生物乙烯研究进展
2016-10-13孙梦婷范晓蕾郭荣波邱艳玲赵晓娴
孙梦婷范晓蕾郭荣波邱艳玲赵晓娴
(1.山东省沼气工业化生产与利用工程实验室 中国科学院青岛生物能源与过程研究所生物燃料重点实验室,青岛 266101;2.中国科学院大学,北京 100049)
生物乙烯研究进展
孙梦婷1,2范晓蕾1郭荣波1邱艳玲1赵晓娴1
(1.山东省沼气工业化生产与利用工程实验室 中国科学院青岛生物能源与过程研究所生物燃料重点实验室,青岛 266101;2.中国科学院大学,北京 100049)
乙烯是世界需求量最大的化工原料,被广泛用于制造塑料制品、纺织品及其他化工产品。目前,最常用的生产乙烯的方式是通过石油裂解制备乙烯,但其存在生产成本高、环境不友好等弊端,这为生物乙烯的发展带来契机。生物乙烯合成以CO2或生物质等可再生能源为原料,具资源节约、环境友好的优势。描述了生物乙烯合成的两种途径──间接和直接合成途径,着重论述微生物通过乙烯合成酶(EFE)直接合成乙烯的原理,描述了EFE的结构、序列及催化机制,列举了数个工程菌异源合成乙烯的成功案例,提出提高产量的策略,并对微生物制乙烯的发展前景进行展望。
生物乙烯;直接合成;微生物;乙烯合成酶;异源表达
乙烯是重要的化工原料,主要用于生产聚乙烯、环氧乙烯、二氯乙烷等,其生产规模、技术水平和产量可衡量一个国家的化工工业水平[1]。目前,多数国家通过石脑油、瓦斯油等裂解制备乙烯。近年,中国乙烯生产量和自给率不断提高,但仍不及需求的增长速度,并且,随石油资源的日益枯竭和价格攀升,乙烯的传统生产成本日益增加,这为生物乙烯提供了发展契机[2]。
乙烯生物合成以生物质为原料,利用生物体生长代谢活动,通过直接或间接方法获得乙烯[3,4]。间接合成以可再生生物质为原料,通过微生物发酵生成乙醇,催化剂作用于乙醇脱水生成乙烯。间接合成经多年发展已实现产业化,并有效缓解了能源紧缺问题,但其存在弊端,如产力低、工艺不合理、规模小等[5]。直接合成是生物体以有机物或CO2为原料在体内直接合成乙烯,通过生物工程手段将其引入微生物来生产乙烯,投资小、环境友好、产品纯度高,是切实可行的新型合成路线[6]。
目前,国内外生物乙烯研究多集中在间接合成的成本降低和优化生产工艺上,并未足够重视直接合成。本文在之前研究基础上,综述了生物制乙烯技术现状,着重论述直接合成法生物学基础和异源表达案例,并对生物乙烯发展前景进行展望。
1 生物乙烯合成
1.1 生物乙烯间接合成
间接合成起步较早,已具备较成熟的生产工艺和流程,具体如图1-A所示,蔗糖、淀粉和纤维素等水解生成葡萄糖等还原糖,经微生物糖酵解生成乙醇,再在催化剂作用下脱水生成乙烯。生物乙醇来源于玉米、甘蔗、甜高粱和甜菜等作物,由此带来成本高和粮食安全问题,因此,近年来国内外研究多集中于降低乙醇制造成本和非粮乙醇方面,将植物秸秆中的纤维素、半纤维素降解并供给微生物发酵生产乙醇,成为未来生物乙醇主要生产模式。目前,间接合成法最成熟的催化剂是活性氧化铝,其稳定性好、寿命长,但存在反应温度高、进料空速低等不足,经改良的HZSM-5、SAPO-34分子筛类催化剂及杂多酸催化剂等,性能优于前者,但仍无法满足商业化需求。因此,开发高性能新型催化剂成为间接合成法的关键所在。乙醇脱水生成乙烯有绝热床和等温床两种工艺,均存在转化率低、能耗高等不足,有待优化。
与石油裂解制乙烯相比,间接合成具以下优点:(1)原料来源广,为可再生生物质;(2)反应条件温和;(3)工艺流程简单;(4)能耗低;(5)投资小;(6)环境友好。但其仍存在生物乙醇生产成本高、生产规模小、催化剂性能有待提高、反应器设计不合理、生产工艺待优化等劣势[7-9]。
1.2 生物乙烯直接合成
许多生物体具乙烯合成功能,包括一些植物、微生物和动物,其分别采取不同的乙烯合成途径[10-12]。直接合成法将适当的乙烯合成途径通过生物工程手段引入微生物,使微生物以有机物或CO2为原料合成乙烯,与间接合成法相比,其具有显著优势:(1)设备少,不需催化剂;(2)成本低,原料来源广;(3)能耗低,CO2排放少,环境友好(光合自养菌可通过光合作用固定CO2转化为乙烯);(4)产物纯度高,副产物少;(5)产率受O2和营养条件影响,生产过程可实现优化控制。
2 生物乙烯直接合成原理
2.1 植物合成乙烯
2.1.1 植物合成乙烯途径 在高等植物中,如图1-b所示,乙烯合成前体为甲硫氨酸(Methionine,Met),关键酶为ACS(ACC合酶)和ACO(ACC氧化酶)。反应过程如下:Met在S-腺苷甲硫氨酸合成酶(S-Adenosylmethionine synthetase,SAMS)作用下生成S-腺苷甲硫氨酸(S-adenosylmethionine,SAM),ACS催化SAM生成5'-甲硫基核糖(5'-methylthioribose,MTR)和1-氨基-1-羧基环丙烷(1-aminocyclopropane-1-carboxylic acid,ACC),MTR通过甲硫氨酸循环参与Met再生,ACC则在ACO催化下生成氰化物并放出乙烯[13]。
2.1.2 植物合成乙烯的调控 植物合成乙烯关键酶是ACS和ACO。ACS催化SAM转化为ACC,是限速步骤。ACS以磷酸吡哆醛为辅基,由多基因家族编码,其基因表达和催化活性响应于内外部各种刺激,乙烯积累可抑制ACS基因表达,自身mRNAs积累增强ACS催化活性。ACO催化ACC生成乙烯,其需要抗坏血酸和氧作辅助底物,Fe2+和CO2作辅助因子,解偶联剂和自由基清除剂能抑制乙烯生成。ACO由多基因家族编码,乙烯、Cu2+、NaCl、ACC等可诱导ACO表达[14,15]。Hamilton等[16]使用番茄植株ACO1基因转化酿酒酵母后,可将ACC转化为乙烯。乙烯合成还受信号转导通路调节,拟芥南中存在5个乙烯受体:ETR1、ERS1、ETR2、ERS2和EIN4,它们均是乙烯合成的负调控因子[17]。植物合成乙烯途径调控机制复杂,易受干扰,为产业化生产带来困难。
2.2 微生物合成乙烯
2.2.1 微生物合成乙烯途径 微生物合成乙烯途径分为两条:一是KMBA途径,发生在大肠杆菌(E. coli)、浅白隐球酵母(C. albidus)等细菌中,如图1-C所示,Met通过转氨作用生成中间产物2-酮-4-甲基硫代丁酸(2-keto-4-methylthiobutyric acid,KMBA),KMBA被氧化裂解放出乙烯[18]。二是EFE途径,发生在丁香假单胞杆菌(P. syringae)、青霉菌(P. digitatum)等微生物中,如图1-D所示,谷氨酸(Glu)脱氨生成 α-酮戊二酸(α-ketoglutarate,AKG),AKG在乙烯合成酶(ethylene-forming enzyme,EFE)作用下生成乙烯。由于后者的乙烯合成速率远高于前者,因此EFE途径是微生物直接合成乙烯的研究重点。
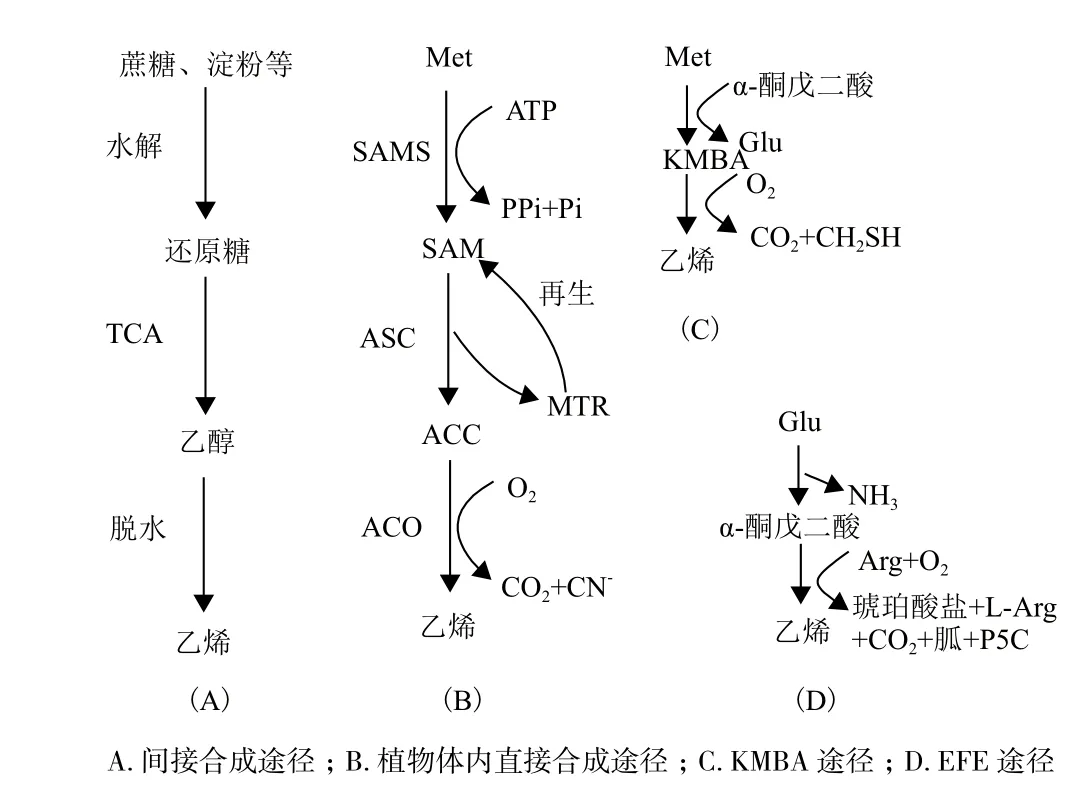
图1 生物乙烯间接与直接合成途径
2.2.2 EFE的发现、结构和efe基因 1940年,Yang和Biale等[19,20]首次报道青霉菌(P. digitatum)产乙烯,青霉菌成为首个被鉴定的产乙烯微生物。自青霉菌提纯EFE,测得EFE是相对分子质量为42 kD的单体蛋白,在Fe2+和O2存在下,催化AKG和精氨酸(Arg)生成琥珀酸盐、胍类、吡咯啉-5-羧酸(P5C)、CO2和乙烯。丁香假单胞菌(P. syringae)是目前已知的产乙烯效率最高的微生物。提取丁香假单胞菌的EFE进行检测,其相对分子质量为42 kD且为单体蛋白,在Fe2+、O2、Arg和AKG存在下催化乙烯生成,这与青霉菌中EFE性质基本一致[21]。
实验表明,青霉菌和丁香假单胞菌的EFE的N端序列不同,但两者具一整段相似保守序列,推测这段序列是EFE催化功能所必需的。EFE功能域相对保守,定向诱变丁香假单胞菌efe基因,以Glu残基取代His残基,使其在大肠杆菌中表达,分离纯化EFE后进行试管实验,检测其Km值、kcat值及热稳定性。实验结果表明,His305和His335位点发生突变的酶,活性分别减少至原活性水平的40%和60%,His189和His233位点发生突变的酶则完全失活,表明这两个位点在络合铁离子时起关键作用,在His268位点突变的酶未完全失活,但活性急剧下降,表明此位点突变破坏了EFE活性部位。除此之外,EFE还含有两个保守的氨基酸残基Asp191和Arg277,对酶催化活性起重要作用[22]。
上述实验将丁香假单胞菌efe基因在大肠杆菌中克隆并表达,观测到乙烯生成,这表明在不同菌株中,单个efe基因是产生乙烯的充分条件,因此,是否具efe基因可确定待测菌株是否具产乙烯能力。Sato等[23]分离丁香假单胞菌突变株P. syringae pv. phaseolicola PK2(Kudzu菌株,产乙烯菌株)efe基因作引物,对P. syringae pv. cannabina 和P. syringae pv. sasemi 两种菌株基因进行PCR扩增,它们的PCR产物均包含大小与efe基因相似的片段,以Kudzu菌株efe基因作DNA探针与其PCR产物杂交,结果显示产物中均含efe同源基因,据此表明两菌株均具产乙烯能力。
2.2.3 EFE催化机制和化学计量分析 对青霉菌和丁香假单胞菌的EFE进行试管实验,测反应化学计量数,得化学方程式如下:
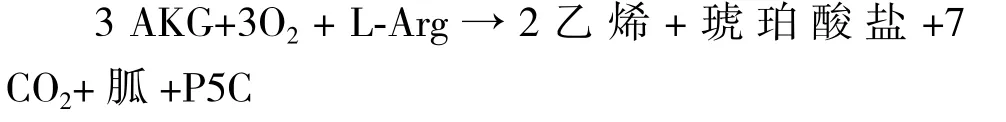
反应底物AKG和L-Arg比例为3∶1,产物乙烯、琥珀酸盐和P5C比例为2∶1∶1。EFE同时催化两个反应发生,主反应是2分子AKG被氧化生成2分子乙烯和6分子CO2,副反应是1分子AKG转化为琥珀酸盐和CO2各1分子的同时,Arg转化为胍类和P5C。如果EFE是可催化两个反应的复合酶,并且两反应可分离,那么利用工程化手段改造EFE,使其仅催化主反应,不催化副反应,不生成副产物,将会大大提高产率和反应物利用率。因此,关于EFE功能结构及催化机制应当深入研究。
2.2.4 EFE反应与胞内碳流分析 对EFE反应所涉及的新陈代谢通路进行分析,优化反应途径,使反应产物与底物摩尔比最大化,降低副产物量、能量和辅因子消耗,从而使EFE反应有序高效进行成为可能。
乙烯的碳来源大体可分为3类:一是CO2固定,存在于光合自养型生物中,CO2被核酮糖1,5-二磷酸加氧酶(Rubisco)固定,通过卡尔文循环进入EFE反应,消耗ATP和还原剂转换为乙烯;二是糖酵解,己糖在NAD(P)+依赖性的甘油-3-磷酸脱氢酶、异柠檬酸脱氢酶、丙酮酸脱氢酶等催化下分解,其中3个碳进入EFE反应生成乙烯;三是磷酸戊糖途径,木糖或混合糖通过磷酸戊糖途径进入EFE反应产生乙烯,还原产物和碳产量与糖酵解相近。
3条途径中,CO2固定途径虽需大量ATP和还原剂,但为碳负过程,另两种途径虽所需辅因子较少,但代谢过程中存在碳丢失,因此,应当找到有效策略来挽救丢失的碳原子[24]。
除确定最佳代谢途径外,阻碍途径和竞争途径也应当被纳入研究范畴,通过流量平衡分析(FBA)和代谢流分析(MFA)找到影响乙烯产量的因素。Larsson等[25]对efe基因转化的酿酒酵母进行FBA,研究表明添加外源脯氨酸(Pro)、使用NAD依赖型谷氨酸脱氢酶(NAD-GDH)代替NADH依赖型谷氨酸脱氢酶(NADH-GDH)、使用谷氨酸盐代替铵盐作氮源均可增加乙烯产量。另外,通过改变O2量来限制其呼吸作用,同样会影响乙烯产量。
2.3 动物合成乙烯
哺乳动物可合成少量乙烯,有报道显示,大鼠肝脏中存在KMBA途径合成乙烯。由于动物合成乙烯量少,其对产业化生产乙烯并无太大价值。
3 生物乙烯异源合成案例
已有研究表明,细菌efe基因在大肠杆菌(E. coli)、酿酒酵母(Saccharomyces cerevisiae)、蓝细菌聚球藻属(Synechococcus)、蓝细菌集胞藻属(Synechocystis)和木霉(Trichoderma)等菌株中实现了异源表达,并使转化后的工程菌株获得乙烯合成能力。
3.1 假单胞菌合成乙烯
目前已知产乙烯微生物中,丁香假单胞菌产量最高,其中Kudzu菌株和丁香假单胞菌大豆致病突变株(P. syringae pv. glycinea)是研究最多的菌株。为进一步提高乙烯产量,Ishihara等[26]构建了高拷贝数、广谱宿主质粒载体pMEFE1,其上携带来源于Kudzu菌株的efe基因,将pMEFE1质粒分别导入丁香假单胞菌(P. syringae)、恶臭假单胞菌(P. putida)、大肠杆菌JM109后,重组细胞均具EFE合成能力,并且乙烯产量比亲代丁香假单胞菌分别高出41倍、20倍和5.3倍。Wang等[27]将丁香假单胞菌efe基因重组至恶臭假单胞菌KT2440的16S rRNA位点,PCR结果显示转化后菌株DC1、DC2、DC3分别具有3、4和5个efe基因拷贝,其乙烯产量分别为36.2、47.2、53.8 mg/(h·g),与亲代丁香假单胞菌14.7 mg/(h·g)的产量相比大幅增加。将携带efe基因的pBBR1MCS2质粒载体转入DC3菌株以继续增加efe拷贝数,乙烯合成量达到80.2 mg/(h·g),进一步说明efe基因拷贝数与乙烯合成量呈正相关。对重组菌乙烯合成动力曲线进行分析,乙烯产量随时间逐渐增高,7 h达到峰值,之后开始下降,继续培养至17 h添加0.25%葡萄糖溶液,产力比7 h时提高40%,这说明碳源葡萄糖是乙烯合成的限制性因子。
3.2 大肠杆菌异源合成乙烯
efe基因最早在大肠杆菌中异源表达。来自丁香假单胞菌Kudzu菌株的efe基因被克隆,通过高拷贝pUC19载体在大肠杆菌中表达。实验将转化后的大肠杆菌分别在25℃和30℃下培养,结果表明,25℃下乙烯产量较高,推测37℃时EFE易形成包含体从而活性降低。另外,将转化后的大肠杆菌在添加外源底物的试管内反应,测得乙烯生成量为相同条件下细胞内反应(即底物仅来自于胞内)的5倍,这表明底物利用率与乙烯产量呈正相关。
3.3 酿酒酵母异源合成乙烯
2008年,Pirkov等[28]将丁香假单胞菌efe基因转入酿酒酵母中,使其利用葡萄糖为碳源合成乙烯,产量达到(320±90)μg/g葡萄糖。2011年,Larsson等[25]对酿酒酵母乙烯合成进行代谢流分析,结果表明,使用NAD-GDH代替NADH-GDH可有效提高乙烯产量,这是由于谷氨酸脱氢酶(GDH)催化谷氨酸盐脱氨产生乙烯合成底物AKG,GDH分为NAD和NADP依赖型两种,NADPH主要来源于磷酸戊糖途径,该途径存在碳丢失,使碳源减少,AKG水平下降,从而使乙烯产量减少。实验以半乳糖替代葡萄糖作碳源,产量急剧下降,说明酵母合成乙烯的最佳碳源是葡萄糖。除此之外,由于乙烯合成副产物P5C是Pro合成的中间体,添加外源脯Pro可使P5C转而合成Glu,提高乙烯产量。为确认影响乙烯产量因素,2013年,Johansson等[29]对酿酒酵母在不同条件下乙烯产量进行测定。分析表明,O2量影响乙烯产量,由于O2维持细胞呼吸作用,并重新氧化NADH用于乙烯合成,是重要限制性因子。实验以谷氨酸盐代替铵盐作氮源,乙烯合成速率大大提高。2014年,Johansson等[30]继续对酵母乙烯合成的氮代谢进行分析,以铵盐、谷氨酸盐、Arg、谷氨酸盐/Arg分别作氮源。结果表明,以谷氨酸盐作氮源乙烯产量最高,这是由于谷氨酸盐在GDH作用下转化为乙烯合成底物AKG;以Arg作氮源产量减少,但Arg缺陷型突变株完全没有乙烯生成,表明虽然Arg作氮源产量较少,但却是乙烯合成所必需。因此,高浓度AKG和一定数量Arg是乙烯产量最大化所必需。综上可得,利用酵母合成乙烯,其产量受到碳源、氮源、O2量、底物浓度、辅因子NADP等影响。
3.4 木霉异源合成乙烯
木霉是一种丝状真菌,其内富含丰富的纤维素酶系,能将纤维素降解为葡萄糖,这意味着,一旦木霉具乙烯合成能力,便能利用农业废弃物中的纤维素为碳源生产乙烯。2008年,Tao等[31]首次将丁香假单胞菌efe基因转入绿色木霉菌(Trichoderma viride)中,得到具乙烯合成能力的转化菌株Y2,速率达1 059.7 nL/(L·h)。同样,乙烯产量受碳源和氮源影响:以纤维素作碳源,Y2菌株乙烯合成速率最高,以羧甲基纤维素钠或乳糖做碳源,产量减少;以蛋白胨作氮源,乙烯产量升高。另外,用包含小麦秸秆的固体培养基培养Y2菌株,可获得较高乙烯产量。2011年,Chen等[32]将丁香假单胞菌efe基因转入里氏木霉菌(Trichoderma reesei)中,观察到乙烯合成量随efe表达量增加而增加,以稻草秸秆为碳源培养转化菌株,乙烯合成量达到4 012 nL/(L·h)。以上两项研究均表明,efe基因转化的木霉菌可利用植物废弃物转化成乙烯,这使乙烯低碳环保生产成为可能。
3.5 蓝细菌异源合成乙烯
与大肠杆菌、酵母、木霉等产乙稀工程菌株相比,产乙稀工程蓝细菌具有明显的优势:第一,乙烯合成速率较高;第二,蓝细菌可通过光合作用将CO2转化为乙烯,将乙烯生产与CO2减排有机结合,真正实现了乙烯的绿色合成;第三,可利用海水培养基对蓝细菌进行培养,培养基廉价易得,资源友好。因此,我们认为蓝细菌光合产乙稀将是最具发展前景的生物乙烯生产方式。早在1997年,Sakai和Fukuda等[33]将丁香假单胞菌Kudzu菌株的efe基因转入蓝细菌聚球藻Synechococcus sp. PCC 7942中,使其具有产乙烯能力,但随着细胞分裂,由于质粒pEXE3和宿主染色体psb AI位点发生同源重组,efe基因拷贝数减少,使得菌株乙烯合成量急剧下降。2003年,Takahama等[34]将丁香假单胞菌efe基因直接插入蓝细菌聚球藻Synechococcus elongatus PCC 7942的psb AI位点,使转化菌株获得451 nL/(mL·h·OD730)的乙烯合成能力,但聚球藻生长和代谢速率随时间减慢,相应乙烯合成量也减少。2012年,Ungerer等[35]将Kudzu菌株efe基因修饰后转入蓝细菌集胞藻Synechocystis sp. PCC 6803中,EFE稳定性大幅提高,使菌株能够持续合成乙烯,最高乙烯合成速率达5 650 μL/(L·h),并观察到高达5.5%的碳进入乙烯合成途径。代谢途径优化是提高蓝细菌产乙烯能力的另一重要途径。本研究单位朱涛等[36]前期研究表明,经过TCA循环途径的修饰和优化,在抑制α-酮戊二酸脱羧酶(OGDC)和琥珀酸半醛脱氢酶(SSADH)表达的同时,将大肠杆菌中编码α-酮戊二酸透过酶(KgtP)的基因导入蓝细菌phaAB位点,从而增加细胞内底物α-酮戊二酸的量,结合过量表达EFE,能够大大提高蓝细菌工程菌株的乙烯合成能力,合成速率达到了9.7 mL/(L·h)。
4 产物乙烯的分离与纯化
产物分离与纯化是乙烯生产的关键步骤。在传统石油化工领域,乙烯分离技术有深冷分离法、吸附分离法、膜分离法、膨胀机法、金属络合分离法及联合工艺等。生物制乙烯与石油裂解制乙烯的产物组分不同,通常,前者的产物中除乙烯外,还存在CO2,水蒸气、N2、O2等,但不存在石油裂解过程中产生的金属及其他污染物。另外,光合自养产乙烯微生物通过光合作用产生O2,乙烯在一定浓度O2中具可燃性,这也是分离生物乙烯时必须考虑的因素[6,27]。因此,在设计生物乙烯分离与纯化工艺过程中,不仅要运用和借鉴石油裂解制乙烯的分离、纯化工艺,还要充分考虑生物制乙烯的产物组分和性质,从而设计出安全、高效的分离与纯化技术。
4.1 深冷分离法
该技术的分离流程包括气体净化系统、压缩冷却系统和精馏分离系统,具体过程是利用产物混合气中各组分相对挥发度的差异,低温下将各组分按工艺要求冷凝下来,再用精馏法将各类产物逐一分离,从而实现乙烯的分离与纯化。此方法乙烯回收率较高,可用于大规模生产。
4.2 吸附分离法
该方法利用吸附剂对混合气中各组分的吸附选择性不同,通过改变压力或温度来实现分离,分为变压吸附法、变温吸附法和变温变压吸附法。按所用载体不同,吸附剂分为分子筛类、树脂类、A1203类、SiO2类、活性炭类和粘土类等,对于生物乙烯来讲,开发适用、高效的新型吸附剂是运用此方法分离乙烯的关键。
4.3 膜分离法
该方法利用混合气各组分在膜中渗透速率的差异对乙烯进行分离。用于分离乙烯的膜主要有平片膜和中空纤维膜,膜中金属离子有Na+、Ag+和Cu2+等,各组分与膜中离子形成络合物,进行迁移从而得到分离。目前,膜分离法在乙烯分离中还处于研究阶段,进一步的研究可根据生物合成乙烯方式与传统石油生产方式的混合气组分的差异变换膜材料,有望实现乙烯的高效分离。
4.4 膨胀机法
该法的核心是膨胀制冷技术,利用高压气体通过膨胀机在近似等熵膨胀的同时输出外功,产生比节流更大的温降,使混合气中露点较高的组分冷凝,达到分离乙烯的目的。其优点是能耗小、操作灵活、对进料要求不太严格,但是膨胀制冷技术在国内尚未得到解决。
4.5 金属络合分离法
该法采用溶于芳烃溶剂中的一种双金属盐类四氯化亚铜铝络合物,从混合气中有选择性地络合吸附乙烯组份,乙烯分子与络合物所形成较弱的键,可在缓和条件下进行汽提解吸,从而得到纯度较高的聚合级乙烯。此方法的局限性是络合物的制备难度较大,且国内目前尚无法掌握其关键技术。
4.6 联合工艺
采用联合工艺会改善分离效果以及经济性,如变压吸附法与蒸馏联合、变压吸附法与膜分离联合等,均使乙烯的选择性大大提高[37]。
5 展望
乙烯作为重要化工原料,其产量与国民生产生活息息相关。随着化石能源枯竭,传统合成成本升高,且产量已无法满足我国对乙烯的需求,因此降低生产成本、提高产量、开辟新型合成途径成为乙烯生产的重点研究方向。微生物合成乙烯具低成本、低能耗、环境友好、易控制等优势,特别是蓝细菌,可直接利用太阳能将二氧化碳合成乙烯,显示出极大的发展潜力,因此我们认为蓝藻乙烯是最具发展前景的生物乙烯生产方式之一。但受限于包括EFE的结构功能关系,以及转基因微生物乙烯合成代谢机制等在内的许多基础问题仍未清晰,生物乙烯的直接合成技术仍处于发展的萌芽期,面临产量低、EFE不稳定、未实现产业化等问题。因此,对微生物合成乙烯的研究,依然任重道远。鉴于此,笔者认为,今后微生物合成乙烯研究重点可集中于以下几个方面:第一,EFE的结构功能关系研究。DNA改组技术是揭示其结构功能关系及提高催化效率的有效手段,通过非理性分子设计结合定向筛选,可赋予EFE特殊的结构与功能。第二,代谢通路研究。通过深入分析来得到各代谢物更为精确的化学计量关系,利用合成生物学代谢途径改造手段设计出高产量通路,使乙烯高效合成成为可能。第三,工艺技术研究。通过培养系统条件优化以及发展低成本高效率的产物分离技术等,从系统集成上促进微生物合成乙烯技术体系的发展。
[1] 贾莹莹, 杜平, 杜风光, 等. 生物乙烯的发展状况及展望[J].当代化工, 2011, 40(10):1071-1072.
[2] Haro P, Ollero P, Trippe F. Technoeconomic assessment of potential processes for bio-ethylene production[J]. Fuel Processing Technology, 2013, 114(4):35-48.
[3] Hu YC, Zhan NN, Dou C, et al. Selective dehydration of bio-ethanol to ethylene catalyzed by lanthanum-phosphorous-modified HZSM-5 Influence of the fusel[J]. Biotechnology Journal, 2010, 5(11):1186-1191.
[4] 孙芝兰, 陈以峰. 乙烯的直接生物合成[J]. 生物工程学报,2013, 29(10):1431-1440.
[5] 胡燚, 李慧, 黄和, 等. 生物乙烯开发进展与产业化[J]. 现代化工, 2009, 29(1):6-9.
[6] Eckert C, Wu X, Wei X, et al. Ethylene-forming enzyme and bioethylene production[J]. Biotechnol Biofuels, 2014, 7(1):33.
[7] Fan D, Dai DJ, Wu HS. Ethylene formation by catalytic dehydration of ethanol with industrial considerations[J]. Materials, 2013, 6(1):101-115.
[8] 杜平, 贾莹莹, 孙沛勇, 等. 纤维乙醇制生物乙烯前景展望[J].广东化工, 2013, 9(40):78-79.
[9] 胡铁刚, 程可可, 张建安, 等. 生物乙醇催化制备乙烯的研究进展[J]. 现代化工, 2007(Z):96-99.
[10] Schlink K. Gene expression profiling in wounded and systemic leaves of Fagus sylvatica reveals up-regulation of ethylene and jasmonic acid signaling[J]. Plant Biology, 2011, 13(3):445-452.
[11] Li Q, Csanády GA, Kessler W, et al. Kinetics of ethylene and ethylene oxide in subcellular fractions of lungs and livers of male B6C3F1 mice and male fischer 344 rats and of human livers[J]. Toxicol Ogical Science, 2011, 123(2):384-398.
[12] Lin ZF, Zhong SL, Grierson D. Recent advances in ethylene research[J]. Journal of Experimental Botany, 2009, 60(12):3311-3336.
[13] 陈建新, 刘国顺, 陈占宽, 等. 乙烯生物合成途径及其相关基因工程的研究进展[J]. 热带亚热带植物学报, 2002, 10(1):83-98.
[14] 刘进平. 乙烯生物合成关键酶基因研究进展[J]. 热带农业科学, 2013, 33(1):51-57.
[15] 陈银华, 黄伟, 王海. ACC氧化酶基因研究进展[J]. 海南大学学报:自然科学版, 2006, 24(2):194-200.
[16] Hamilton AJ, Bouzayen M, Grierson D. Identification of a tomato gene for the ethylene-forming enzyme by expression in yeast[J]. Plant Biology, 1991, 88(8):7434-7437.
[17] Chen YF, Gao ZY, Kerris RJ, et al. Ethylene receptors function as components of high-molecular-mass protein complexes in Arabidopsis[J]. PLoS One, 2010, 5(1):e8640.
[18] Qadir A, Hewett EW, Long PG, et al. A non-ACC pathway for ethylene biosynthesis in Botrytis cinerea[J]. Postharvest Biology and Technology, 2011, 62(3):314-318.
[19] Young RE, Pratt HK, Biale JB, et al. Identification of ethylene as a volatile product of the fungus Penicillium digitatum[J]. Plant Physiology, 1951, 26(7):304-310.
[20] Biale JB. Effect of emanations from several species of fungi on respiration and color development of citrus fruits[J]. Science,1940, 91(2367):458-459.
[21] Nagahama K, Ogawa T, Fujii T, et al. Purification and properties of an ethylene-forming enzyme from Pseudomonas syringae pv. phaseolicola PK2[J]. Journal of General Microbiology, 1991, 10(137):2281-2286.
[22] Nagahama K, Yoshino K, Matsuoka M, et al. Site-directed mutagenesis of histidine residues in the ethylene-forming enzyme from Pseudomonas syringae[J]. Journal of Fermentation Bioengineering, 1998, 85(3):255-258.
[23] Sato M, Watanabe K, Yazawa M, et al. Detection of new ethyleneproducing bacteria, Pseudomonas syringae pvs. cannabina and sesami, by PCR amplification of genes for the ethylene-forming enzyme[J]. Bacteriology, 1997, 87(12):1192-1196.
[24] Zhang C, Wei ZH, Ye BC. Quantitative monitoring of 2-oxoglutarate in Escherichia coli cells by a fluorescence resonance energy transfer-based biosensor[J]. Appl Microbiol Biotechnol, 2013,97(7):8307-8316.
[25] Larsson C, Snoep JL, Norbeck J, et al. Flux balance analysis for ethylene formation in genetically engineered saccharomyces cerevisiae[J]. IET Systems Biology, 2011, 5(4):245-251.
[26] Ishihara K, Matsuoka M, Ogawa T, et al. Ethylene production using a broad-host-range plasmid in Pseudomonas syringae and Pseudomonas putida[J]. Journal of Fermentation Bioengineering,1996, 82(5):509-511.
[27] Wang Jp, Wu Lx, Xu F, et al. Metabolic engineering for ethylene production by inserting the ethylene-forming enzyme gene(efe)at the 16S rDNA sites of Pseudomonas putida KT2440[J]. Bioresource Technology, 2010, 101(3):6404-6409.
[28] Pirkov I, Albers E, Norbeck J, et al. Ethylene production by metabolic engineering of the yeast Saccharomyces cerevisiae[J]. Metabolic Engineering, 2008, 10(5):276-280.
[29] Johansson N, Quehl P, Norbeck J, et al. Identification of factors for improved ethylene production via the ethylene forming enzyme in chemostat cultures of Saccharomyces cerevisiae[J]. Microbial Cell Factories, 2013, 12(1):89.
[30] Johansson N, Persson KO, Quehl P, et al. Ethylene production in relation to nitrogen metabolism in Saccharomyces cerevisiae[J]. FEMS Yeast Research, 2014, 14(7):1110-1118.
[31] Tao L, Dong HJ, Chen X, et al. Expression of ethylene-forming enzyme(EFE)of Pseudomonas syringae pv. glycinea in Trichoderma viride[J]. Appl Microbiol Biotechnol, 2008, 80(4):573-578.
[32] Chen SF, Qin WS, Chen X, et al. Overexpression of bacterial ethylene-forming enzyme gene in Trichoderma reesei enhanced the production of ethylene[J]. International Journal of Biological Sciences, 2010, 6(1):96-106.
[33] Sakai M, Ogawa T, Matsuoka M, et al. Photosynthetic conversion of carbon dioxide to ethylene by the recombinant cyanobacterium,Synechococcus sp. PCC 7942 which harbors a gene for the ethyleneforming enzyme of Pseudomonas syringae[J]. J Ferment Bioeng,1997, 84(5):434-443.
[34] Takahama K, Matsuoka M, Nagahama K, et al. Construction and analysis of a recombinant Cyanobacterium expressing a chromosomally inserted gene for an ethylene-forming enzyme at the psbAI locus[J]. Journal of Bioscience and Bioengineering, 2003,95(3):302-305.
[35] Ungerer J, Tao L, Davis M, et al. Sustained photosynthetic conversion of CO2to ethylene in recombinant Cyanobacterium Synechocystis 6803[J]. Energy & Environmental Science, 2012,5(10):8998-9006.
[36] Zhu T, Xie XM, Li ZM, et al. Enhancing photosynthetic production of ethylene in genetically engineered Synechocystis sp. PCC 6803[J]. Green Chemistry, 2015, 17(1):421-434.
[37] 赵光辉, 李景燕, 李小军, 等. 炼厂干气中乙烯的分离技术及综合利用[J]. 化工中间体, 2008, 1(3):25-29.
(责任编辑 狄艳红)
Research Progress on Bio-ethylene
SUN Meng-ting1,2FAN Xiao-lei1GUO Rong-bo1QIU Yan-ling1ZHAO Xiao-xian1
(1. Shandong Industrial Engineering Laboratory of Biogas Production & Utilization,Qingdao Institute of Bioenergy and Bioprocess Technology,Chinese Academy of Sciences,Qingdao 266101;2. University of Chinese Academy of Sciences,Beijing 100049)
Ethylene is the chemical raw material of largest demand in the world, and is widely utilized for producing plastics, textiles and other chemical products. Currently, the most common method of producing ethylene is steam cracking of fossil oil;however, there exist drawbacks of high cost and non-friendly to environment, which bring the opportunity for the development of bio-ethylene. Bio-ethylene is produced from renewable resources of CO2and biomass , which contributes to saving resources and protecting environment. In this review,we introduce two pathways of bio-ethylene production—direct and indirect synthesis. Here we focus on the direct synthesis pathway of microorganisms via an ethylene forming enzyme(EFE), describing the structure, sequence and catalytic mechanism of EFE, illustrating several successful cases of engineered heterologous hosts synthetizing ethylene, presenting the strategies for improving yields, and prospecting the development of producing ethylene by microorganisms.
bio-ethylene;direct synthesis;microorganism;ethylene forming enzyme;heterologous expression
10.13560/j.cnki.biotech.bull.1985.2016.02.004
2015-04-20
国家“863”计划项目(2012AA052103),国家自然科学基金面上项目(41276143),国家科技支撑项目(2013BAD22B00)
孙梦婷,女,硕士,研究方向:生物化工;E-mail:374695153@qq.com
范晓蕾,女,博士,副研究员,研究方向:生物化工;E-mail:fanxl@qibebt.ac.cn
