, 张永山, 等. 胶州湾浮游植物数量长期动态变化的研究[J]. 海洋与湖沼, 2004, 35(6): 518-523. Wu Yulin, Sun Song, Zhang Yongshan, et al. Quantitative study on long term variation of phytoplankton in JiaoZhou Bay[J]. Oceanologia et Limnologia Sinica, 2004, 35(6): 518-523.
[5] 吴玉霖, 孙松, 张永山. 环境长期变化对胶州湾浮游植物群落结构的影响[J]. 海洋与湖沼, 2005, 36 (6): 487-498. Wu Yulin, Sun song, Zhang Yongshan. Long-term change of environment it’s influence on phytoplankton community structure in JiaoZhou Bay[J]. Oceanologia et Limnologia Sinica, 2005, 36 (6): 487-498.
[6] 廖秀丽, 陈丕茂, 马胜伟, 等. 大亚湾杨梅坑海域投礁前后浮游植物群落结构及其与环境因子的关系[J]. 南方水产科学, 2013, 9(5): 109-119. Liao Xiuli, Chen Pimao, Ma Shengwei, et al. Community structure of phytoplankton and its relationship with environmental factors before and after construction of artificial reefs in Yangmeikeng, Daya Bay[J]. South China Fisheries Science, 2013, 9(5): 109-119.
[7] 孙军, 刘东艳, 杨世民, 等. 渤海中部和渤海海峡及邻近海域浮游植物群落结构的初步研究[J]. 海洋与湖沼, 2002, 33(5) : 461-471. Sun Jun, Liu Dongyan, Yang Shimin, et al. The preliminary study on phytoplankton community structure in the central Bohai Sea and the Bohai strait and its adjacent area[J]. Oceanologia et Limnologia Sinica, 2002, 33(5) : 461-471.
[8] 广东省海岸带和海涂资源综合调查大队, 广东省海岸带和海涂资源综合调查领导小组办公室. 广东省海岸带和海涂资源综合调查报告[M]. 北京: 海洋出版社, 1987: 205-228. Group and Leading Team Office of Comprechensive Resource Inverstigation on the Littoral Zone and Tidal Flat of Guangdong Provice. The report of comprehensive resource investigation on the littoral zone and tidal flat[M]. Beijing: China Ocean Press, 1987: 205-208.
[9] 国家海洋局. 南海中部海域环境资源综合调查报告[M]. 北京: 科学出版社, 1988: 162-207. State Oceanic Administratration of China. The report of environmental resource investigation in the central part of the South Sea[M]. Beijing: Science Press, 1988: 162-207.
[10] 农牧渔业部水产局, 农牧渔业部南海区渔业指挥部. 南海区渔业资源调查和区划[M]. 广州: 广东科技出版社, 1989: 37-81. Fishery Bureau, Fishery Administration Office of South China Sea, Ministry of Agriculture, Herd and Fishery. Fishery resource investigation and section of South China Sea[M]. Guangzhou: Guangdong Science and Technology Press, 1989: 37-81.
[11] 国家技术监督局. 海洋调查规范(GB12763.6-91)[S]. 北京: 中国国家标准出版社, 1992. Technical Supervise Bureau of China. Standard methods for marine investigation[S]. Beijing: China Standard Press, 1992.
[12] Jaccard P. Nouvelles recheres surla distribution floral[J]. Bull Soc Vaud Sa Nat, 1998, 44: 223-270.
[13] 王备新, 杨莲芳, 胡本进, 等. 应用底栖动物完整性指数B-IBI评价溪流健康[J]. 生态学报, 2005, 25(6): 1481-1490. Wang Beixin, Yang Lianfang, Hu Benjin, et al. A preliminary study on the assessment of stream ecosystem health in south of Anhui Province using Benthic-Index of Biotic Integrity[J]. Acta Ecologica Sinica, 2005, 25(6): 1481-1490.
[14] Shannon C E, Weaver W. The mat hematical theory of communication[M]. Urbana 1L: University f Illinois Press, 1949: 1-125.
[15] 马克平, 刘玉明. 生物群落多样性的测度方法I a多样性的测度方法(下)[J]. 生物多样性, 1994, 2(4): 231-239. Ma Keping, Liu Yuming. Measurement of biotic community diversity. I. α diversity (2)[J].Biodiversity Science, 1994, 2(4): 231-239.
[16] Pielou E C. An introduction to mathematical ecology[M]. New York: Wiley Interscience: 1969: 1-286.
[17] 戴明, 李纯厚, 张汉华, 等. 海南岛以南海域浮游植物群落特征研究[J]. 生物多样性, 2007, 15(1): 23-30. Dai Ming, Li Chunhou, Zhang Hanhua, et al. Characteristics of the phytoplankton community in the southern waters of Hainan Island[J]. Biodiversity Science, 2007, 15(1): 23-30.
[18] Bray J R, Curtis J T. An ordination of the upland forest communities of southern Wisconsin[J]. Ecological Monographs, 1957, 27: 325-349.
[19] Rohlf F J. NTSYS-Pc: Numerical taxonomy and multivariate analysis system, Version 2.1.Users Guide[M]. New York: Exeter publishing Setauket, 2000.
[20] Perry M J, Eppley R W. Phosphate uptake by phytoplankton in the central North Pacific Ocean[J]. Deep-Sea Res, 1981, 28: 39-49.
[21] Carpenter E J, Capone D G. Nitrogen in marine environment[M]. New York: Academic Press, 1983: 233-274.
[22] 殷旭旺, 渠晓东, 李庆南, 等. 基于着生藻类的太子河流域水生态系统健康评价[J]. 生态学报, 2012, 32(6): 1677-1691. Yin Xuwang, Qu Xiaodong, Li Qingnan, et al. Using periphyton assemblages to assess stream conditions of Taizi River Basin, China[J]. Acta Ecologica Sinica, 2012, 32(6): 1677-1691.
[23] 章守宇, 邵君波, 戴小杰. 杭州湾富营养化及浮游植物多样性问题的探讨[J]. 水产学报, 2001, 25(6): 512-517. Zhang Shouyu, Shao Junbo, Dai Xiaojie. Studies on eutrophication and phytoplankton diversity in Hangzhou Bay[J]. Journal of Fisheries of China, 2001, 25(6): 512-517.
[24] 龚玉艳, 张才学, 孙省利, 等. 2010年夏季雷州半岛海岸带浮游植物群落结构特征及其与主要环境因子的关系[J]. 生态学报, 2012, 32(19): 5972-5985. Gong Yuyan, Zhang Caixue, Sun Xingli, et al. Community characteristics of phytoplankton in the coastal area of Leizhou Peninsula and their relationships with primary environmental factors in the summer of 2010[J]. Acta Ecologica Sinica, 2012, 32(19): 5972-5985.
[25] 赵明辉, 黄洪辉, 齐占会, 等. 南海北部湾浮游动物的景观格局分析[J]. 南方水产, 2010, 6(6): 41-45. Zhao Minghui, Huang Honghui, Qi Zhanhui, et al. Landscape pattern analysis ofpPlankton in Northern South China Sea[J]. South China Fisheries Science, 2010, 6(6): 41-45.
[26] 孙晓庆, 董树刚, 汤志宏. 营养盐和光照对浮游植物群落结构的影响[J]. 南方水产, 2008, 14(1): 1-9. Sun Xiaoqing, Dong Shugang, Tang Zhihong. Influences of nutrients and illuminance on phytoplankton community structure[J]. South China Fisheries Science, 2008, 14(1): 1-9.
[27] 吴迪生, 黎广媚, 赵雪, 等. 广东沿海赤潮与海洋水文气象关系研究[J]. 海洋通报, 2005, 24(5): 16-22. Wu Disheng, Li Guangmei, Zhao Xue, et al. The research on the relationship between red tide in the coastal Guangdong and marine hydrometeorolgy[J]. Marine Science Bulletin, 2005, 24(5): 16-22.
[28] 郭瑾, 杨维东, 刘洁生, 等. 温度、盐度和光照对球形棕壤藻生长和产毒的影响研究[J]. 环境科学, 2007, 27(8): 1341-1346. Guo Jin, Yang Weidong, Liu Jiesheng, et al. Effects of salinity、temperature and light intensity on the growth and toxin production of[J]. Acta Scientiae Circumstantiae, 2007, 27(8): 1341-1346.
[29] Lancelot C, Keller M D. Autecology of the marine haptophytesp[C]//Physiobgical ecology of hamful algal blooms. Berlin, Germany: Springer, 1998: 209-224.
[30] 徐宁, 齐雨藻, 陈菊芳, 等. 球形棕壤藻赤潮成因分析[J]. 环境科学学报, 2003, 23(1): 113-118. Xu Ning, Qi Yuzao, Chen Jufang, et al. Analysis on the cause ofred tide[J]. Acta Scientiae Circumstantiae, 2003, 23(1): 113-118.
[31] 蔡文贵, 李纯厚, 贾晓平, 等. 粤西海域浮游植物种类的动态变化及多样性[J]. 海洋环境科学, 2003, 22(4): 34-37. Cai Wengui, Li Chunhou, Jia Xiaoping, et al. Dynamic variation and the biodiversity for the species of phytoplankton in West Guangdong Waters[J]. Marine Environmental Science, 2003, 22(4): 34-37.
[32] 蔡文贵, 贾晓平, 李纯厚. 粤西海域浮游植物的时空分布及其漂移特点[J]. 热带海洋学报, 2005, 24(3): 65-72. Cai Weigui, Jia Xiaoping, Li Chunhou, et al. Spatial temporal distribution and excursion characteristic of phytoplankton in Western Guangdong waters[j]. Jouranal of Tropical Oceanography, 2005, 24(3): 65-72.
[33] 刘玉, 吴群河, 陈新庚. 湛江海域浮游藻类生态及其多样性[J]. 生态科学, 1995, 1: 21-25. Liu Yu, Wu Qunhe, Chen Xingeng. Phytoplankton ecology and its diversity in Zhanjiang seawaters[J]. Ecologic Science, 1995, 1: 21-25.
[34] 吕颂辉, 齐雨藻, 钱宏林, 等. 南海港湾(雷州湾)浮游植物与赤潮生物研究[J]. 暨南大学学报: 自然科学与医学版, 1994, 15 (1) : 146-150. LYv Songhui, Qi Yuzao, Qian Honglin, et al. Studies on phytoplankton and red tide organisms in embayments on West Guangdong coast Ⅳ. Leizhou bay[J]. Journal of Jinan University(Natural Science), 1994, 15(1): 146-150.
[35] 乐凤凤, 孙军, 宁修仁, 等. 2004年夏季中国南海北部的浮游植物[J]. 海洋与湖沼, 2006, 37(3): 238-248. Le Fengfeng, Sun Jun, Ning Xiuren, et al. Phytoplankton in the Northern south China sea in summer 2004[J]. Oceanologia Et Limnologia Sinica, 2006, 37(3): 238-248.
Ecological characteristics of the phytoplankton community structure in the West Guangdong coastal area
LIN Hua-jian, XU Shan-nan, LIU Yong, GONG Yu-yan, LI Chun-hou
(Key Laboratory of South China Sea Fishery Resources Exploitation & Utilization, Ministry of Agriculture, Key Laboratory of Fishery Ecology and Environment, Guangdong Province, South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China)Received:Nov.12, 2014
West Guangdong coastal area; phytoplankton; abundance; community structure
Based on data collected during an ecological investigation of West Guangdong before 30 meters coastal waters during a wet (September 2011) and a dry season (January 2012), the species composition, dominant species, abundance, and community diversity of phytoplankton were analyzed. A total of 116 species consisting of 41 genera belonging to 4 phyla were identified. Diatom was the predominant phyla group. Forty common species occurred in both wet and dry seasons, with a species similarity index between the two seasons of 34.48% suggesting significant seasonal variation in the species composition. Only one dominant species occurred during the dry season,, with a dominance of 0.496. During the wet season, there were four dominant species, the main one waswith a dominance of 0.249. Phytoplankton abundance showed significant seasonal and regional differences and was higher in inshore waters than in offshore. During the wet season, the phytoplankton abundance in the Zhanjiang coastal waters was the highest. During the dry season, phytoplankton abundance decreased from the southwest to northeast coastal waters. In general, the phytoplankton abundance during the dry season was higher than during the wet season. Community diversity indexes in the southwest waters during dry season were obviously lower than those in the northeast waters. Diversity indexes during the wet season were relatively higher than during the dry season, but there were no significant differences in diversity during the wet season. Correlation analysis demonstrated that the phytoplankton abundance was positively correlated to pH and suspended solids (SS), and negatively correlated to water temperature during the wet season and salinity during the dry season.
S949
A
1000-3096(2016)07-0073-10
10.11759/hykx20141112002
2014-11-12;
2014-12-09
国家自然科学基金项目(31100362); 公益性行业(农业)科研专项经费(201403008); 国家重点基础研究发展计划项目(2015CB452904); “十二五”国家科技支撑计划项目(2012BAD18B01); 广州市科技计划项目(2014J2200020)
[Foundation: National Natural Science Foundation of China, No. 31100362; Special Fund for Agro-scientific Research in the Public Interest, No.201403008; National Key Basic Research Program of China, No.2015CB452904; Key Projects in the National Science & Technology Pillar Program during the Twelfth Five-year Plan Period, No. 2012BAD18B01; Science and Technology Program of Guangzhou, China, No. 2014J2200020]
林华剑(1988-), 男, 福建泉州人, 硕士研究生, 从事海洋生态环境保护研究, 电话: 15602389557, E-mail: lhj0914@126.com;李纯厚,通信作者, E-mail: scslch@vip.163.com
(本文编辑: 谭雪静)
