基于3种基因序列对百合属Leucolirion组的系统发育与种间关系研究
2016-10-14赖山潘沈呈娟周颂东何兴金
赖山潘,沈呈娟,周颂东,何兴金
(四川大学 生命科学学院,生物资源与生态环境教育部重点实验室,成都 610064)
基于3种基因序列对百合属Leucolirion组的系统发育与种间关系研究
赖山潘,沈呈娟,周颂东,何兴金*
(四川大学 生命科学学院,生物资源与生态环境教育部重点实验室,成都 610064)
中国拥有丰富的野生百合种质资源,但有关百合属的系统发育与进化关系研究相对较少。该研究基于ITS、cpDNA(trnV-ndhC、petL-psbE)及单拷贝核基因AKT等3个基因片段,选取特有分布于西南地区的百合属6个近缘类群共48份样本及GenBank获得的数据,对百合属Leucolirion组进行了系统发育与种间关系研究。结果表明:(1)ITS结果支持前人的处理,即将Leucolirion组细分为2个亚组,subsect.Leucolirion6a和subsect.Leucolirion6b。建议将湖北百合(L.henryi)和南川百合(L.rosthornii)归入到subsect.Leucolirion6a,与泸定百合(L.sargentiae)、淡黄花百合(L.sulphureum)、宜昌百合(L.leucanthum)和岷江百合(L.regale)等4个种一起组成Leucolirion组;原本属于Archelirion组的野百合(L.brownii)与subsect.Leucolirion6b聚为一支,故将它与台湾百合(L.longiflorm)、麝香百合(L.formosanum)等组成一个新组,命名为百合组(sect.Lilium)。(2)组间关系上,Leucolirion组与百合组的系统发育关系较远,前者与分布于日本的Archelirion组形成姐妹类群,而后者与Sinomartagon组的亲缘关系更近。(3)基于叶绿体联合基因和单拷贝核基因AKT的6个近缘种的遗传距离显示,湖北百合与南川百合明显区别于Leucolirion组内拥有白色管状花的4个类群。叶绿体联合基因树和单拷贝核基因树结果表明,泸定百合、淡黄花百合、岷江百合和宜昌百合在遗传水平上表现出了极高的相似性,遗传关系十分相近,这可能与百合属植物特殊的繁育系统有关,影响了物种间遗传变异与分化。
Leucolirion;系统发育;ITS;叶绿体基因;单拷贝核基因;种间关系
百合属(LiliumL.)隶属于百合科(Liliaceae Juss.),全世界约110~115余种,广泛分布于北半球温带地区,中国有55种[1],是百合属植物的起源和分布中心[2],其中以西南地区分布最为丰富。Comber[3]依据13个形态性状和2个生殖特性将百合属划分为7个组9个亚组。其中,Leucolirion组的主要特征为叶散生,无柄,花为喇叭状,子叶出土型发芽。该组组下依据鳞茎颜色又分成了2个亚组,分别为subsect.Leucolirion6a和subsect.Leucolirion6b。前者包括了岷江百合(L.regale)、宜昌百合(L.leucanthum)、泸定百合(L.sargentiae)和淡黄花百合(L.sulphureum),鳞茎为紫色,特有分布于中国西南地区;后者由麝香百合(L.longiflorum)、台湾百合(L.formosanum)和菲律宾百合(L.philippinense)等组成,鳞茎为白色,主要分布于中国东部沿海及台湾地区。但随着细胞学与分子系统学研究的深入,该组的划分及组下类群得到了进一步的修订与发展。
最早,Nishikawa等[4-5]及Hayashi 等[6]对百合属的分子系统学研究表明,Leucolirion组不是一个单系,该组下存在2个亲缘关系较远的分支。Gao等[7]对百合属的核型研究显示,形态上同为白色管状花的类群,依据居间随体的有无将Leucolirion组分成2个亚组,产中国西南地区的岷江百合、宜昌百合、泸定百合和淡黄花百合具有居间随体被分为了一个亚组;而文山百合(L.wenshanense)、安徽百合、麝香百合和台湾百合则没有这一结构被归于另一亚组,这一处理也支持了Comber[3]基于鳞茎颜色对该组的划分,但相较于Comber在1949年对Leucolirion组的划分,加入了许多后来发表的新类群。近年来,更大尺度下的系统发育重建使得百合属的分类系统越发清晰。Lee等[8]对百合属83种14变种进行ITS建树分析,Leucolirion组被分成2个单独的分支,Group I和Group VI。Gao等[2,9]基于ITS和matK对百合属的系统发育研究结果显示,湖北百合(L.henryi)与subsect.Leucolirion6a聚为一支,丰富了这一亚组的物种组成,且该亚组与分布于日本的Archelirion组形成了姐妹类群关系;subsect.Leucolirion6b则包括了野百合(L.brownii)、安徽百合(L.anhuiense)、文山百合、麝香百合、台湾百合和菲律宾百合等6个种,单独聚为一支,与subsect.Leucolirion6a的亲缘关系较远。Du等[10]构建了目前最大的ITS进化树,结果支持把湖北百合和南川百合(L.rosthornii)归入到subsect.Leucolirion6a中,Leucolirion组被修正为2个分支。以上的研究结果表明,Leucolirion组是不自然的,根据其地理分布式样以及细胞学、分子系统学证据,该组需要被细分和整理。
目前对湖北百合和南川百合的系统位置还没有一个定论,最早Comber[3]依据形态特征将二者归入Sinomartagon组,但分子系统学的研究与形态分类结果出现了巨大的差异,它们的系统位置需要重新评估。而岷江百合、宜昌百合、泸定百合和淡黄花百合的形态十分相近,都为白色管状花,但种间界限并不明显,分子系统学的结果显示系统发育树的节点支持率并不高,分支内存在大量的平行支[2,10],因此种间关系需要进一步研究和探讨。本实验在前人研究的基础上,通过大量的野外调查和取样,选取上述特产于中国西南地区的形态相似、分子系统上近缘的6个类群(图1),基于叶绿体基因片段ndhC-trnV和petL-psbE、核糖体基因ITS片段及单拷贝核基因AKT片段,对其系统发育与种间关系进行了分析研究。本研究类群的许多成员花大美丽,且地下鳞茎可作药用,广泛运用于遗传育种,花卉培育等领域,因此开展这些种的系统发育关系的研究有利于为其合理的开发与利用提供理论基础。
1 材料和方法
1.1 材料取样
本研究从2013~2014年开始展开野外调查以及样品收集工作。湖北百合、南川百合主要分布于重庆、贵州和湖北一带,共收集到10个居群。岷江百合狭域分布于岷江流域,在汶川、茂县一带收集到4个居群。宜昌百合及其变种紫脊百合L.leucanthumvar.centifolium共5个居群,泸定百合主要分布于四川、重庆,共计11个居群;而淡黄花百合分布最为广泛,云南、贵州、广西等地均产,共收集到18个居群。本研究利用所获得的基因序列以及从GenBank获得的其他百合属植物的基因序列做对比,对Leucolirion组的系统发育与种间关系进行了研究。通常情况下,每个居群取1~2个个体作为代表。材料采集信息、凭证标本见表1。新鲜叶片均用硅胶快速干燥保存,凭证标本存放在四川大学植物标本馆(SZ),每份标本的地理采集信息都用佳明GPS (GPSMAP 62sc, Shanghai)设备记录点位。
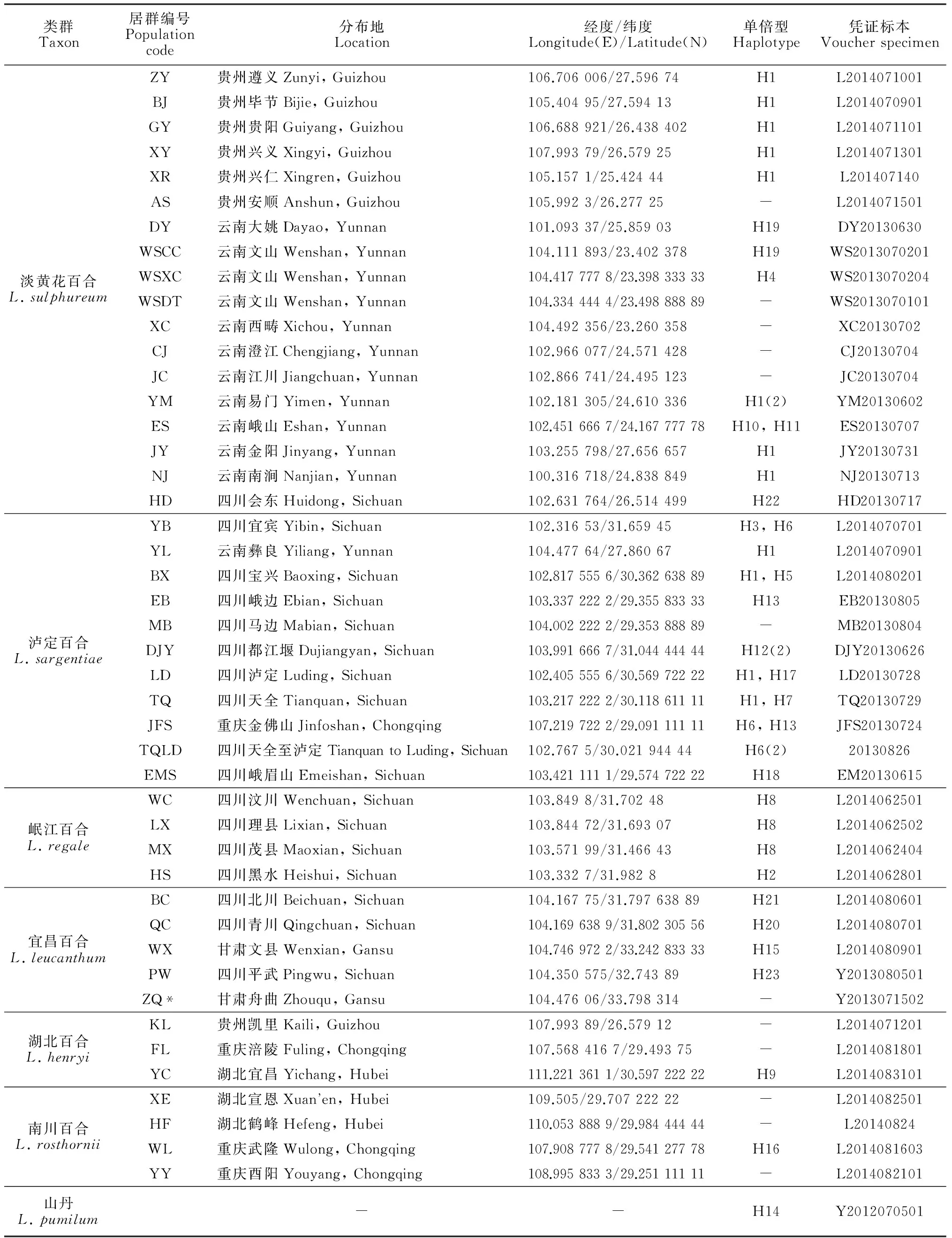
表1 材料来源
注:ZQ*居群为宜昌百合的变种,紫脊百合
Note: ZQ* population indicate a variety ofL.leucanthum,L.leucanthumvar.centifolium
1.2 方 法
1.2.1 DNA提取和PCR扩增 本研究使用天根生化科技有限公司(Tiangen Biotech CO. LTD., 北京)生产的植物基因组DNA 提取试剂盒提取本研究的所有样品,试剂盒法本质上是基于CTAB法[11],实验流程按试剂盒说明书进行。提取得到的DNA产物溶于TE缓冲液,保存在-20 ℃冰箱中,备用。
核糖体DNA(nrDNA)的内转录间隔区(Internal transcribed spacers, ITS)是分子系统学研究运用最为广泛的基因。本实验对分子序列ITS进行整段扩增(包括ITS1、ITS2以及5.8S),依据White等[12]开发的通用引物,具体的扩增程序如下:94 ℃预变性2 min;中间环节为94 ℃变性45 s, 45 ℃退火45 s, 72 ℃延伸1 min,35个循环;最后72 ℃延伸10 min。叶绿体基因片段trnV-ndhC、petL-psbE的详细信息及扩增程序参考Shaw等[13]。AKT单拷贝基因引物参考Douglas等[14],PCR扩增体系与程序稍作调整如下:包括2.5 μL 10×PCR buffer,200 μmol/L dNTPs,2 μL DNA模版,2.5 mmol/L MgCl2,4 μmol/L 引物对和0.5 UTaqDNA酶;95 ℃预变性4 min,中间35个循环分别为95 ℃变性30 s,58 ℃延伸30 s,72 ℃延伸2.5 min,最后72 ℃延伸4 min。PCR 产物的纯化与测序工作由北京华大基因科技有限公司完成,测序所用引物与PCR 引物相同,正反双向测序,以保证测序结果的准确性。
1.2.2 数据分析 测序得到的双向序列用Lasergene文件包中的SeqMan[15]进行拼接合成,然后用BioEdit软件导入数据成fas格式文件,在Mega 4[16]打开,对序列进行自动比对和必要的手工调整,调整原则参照Morrison[17]。空格插入作为缺失数据处理,并在Mega 4中分析序列的变异位点(variable sites)、保守位点(conserved sites)、简约信息位点(parsimony information sites)和GC含量。采用1 000代运算构建临近树(neighbor-joining, NJ)对研究类群进行聚类分析,kimura 2-parameter(K2P)模型计算种间遗传距离(genetic distances)。
利用PAUP ver. 4.0b10[18]对研究类群构建最大简约树(Maximum Parsimony, MP),所有的特征设置为无序和不加权等,采用启发式搜索(heuristic search)进行1 000次随机添加(random addition)。以TBR(tree-bisection-reconnection)进行枝长交换获取分支树,计算100万代,通过自展法(bootstrap, BS)检测各分支的支持率。贝叶斯树(Bayesian inferences, BI)在软件MrBayes version 3.1.2[19]上完成,首先在MrModeltest version 2.2[20]上搜索最合适的模型,确定最适替代模型为SYM+G,以随机树开始,4条马尔科夫-蒙塔卡罗(Markov Chain Monte Clarlo, MCMC)同时运行10 000 000,每1 000代保存1棵树,设置温度为0.2 ℃,前30%的树被燃烧丢弃(burn-in),剩下的树合并后用于后序建树分析。
2 结果与分析
2.1 基因序列特征
核基因ITS的数据矩阵包含了98条序列,包括Leucolirion组和Archelirion组主要成员以及产于中国的其他百合属植物,比对后的ITS基因序列平均长度为621(615~639)bp,包含了234个变异位点,信息位点135个,占21.74%,G+C含量62.1%。叶绿体基因片段petL-psbE和ndhC-trnV对6个近缘种的18个个体进行扩增测序,长度分别为1 192~1 196 bp和746~753 bp,信息位点分别为11和10个。由于基因序列的信息位点有限,对petL-psbE和ndhC-trnV进行了非一致性长度检验(ILD检验),结果显示两条序列能够拼接合并(P>0.5),总长为1 939~1 946 bp,含有23个变异位点,信息位点21个(1.1%)。单拷贝核基因AKT对研究类群的47个个体进行扩增,序列平均长度为1 459 bp,G+C含量达37.9%,变异位点100个,简约信息位点67个(4.6%)。
2.2 遗传距离分析
以cpDNA联合基因和单拷贝核基因AKT的序列为数据基础,利用Mega软件中的K2P模型对本研究类群的6个近缘种3个居群的18个个体进行遗传距离分析(表2)。基于cpDNA联合基因的遗传距离矩阵结果显示,6个近缘种种间的遗传距离很小,平均约为0.004 4。泸定百合、淡黄花百合、岷江百合和宜昌百合的种间遗传距离相似,为0.000~0.002。湖北百合、南川百合与4个白色管状花类群的遗传距离在0.006~0.008之间。单拷贝核基因AKT在种间的遗传距离平均为0.019 9,与叶绿体基因的结果类似,前4个种种间遗传距离很小,大小在0.001~0.007之间。湖北百合与南川百合的种间遗传距离平均为0.006,与泸定百合等4个种的遗传距离在0.032~0.036之间。
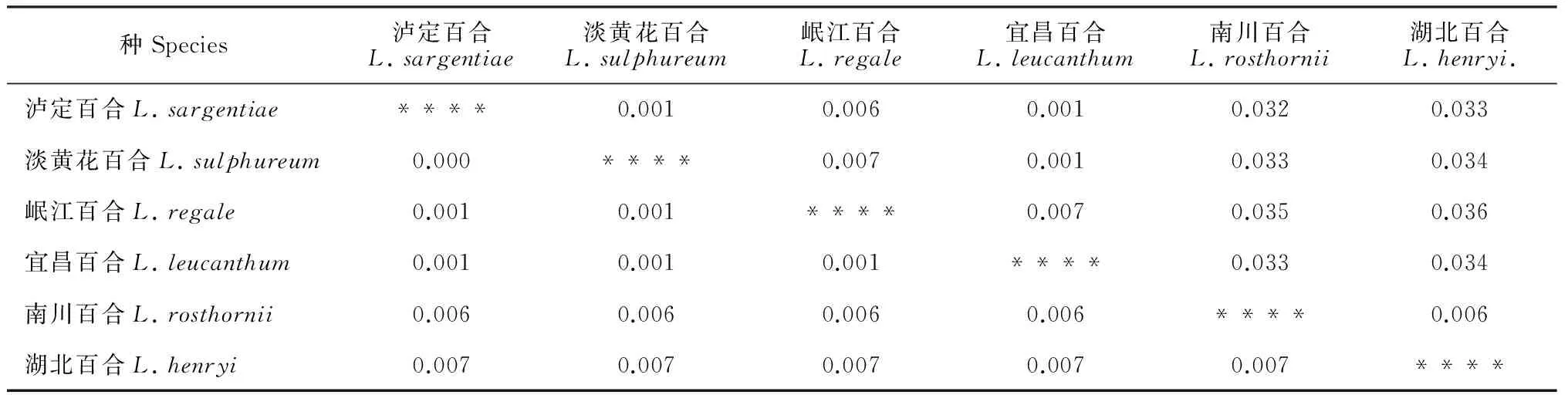
表2 6个近缘种的种间遗传距离
注:对角线上方为基于单拷贝基因AKT的遗传距离,对角线下方为cpDNA联合片段的遗传距离
Note: Genetic distances based on nuclear single-copy AKT are above diagonal, and genetic distances based on cpDNA combination are below diagonal
2.3 系统发育关系分析
基于ITS序列构建的MP树和贝叶斯树的结果显示,两种方法构建的系统发育树拓补结构相似。如图2所示,Leucolirion组被分为了2个分支,分别为subsect.Leucolirion6a和subsect.Leucolirion6b,且这2个分支的系统发育关系很远。subsect.Leucolirion6a包含了泸定百合、淡黄花百合、岷江百合和宜昌百合等4个种,湖北百合和南川百合与该亚组很好地聚为一支,该分支与Archelirion组形成了姐妹类群关系(PP/BS, 0.85/80)。subsect.Leucolirion6b包括了台湾百合、麝香百合,野百合与该亚组的聚类结果得到很高的支持率(PP/BS, 0.95/88),整个分支与Sinomartagon组的系统发育关系更为接近。
在种间关系上,本研究中的6个近缘种形成了一个很好的独立分支,支持前人的分子系统研究结果。湖北百合与南川百合先分化出来,但二者之间的种间关系不明确。泸定百合、淡黄花百合、岷江百合和宜昌百合等4个种的多个居群数据并没有得到很好的聚类结果,内部表现出近似平行的分支,种间界限模糊,在分子水平上表现出了极高的相似性,但这4个类群在分子系统树上却稳定地聚为一个高支持率的单系分支,表明这4个类群拥有一个最近的共同祖先,它们具有非常紧密的亲缘关系,也支持了它们的单系性。
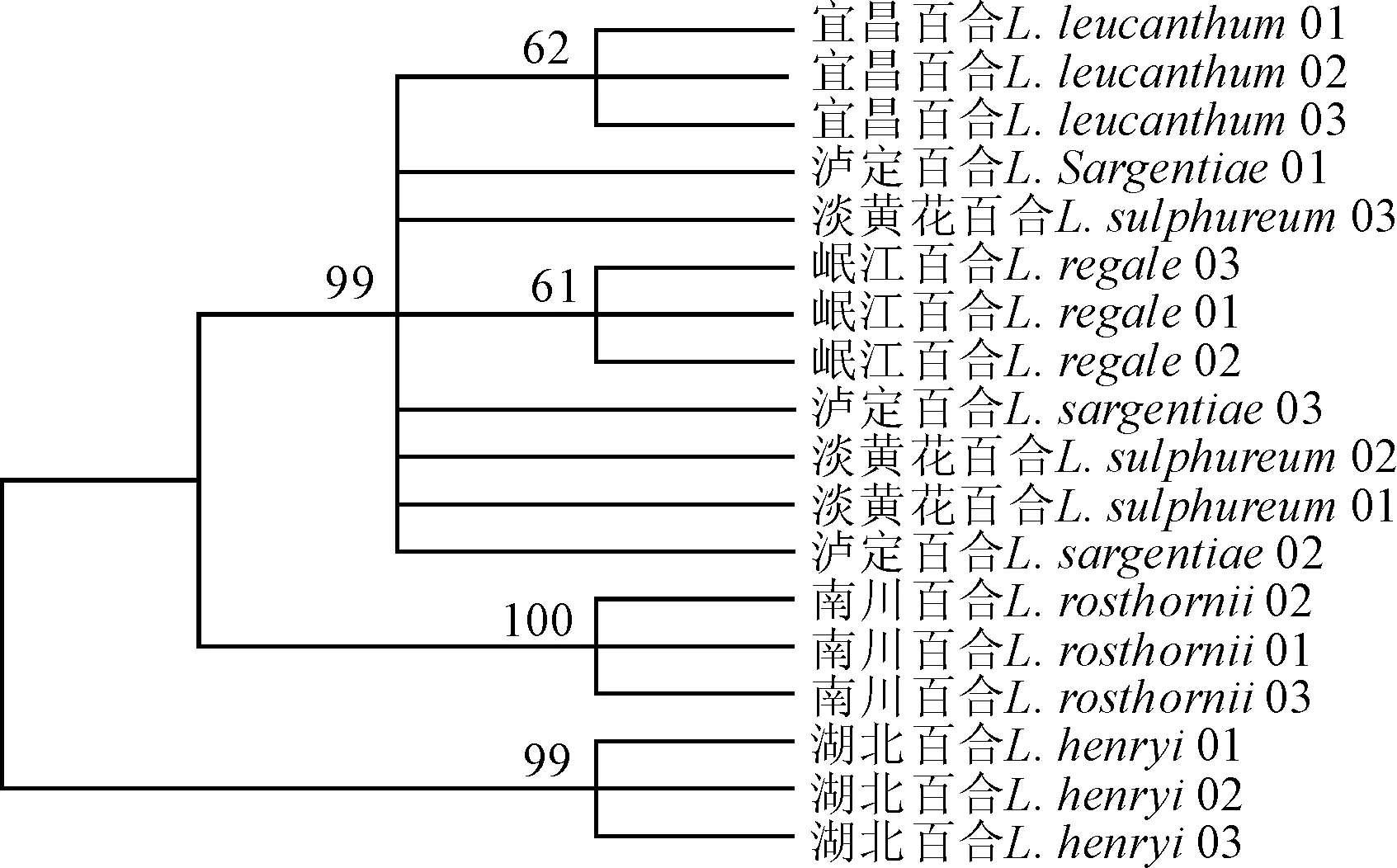
在图中标注了支持率大于50%的支系;种名后的数字表示不同居群个体编号图3 联合2个叶绿体基因(ndhC-trnV+petL-psbE)对6个近缘类群构建NJ(不包含外群)系统发育树Bootstrap values > 50% are marked. The letter plus number behind specific name represent individuals from different populationsFig. 3 Phylogenetic relationships of neighbor-joining(no outgroup) for chloroplast DNA combination (ndhC-trnV+petL-psbE) in 6 close related taxa
基于叶绿体基因ndhC-trnV和petL-psbE,选取研究类群的部分个体(不包含外类群),对其种间关系进行建树分析。NJ树聚类分析结果显示(图3),湖北百合和南川百合的不同居群各自聚为一支,并拥有较高的支持率,但二者形成一个并系的系统位置关系。同为白色管状花的4个近缘种聚为一支,得到很高的支持率(99%)。岷江百合和宜昌百合虽然也各自聚为一支,但支持率不高,嵌套在整个分支内部, 泸定百合和淡黄花百合的种间关系不确定,为平行分支交错在一起。
扩增测序的类群不一,且外类群的不同基因序列也未能获得,所以没有通过多基因联合建树来进一步验证百合属内的系统发育分析。以山丹L.pumilum为外类群,运用单拷贝核基因AKT构建的系统发育树的结果显示(图4),泸定百合、淡黄花百合、岷江百合和宜昌百合4个种很好地聚为了一支(PP/BS, 0.98/97),但它们的种间关系仍然不明确,存在一个很大的平行分支。湖北百合、南川百合与该分支的系统关系靠近,但没有获得支持率。单倍型分析结果显示,42条基因序列发现了23种单倍型(表1),从单倍型网络图上看(图5),单倍型H1是分布最广,出现频率最高的单倍型,源自泸定百合和淡黄花百合的大部分居群,共享单倍型H6、H13、H19也都是来自泸定百合和淡黄花百合居群,仅H8例外,全都来自岷江百合居群。私有单倍型丰富,H2、H3、H4、H5、H7、H15、H17、H18、H21、H23与单倍型H1位置邻近,表现出了极为亲近的位置关系。

节点处数字分别代表贝叶斯树后验概率(PP)和最大简约树Bootstrap支持率(BS),种名后字母与数字分别表示GenBank号和居群个体编号。下同图2 基于ITS序列对Leucolirion组及相关类群的系统发育分析Bayesian posterior probabilities (PP) and parsimony bootstrap (BS) are given at nodes, respectivelyThe letter plus number back of specific name represent population or individual codes. The same as belowFig. 2 Phylogeny analysis of section Leucolirion and its related taxa based on ITS sequence
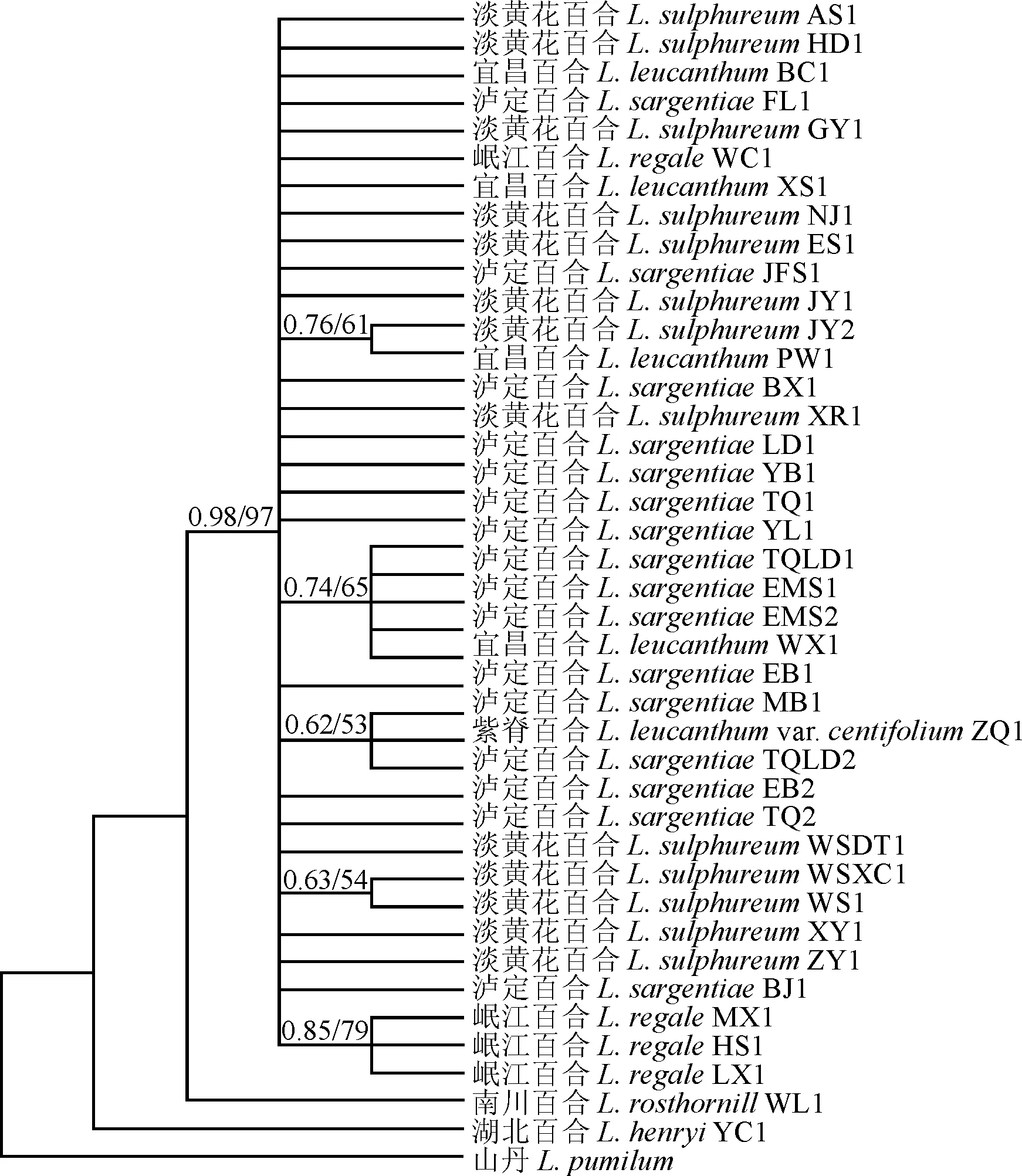
图4 基于单拷贝核基因AKT对6个近缘类群的系统发育分析Fig. 4 Phylogeny analysis of 6 close related taxa based on the nuclear single-copy sequence AKT
3 讨 论
3.1 Leucolirion组的系统发育分析
前人在世界范围内对整个百合属的系统发育做了较为全面的研究,目前的研究表明,在以分子标记所构建的百合属系统发育树中,整个百合属(含豹子花属)构成一个较为自然的单系类群,Comber[3]基于形态特征建立7个组的百合属属下分类系统得到了大多数学者的认可。然而,分子系统发育研究表明,百合属的分子系统与Comber基于形态特征建立的百合属的分类系统间存在较大的差异,Comber建立的百合属属下7个组几乎与分子没有太大的关联性。
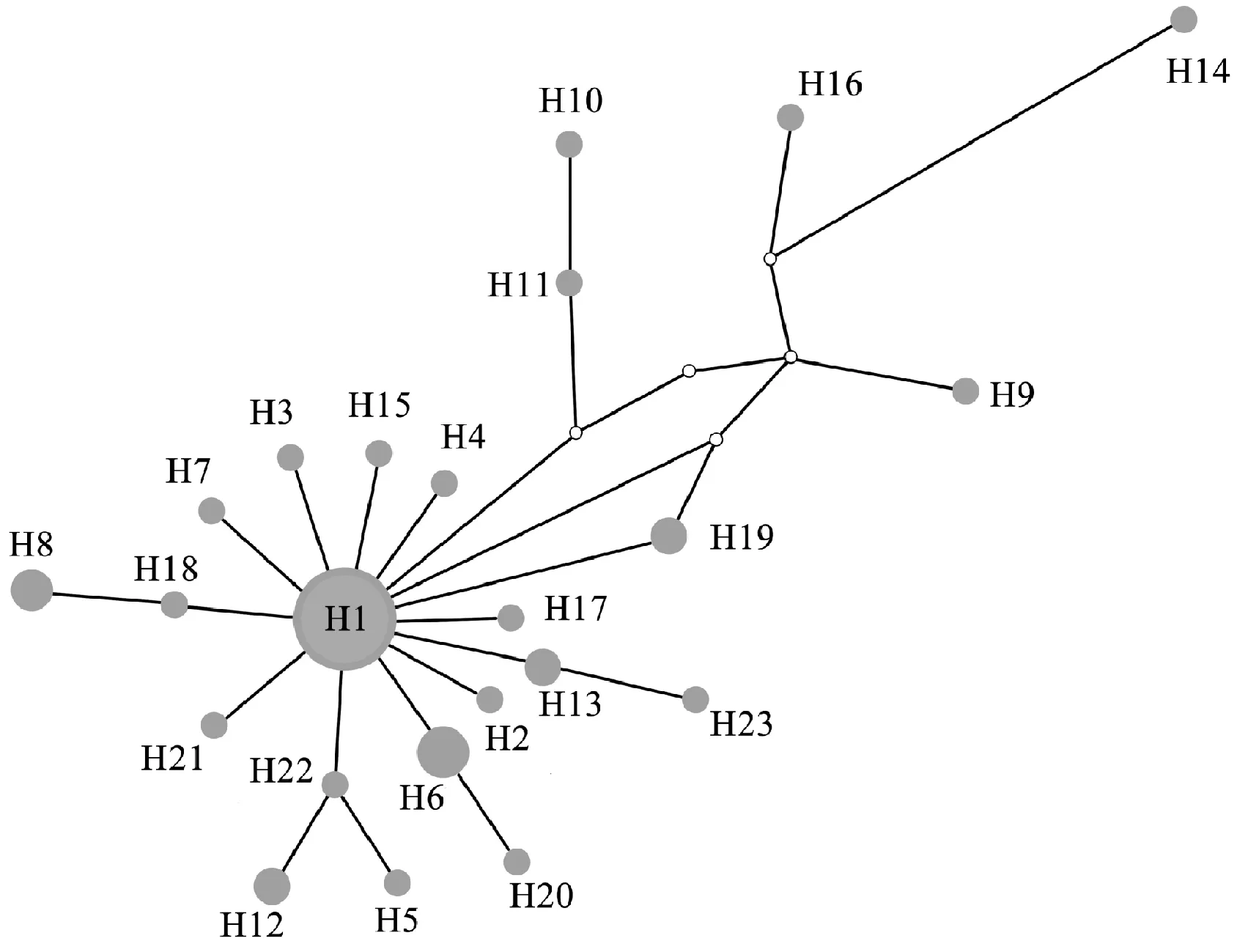
圆圈大小呈现出每一种单倍型的频率;空心的圆点代表中间单倍型,在本研究中未检测到或已经灭绝图5 基于单拷贝核基因AKT对6个近缘种构建的单倍型网络关系图The size of the circles indicates the frequency of each haplotype. Empty dots indicate that intermediate haplotypes were not detected in our research or extinctionFig. 5 Relationships wthin the haplotype network for 6 close related taxa
前人研究结果表明,湖北百合与Leucolirion组表现出了极为近缘的关系[4-5],而对南川百合的相关研究涉及较少。本研究收集6个近缘种的多个居群样品,联合相关类群对百合属进行系统发育重建。ITS树结果显示,该组划分出了2个独立分支,具有白色管状花的泸定百合、淡黄花百合、宜昌百合和岷江百合,与拥有橘黄色反卷花被片的湖北百合、南川百合分子系统关系极为亲密。这一结果与前人研究结果一致,湖北百合与南川百合应该归入到subsect.Leucolirion6a中,且二者都拥有紫色鳞茎这一特征,进一步佐证了Comber[3]基于鳞茎颜色对Leucolirion组的组下划分。而原本属于Archelirion组的野百合与subsect.Leucolirion6b聚为一支,在系统位置上与Sinomartagon组的亲缘关系更为接近。依据《中国植物志》[21]的形态分类处理,并结合分子系统发育关系,对Leucolirion组进行了重新划分整理。同意高云东[22]的提议,将泸定百合、淡黄花百合、宜昌百合、岷江百合以及新划入的湖北百合和南川百合等6个近缘种组成Leucolirion组,它们均为中国西南地区特有种,这样的处理也符合该组的形态特征(鳞茎紫色)和地理分布模式。前人的种间杂交也表明了湖北百合与泸定百合、淡黄花百合、宜昌百合、岷江百合拥有很近的亲缘关系[23-25]。将野百合与subsect.Leucolirion6b中的台湾百合、麝香百合等组成一个新组,命名为百合组(sect.Lilium),其成员大多分布在东亚沿海地区,均具有喇叭状花冠,白色鳞茎等特征。
从组间关系上看,Leucolirion组与分布在东亚沿海地区的Archelirion组形成姐妹类群,而百合组在系统位置上与Sinomartagon组更为接近[2,10],这一组间关系在种间杂交试验也得到了支持。通常,亲缘关系更近的物种,杂交成功率更高。园艺上的东方百合杂交株系便是从Archelirion组内的L.speciosum、L.japonicum与湖北百合的杂交种中选育而来。从遗传学角度上说,分子系统分类的结果更加准确可靠,更加稳定。而基于形态所作出的分类结果则依据对分类特征侧重点不同而表现出较大的差异。
3.2 近缘类群的种间关系
本研究试图对所选取的6个近缘种的种间系统关系进行进一步的确认。从叶绿体联合基因和单拷贝核基因AKT计算得到的种间遗传距离来看,湖北百合和南川百合虽然与泸定百合、淡黄花百合、岷江百合、宜昌百合聚为一支,但不管在形态上还是遗传距离上都与后4个种差异明显。此外,湖北百合和南川百合的种间界限模糊,二者的形态十分相似,都具有橘黄色反卷花被片、紫色鳞茎和两型叶等特征;对这二者的种间关系很少有报道,Du等[10]利用荧光原位杂交技术在染色体水平上说明了两者的区别在于B染色体的有无上,然而B染色体这一结构并不能作为不同种的分类依据,因为它的存在与否更多与环境条件相关[22,26]。湖北百合主要分布于贵州、重庆和湖北境内的武陵山山脉及巫山山脉的崖壁生境上,居群分布格局相对隔离,而南川百合仅发现于重庆和湖北交界处的一些细小河流的石隙或者湿泥之中,沿河流生长分布,二者在生境选择上有很大的差异。本研究基于ITS构建的系统发育树显示,虽然二者能聚为一个独立的分支,但从多个居群取样的结果并不能很好地加以区分。在叶绿体联合基因树中,来自不同居群的2个种的个体很好地各自聚类在一个分支内,但在系统位置关系上是一个并系分支,说明了二者在遗传水平上存在着广泛的交流。形态上,湖北百合与南川百合主要区别在于叶片与果实的大小;然而,分子水平上二者却表现出了极高的相似性,遗传距离很小,关于这2个种的种间界限还需要进一步讨论。
同为白色管状花的泸定百合、淡黄花百合、岷江百合、宜昌百合在形态上十分相近,但系统位置一直相对稳定。叶绿体联合基因树并未能很好地解释它们之间的关系,4个类群聚为一个大的平行分支,互为姐妹类群关系,并且具有较高的支持率。这说明了这4个类群的单系性,亲缘关系密切。单拷贝核基因的MP树和单倍型分析也同样不能区分这4个类群。从地理分布上看,该4个类群的分布区域邻近,花期物候十分相似,为种间的自然杂交提供了可能,促进了它们的基因交流,从而在基因水平表现出很强的相似性,这一特点在百合科顶冰花属类群中也存在[27]。特别强调的是泸定百合和淡黄花百合,这二者从地理分布上看最为广泛,且没有明显地理阻隔,形态差异极小,都能产生珠芽进行无性繁殖,这加速了二者的扩张与传播,在前人研究报道中,经常把二者混淆,所以他们的种间关系还需要进一步探讨。
本研究的6个近缘类群的种内遗传变异相差很小,在基因水平上遗传差异不明显。居群间的遗传分化和遗传结构往往与基因流、花粉种子的传播以及繁育系统等因素相关[28]。百合属植物拥有较为复杂而又健全的繁殖机制,能以鳞茎进行无性生殖,这使得有性生殖过程减少,为百合的基因库保存提供了保障,但同时也放缓了基因变异与分化的速率,使百合属植物在基因水平上呈现出了极高的相似性,种间的遗传差异不明显,给种间关系的鉴定上造成了极大的困难。
综上所述,本研究基于3种分子序列分析了百合属下Leucolirion组的系统发育关系,明确了Leucolirion组存在2个独立的分支,将湖北百合、南川百合、泸定百合、淡黄花百合、岷江百合和宜昌百合等6个近缘种归入到同一个组中,构成Leucolirion组。其组内6个近缘类群的种间关系仍不明确,需要借助更多分子手段进行更为深入的研究。
[1] LIANG S Y, TAMURA M N. Flora of China[M]. Beijing: Science Press, 2000, 24: 135-149.
[2] GAO Y D, HARRIS A J, ZHOU S D,etal. Evolutionary events inLilium(includingNomocharis, Liliaceae) are temporally correlated with orogenies of the Q-T plateau and the Hengduan Mountains[J].MolecularPhylogeneticsandEvolution, 2013, 68(3): 443-460.
[3] COMBER H F. A new classification of genusLilium[J].LilyYearbook,RoyalHorticulturalSociety(London), 1949, 15: 86-105.
[4] NISHIKAWA T, OKAZAKI K, UCHINO T,etal, NAGAMINE T. A molecular phylogeny ofLiliumin the internal transcribed spacer region of nuclear ribosomal DNA[J].JournalofMolecularEvolution, 1999, 49: 238-249.
[5] NISHIKAWA T, OKAZAKI K, ARAKAWA K,etal. Phylogenetic analysis of sectionSinomartagonin genusLiliumusing sequences of the internal transcribed spacer region in nuclear ribosomal DNA[J].BreedingScience, 2001, 51(1): 39-46.
[6] HAYASHI K, KAWANO S. Molecular systematics ofLiliumand allied genera (Liliaceae): phylogenetic relationships amongLiliumand related genera based on therbcLandmatKgene sequence data[J].PlantSpeciesBiology, 2000, 15(1): 73-93.
[7] GAO Y D, ZHOU S D, HE X J. Karyotype studies in thirty-two species ofLilium(Liliaceae) from China[J].NordicJournalofBotany, 2011, 29(6): 746-761.
[8] LEE C S, KIM S C, YEAU S H,etal. Major lineages of the genusLilium(Liliaceae) based on nrDNA sequences, with special emphasis on the Korean species[J].JournalofPlantBiology, 2011, 54(3): 159-171.
[9] GAO Y D, HOHENEGGER M, HARRIS A,etal. A new species in the genusNomocharisFranchet (Liliaceae): evidence that brings the genusNomocharisintoLilium[J].PlantSystematicsandEvolution, 2012, 298: 69-85.
[10] DU Y P, HE H B, WANG Z X,etal. Molecular phylogeny and genetic variation in the genusLiliumnative to China based on the internal transcribed spacer sequences of nuclear ribosomal DNA[J].JournalofPlantResearch, 2014, 127(2): 249-263.
[11] DOYLE J. A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J].Phytochem.Bull., 1987,19:11-15.
[12] WHITE T J, BRUNS T, LEE S,etal. Amplifications and direct sequencing of fungal ribosomal RNA genes for phylogenetics[M]// INNIS M A, GELFAND D H, SNINSKY J J. PCR Protocols: A Guide to Methods and Applications. San Diego: Academic Press, 1990: 315-322.
[13] SHAW J, LICKEY E B, SCHILLING E E,etal. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: the tortoise and the hare III[J].AmericanJournalofBotany, 2007, 94(3): 275-288.
[14] DOUGLAS N A, WALL W A, XIANG Q Y,etal. Recent vicariance and the origin of the rare, edaphically specialized Sandhills lily,Liliumpyrophilum(Liliaceae): evidence from phylogenetic and coalescent analyses[J].BmcComplementary&AlternativeMedicine, 2011, 20(14): 2 901-2 915.
[15] BURLAND T G. DNASTAR’s Lasergene sequence analysis software[J].MethodsinMolecularBiology, 2000, 132: 71-91.
[16] TAMURA K, DUDLEY J, NEI M,etal. MEGA4: Molecular evolutionary genetics analysis (MEGA) software version 4.0[J].MolecularBiology&Evolution, 2007, 24(8):1 596-1 599.
[17] MORRISON D A. A framework for phylogenetic sequence alignment[J].PlantSystematics&Evolution, 2009, 282(3):127-149.
[18] SWOFFORD D L. PAUP* 4.0: Phylogenetic Analysis Using Parsimony (*and Other Methods)[M]. Sunderland: Sinauer Associates, 2001.
[19] FREDRIK R, HUELSENBECK J P. MrBayes 3: Bayesian phylogenetic inference under mixed models[J].Bioinformatics, 2003, 19(12): 1 572-1 574.
[20] NYLANDER J. MrModeltest 2.0: Program distributed by the author[M]. Evolutionary Biology Centre, Uppsala University, 2004.
[21] 陈启心,许介眉,梁松筠.中国植物志[M].北京:科学出版社,1980:116.
[22] 高云东.百合属及其近缘属植物的核型进化及百合属的分子系统学、生物地理学研究[D].成都:四川大学,2013.
[23] LIGHT R W. Cytological and interspecific hybridization studies inLiliumL. and their significance for classification[D]. Ithaca: Cornell University, 1960.
[24] SYNCE P M. Lilies[M]. London: Batsford, 1980.
[25] WOODCOCK H B D, STEARN WT. Lilies of the world[M]. London/New York: Country Life/Scribner, 1950.
[26] 洪德元.植物细胞分类学[M].北京:科学出版社,1990.
[27] PETERSON A, JOHN H, KOCH E,etal. A molecular phylogeny of the genusGagea(Liliaceae) in Germany inferred from non-coding chloroplast and nuclear DNA sequences[J].PlantSystematics&Evolution, 2004, 245(3): 145-162.
[28] PETERSON A, BARTISH I V, PETERSON J. Genetic structure detected in a small population of the endangered plantAnthericumliliago(Anthericaceae) by RAPD analysis[J].Ecography, 2002, 25(6): 677-684.
(编辑:宋亚珍)
Phylogeny and Interspecific Relationship ofLiliumSectionLeucolironBased on Three Gene Sequences
LAI Shanpan, SHEN Chengjuan, ZHOU Songdong, HE Xingjin*
(Key Laboratory of Bio-Resources and Eco-Environment of Ministry of Education, College of Life Science, Sichuan University, Chengdu 610064, China)
Despite abundant germplasm resources of wild lilies in China, the study of phylogenetic and interspecific relationship inLiliumwas scarce. In this study, phylogeny of the sectionLeucolirionComber which includes GenBank data and 48 specimens covering 6 closed related lilies endemic to Southwest China was explored based onITS, cpDNA and a single-copy nuclear gene,AKT. The results showed that: (1) in accordance with previous studies that the sectionLeucolirionComber should be divided into two subsection, subsect.Leucolirion6a and subsect.Leucolirion6b, andL.henryiandL.rosthorniishould be classified into subsect.Leucolirion6a. We revise sectionLeucolirionincludingL.sargentiae,L.sulphureum,L.leucanthum,L.regale,L.henryiandL.rosthornii.L.bownii, which originally belonged to sectionArchelirionwas placed into subsect.Leucolirion6b, along withL.formosanumandL.longiflorum, we establish a new group, named sectionLilium. (2)As for infrageneric relationships, sectionArchelirionwhich mainly distributes in Japan showed a sister relationship with sectionLeucolirion. However, sectionLiliumandSinomartagonshowed a close relationship. (3) The genetic distances among all taxa of chloroplast combination andAKTshowed thatL.henryiandL.rosthorniidistinctly differ from other members of sectionLeucolirion. Phylogenetic tree based on chloroplast combination data,AKTfragment indicated thatL.sargentiae,L.sulphureum,L.regaleandL.leucanthumhave a very close relationship. High genetic similarity between these species may be attributed to the particular mating system ofLilium, leading to limited genetic variation and differentiation among species.
Leucolirion; phylogeny;ITS; chloroplast gene; single-copy nuclear gene; interspecific relationship
1000-4025(2016)08-1541-10
10.7606/j.issn.1000-4025.2016.08.1541
2016-04-25;修改稿收到日期:2016-06-12
国家自然科学基金(31270241, 31470009, 31570198, 31110103911);国家基础条件平台;国家标本平台教学标本子平台(http://mnh.scu.edu.cn);科技部基础性研究专项(2013FY112100)
赖山潘(1990-),男,在读硕士研究生,主要从事植物系统进化研究。E-mail: laistephen@126.com
*通信作者:何兴金,教授,博士生导师,主要从事植物分类与系统进化相关方面的研究。E-mail: xingjinhe@aliyun.com
Q789
A
