DNA End Resection:Facts and Mechanisms
2016-09-28TingLiuJunHuang
Ting Liu*,Jun Huang*,b
1Department of Cell Biology and Program in Molecular Cell Biology,Zhejiang University School of Medicine,
Hangzhou 310058,China
2Life Sciences Institute and Innovation Center for Cell Signaling Network,Zhejiang University,Hangzhou 310058,China
REVIEW
DNA End Resection:Facts and Mechanisms
Ting Liu1,*,a,Jun Huang2,*,b
1Department of Cell Biology and Program in Molecular Cell Biology,Zhejiang University School of Medicine,
Hangzhou 310058,China
2Life Sciences Institute and Innovation Center for Cell Signaling Network,Zhejiang University,Hangzhou 310058,China
Available online 27 May 2016
Handled by Xingzhi Xu
KEYWORDS
DNA end resection;
Homologous recombination;
DNA double-strand breaks;
Chromatin remodeling
factors;
Genome stability
AbstractDNA double-strand breaks(DSBs),which arise following exposure to a number of endogenous and exogenous agents,can be repaired by either the homologous recombination(HR)or non-homologous end-joining(NHEJ)pathways in eukaryotic cells.A vital step in HR repair is DNA end resection,which generates a long 3′single-stranded DNA(ssDNA)tail that can invade the homologous DNA strand.The generation of 3′ssDNA is not only essential for HR repair,but also promotes activation of the ataxia telangiectasia and Rad3-related protein(ATR).Multiple factors,including the MRN/X complex,C-terminal-binding protein interacting protein(CtIP)/Sae2,exonuclease 1(EXO1),Bloom syndrome protein(BLM)/Sgs1,DNA2 nuclease/helicase,and several chromatin remodelers,cooperate to complete the process of end resection.Here we review the basic machinery involved in DNA end resection in eukaryotic cells.
Introduction
Double-strand breaks(DSBs)are one of the most dangerous types of DNA damage because they disrupt the continuity of chromosomes[1,2].Failure to eliminate DSBs leads to genome instability and tumorigenesis[1,3].DSBs are predominantly repaired by either the non-homologous end-joining(NHEJ)pathway or the homologous recombination(HR)pathway[4,5].NHEJ directly ligates the broken DNA ends,whereas HR uses a homologous sequence from sister chromatid as a repair template[4,6].
Using an identical or similar sequence as a template,HR is commonly considered to be an error-free mechanism for the repair of DSBs[7,8].When DSBs occur,a process termed DNA end resection is activated,which catalyzes the nucleolytic degradation of the broken ends in the 5′to the 3′direction[9,10].The resulting 3′single-stranded DNA(ssDNA)then provides a platform for the recruitment of proteins that participate in HR repair[9-11].Interestingly,DNA end resection inhibits NHEJ and triggers homology-directed DSB repair[11].Multiple proteins or protein complexes have been shown to be involved in this process.These include the MRNcomplex(MRXcomplexinbuddingyeast),C-terminal-binding protein interacting protein(CtIP;Sae2 in budding yeast),exonuclease 1(EXO1),Bloom syndrome protein(BLM;Sgs1 in budding yeast),DNA2 nuclease/helicase,and several chromatin remodeling factors[12].Here,we discuss the pivotal proteins and their mechanisms during DNA end resection.
DNA end resection and the repair pathway choice
Although DSBs can occur at any phase of the cell cycle,DNA end resection only happens in the S and G2 phases[9,13]. During other cell cycle phases,DNA end resection is inhibited by Ku70/80 heterodimers and other proteins;therefore,only the NHEJ pathway can be initiated[11].NHEJ promotes direct ligation of the DNA ends;subsequent processing of the broken DNA ends is unnecessary[11].This phenomenon is also consistent with the finding that sister chromatids only exist in the S and G2 phases.However,the repair pathway choice also depends on substrate complexity and other factors besides the cell cycle[13,14].
The MRN/X complex
The MRN complex,which comprises MRE11,RAD50,and nibrin(NBS1),plays key roles in DNA end resection and HR repair in mammalian cells[15,16].The counterpart of the MRN complex in budding yeast is the MRX complex,which consisting of Mre11,Rad50,and Xrs2[17].The MRN/X complex not only functions in DNA end resection,but also plays critical roles in the DNA damage checkpoint response[18].
The MRN complex binds DNA through its globular domain,in which MRE11 and NBS1 associate with the Walker A and Walker B motifs of RAD50[19].Previous studies suggest that the DNA binding activity requires primarily MRE11 and RAD50[19-21].The extended coiled-coil tail of RAD50 forms another structural domain in the whole MRN complex,which is important for the DNA-binding and-tethering activities of the complex[22-26].
MRE11 is the core component of the MRN complex and exhibits a variety of enzymatic activities,including 3′to 5′exonuclease activity on dsDNA,endonuclease activity on ssDNA,andDNA-annealingand-unwindingactivities[20,21,27,28].In vitro experiments revealed that the five phosphoesterase motifs within the N-terminal region of MRE11 are essential for its biochemical activities[12,29]. Paradoxically,generation of the 3′overhang requires the activity of 5′to 3′exonuclease,which is opposite to the observed exonuclease activity of MRE11[30,31].A twostep mechanism of MRE11 has thus been proposed,that is,MRE11 makes the initial ssDNA nick via its ssDNA endonuclease activity at first and then digests toward the DSB end through its 3′to 5′exonuclease activity to produce 3′ssDNA tails[31-34].
Human NBS1 contains two BRCA1 C terminus(BRCT)domains and a forkhead-associated(FHA)domain[35]. Mutations within the NBS1 gene are responsible for the Nijmegen breakage syndrome,a rare autosomal recessive disease that increases the predisposition to develop malignancies[36-38].Cells derived from NBS patients exhibit defects in DSB repair and cell cycle checkpoint[36].Although lacking enzymatic activities,NBS1 is considered to be an important regulator in the MRN complex,since NBS1 influences both DNA binding and nuclease activity of MRE11[39-41].
CtIP/Sae2
CtIP was first identified as a cofactor for the transcriptional repressor C-terminal-binding protein(CtBP)[12,42].Further studies reveal that CtIP functions in many other cellular processes,including cell cycle regulation and tumorigenesis[43].Interestingly,CtIP is now better known as an interacting partner of the MRN complex,for its involvement in DNA end resection and DSB repair[44-47].
CtIP shows sequence homology to the budding yeast Sae2 at the C terminus[48].CtIP plays at least two roles in the process of DNA end resection,distinguished by the involvement of its catalytic activity or not[49].Briefly,the resection of DSBs with clean broken ends produced by restriction enzymes is dependent on the presence of CtIP protein,but independent of its nuclease activity[49,50].By contrast,the repair of more complex DNA lesions created by topoisomerase poisons or ionizing radiation(IR)requires not only the presence of CtIP protein but also its endonuclease activity[51].
EXO1
EXO1 belongs to the xeroderma pigmentosum complementation group G(XPG)family of nucleases,which contain conserved nuclease motifs in the N-terminal region[52,53]. EXO1 exhibits 5′to 3′dsDNA exonuclease and 5′flap endonuclease activities in vitro[12,53].Interestingly,EXO1 prefers dsDNA substrates with a recessed 5′end,which is produced by the MRN/X complex and CtIP/Sae2[54-56].Taken together with the finding that MRE11 lacks the 5′to 3′exonuclease activity required to produce long 3′ssDNA tails necessary for replication protein A(RPA)binding,a two-step model has been suggested for DSB processing[12].In this model,the MRN/X complex and CtIP/Sae2 remove the first 50-100 nucleotides from the 5′end of the broken DNA,followed by the generation of long 3′ssDNA tails catalyzed by EXO1[12].This model is also supported by the finding that CtIP is required for the accumulation of EXO1 at DSB sites in vivo[57].
DNA2-BLM/Sgs1
BLM is a member of the RecQ family of helicases that unwinds DNA in mammals and Sgs1 is its ortholog in Saccharomyces cerevisiae[58].DNA2,which is related to the bacterial RecB proteins,exhibits both helicase and nuclease activities in vitro[33].However,the helicase activity of DNA2 is not necessary for DNA end resection,while the nuclease activity of DNA2 is essential to this process[59-61].Previous reports suggest that EXO1 and DNA2-BLM/Sgs1 function in parallel at the second step of end resection[12].Interestingly,studies in yeast indicate that Sgs1-Dna2-catalyzed end resection is dependent on RPA[61].In the absence of RPA,DNA2 cannot be recruited to DSBs[61].Although it can degrade either 3′-or 5′-terminated ssDNA,DNA2 exhibits 5′endonuclease activityonly in the presence of RPA,which may explain the strand bias in the end resection[61].
Chromatin remodeling factors
Eukaryotic DNA is normally wrapped around a histone octamer to form nucleosomes[62].This condensation allows the long genetic molecules to fit into the relatively-small nucleus,but at the same time,forms a barrier for resection enzymes to access[63].Certain histone modifiers,histone chaperones,and chromatin remodelers modify chromatin structure and hence regulate the dynamics of the chromatin[64,65].For instance,several chromatin remodeling factors,such as remodels the structure of chromatin(RSC),INO80,switch/sucrose non-fermentable(SWI/SNF),SWI/SNF-relatedmatrixassociated actin-dependent regulator of chromatin subfamily A containing DEAD/H box 1(SMARCAD1;Fun30 in yeast),and Snf2-related CREB-binding protein(CREBBP)activator protein(SRCAP;SWR1 in yeast),are involved in the process of overcoming barriers to allow repair proteins to access(Figure 1)[66-71].
Rad9,the yeast 53BP1 ortholog,is a checkpoint mediator proteinand isrecruitedtoDSBsitesbyγ-H2Aand Resection initiation is stimulated by SRCAP,CtIP,and the MRN complex.SMARCAD1 cooperates with EXO1 and BLM/DNA2 to promote resection extension.The figure is adapted from[70]. ATR,ataxia telangiectasia and Rad3-related protein;BLM,Bloom syndrome protein;CtIP,C-terminal-binding protein interacting protein;EXO1,exonuclease 1;HR,homologous recombination;MRN,MRE11,RAD50,and nibrin;RPA,replication proteinA;SMARCAD1,switch/sucrosenon-fermentable(SWI/SNF)-related matrix-associated actin-dependent regulator of chromatin subfamily A containing DEAD/H box 1;SRCAP,Snf2-related CREB-binding protein activator protein;ssDNA,single-stranded DNA.
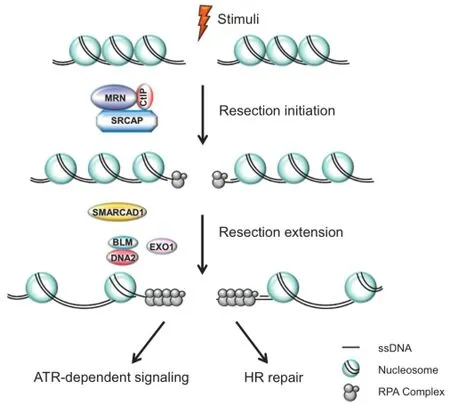
Figure 1 DNA-end resection occurs via a two-step process—resection initiation and resection extension
K79-methylated histone H3[72].Rad9 is known to inhibit DNA end resection[72].On the other hand,the yeast Fun30,which possesses intrinsic ATP-dependent chromatin remodeling activity,works together with DNA2 and EXO1 to promote extensive DSB end resection[71,73-75].Mechanically,Fun30 overcomes the barrier formed by Rad9-bound chromatin,thuspromotingextensiveresectionprocess[71,73-75].
SRCAP is a member of the INO80 ATPase family and belongs to the human SRCAP chromatin remodeling complex[76].SRCAP was first discovered as the binding partner of CREBBP(also known as CBP),and mutations in SRCAP cause a rare genetic disorder known as Floating-Harbor syndrome[76,77].Our recent findings support a new function for SRCAP in promoting DSB resection and HR repair(Figure 1).SRCAP-depleted cells exhibit RPA2 hyperphosphorylation and defects in RPA2 focus formation,indicating that SRCAP is involved in DSB end processing[78].Accordingly,SRCAP depletion only affects IR-and camptothecin(CPT)-induced RPA2 focus formation but not hydroxyurea(HU)-induced RPA2 focus formation(HU stalls replication forks by deprivation of dNTPs)[78].Moreover,zinc finger HIT-type 1(ZNHIT1)/p18,another component of the SRCAP complex,also promotes DNA end resection[78].Mechanistically,SRCAP promotes chromatin relaxation to allow CtIP accumulation at DSB sites,thereby facilitating DSB end processing(Figure 1)[78].
Summary
Correct repair of DSBs is critical for the maintenance of genome stability.HR and NHEJ are the two dominant repair pathways involved in DSB repair[4,6].While NHEJ facilitates the direct ligation of the DSB ends in an error-prone manner,HR allows for precise repair of DSBs due to the employment of homologous chromatids[8].DNA end resection is a pivotal step in HR repair to produce 3’overhangs that not only inhibit NHEJ but also provide a platform to recruit proteins involved in HR repair[11].DNA end resection is completed through a two-step process in which the MRN/X complex and CtIP/Sae2 protein are involved in the initial step,and EXO1 and DNA2-BLM/Sgs1 are involved in the second step[79].However,precisely how these resection factors are regulated in a coordinated manner is still unclear.Further studies are therefore required to resolve this issue.
Competing interests
The authors declare no competing financial interests.
Acknowledgments
This work was supported in part by the grants from the National Natural Science Foundation of China(Grant Nos. 31071243 and 31171347),the Fundamental Research Funds for the Central Universities of China,and the Research Fund for the Doctoral Program of Higher Education of China(Grant No.20110101120152).
References
[1]van Gent DC,Hoeijmakers JH,Kanaar R.Chromosomal stability and the DNA double-stranded break connection.Nat Rev Genet 2001;2:196-206.
[2]Shiloh Y,Lehmann AR.Maintaining integrity.Nat Cell Biol 2004;6:923-8.
[3]O’Driscoll M.Diseases associated with defective responses to DNA damage.Cold Spring Harb Perspect Biol 2012:4.http:// dx.doi.org/10.1101/cshperspect.a012773.
[4]Liu T,Huang J.Quality control of homologous recombination. Cell Mol Life Sci 2014;71:3779-97.
[5]San Filippo J,Sung P,Klein H.Mechanism of eukaryotic homologous recombination.Annu Rev Biochem 2008;77:229-57.
[6]Lieber MR.The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway.Annu Rev Biochem 2010;79:181-211.
[7]Betermier M,Bertrand P,Lopez BS.Is non-homologous endjoining really an inherently error-prone process?PLoS Genet 2014;10:e1004086.
[8]Sonoda E,Hochegger H,Saberi A,Taniguchi Y,Takeda S. Differential usage of non-homologous end-joining and homologous recombination in double strand break repair.DNA Repair(Amst)2006;5:1021-9.
[9]Longhese MP,Bonetti D,Manfrini N,Clerici M.Mechanisms and regulation of DNA end resection.EMBO J 2010;29:2864-74.
[10]Mimitou EP,Symington LS.DNA end resection-unraveling the tail.DNA Repair(Amst)2011;10:344-8.
[11]Huertas P.DNA resection in eukaryotes:deciding how to fix the break.Nat Struct Mol Biol 2010;17:11-6.
[12]Mimitou EP,Symington LS.DNA end resection:many nucleases make light work.DNA Repair(Amst)2009;8:983-95.
[13]Daley JM,Sung P.To Cut or Not to Cut:Discovery of a novel regulator of DNA break resection.Mol Cell 2016;61:325-6.
[14]Kakarougkas A,Jeggo PA.DNA DSB repair pathway choice:an orchestrated handover mechanism.Br J Radiol 2014;87:20130685.
[15]Ohta K,Nicolas A,Furuse M,Nabetani A,Ogawa H,Shibata T. Mutations in the MRE11,RAD50,XRS2,and MRE2 genes alter chromatin configuration at meiotic DNA double-stranded break sites in premeiotic and meiotic cells.Proc Natl Acad Sci U S A 1998;95:646-51.
[16]D’Amours D,Jackson SP.The Mre11 complex:at the crossroads of dna repair and checkpoint signalling.Nat Rev Mol Cell Biol 2002;3:317-27.
[17]Trujillo KM,Roh DH,Chen L,Van Komen S,Tomkinson A,Sung P.Yeast Xrs2 binds DNA and helps target Rad50 and Mre11 to DNA ends.J Biol Chem 2003;278:48957-64.
[18]Lukas C,Melander F,Stucki M,Falck J,Bekker-Jensen S,Goldberg M,et al.Mdc1 couples DNA double-strand break recognition by Nbs1 with its H2AX-dependent chromatin retention.EMBO J 2004;23:2674-83.
[19]Hopfner KP,Karcher A,Craig L,Woo TT,Carney JP,Tainer JA.Structural biochemistry and interaction architecture of the DNA double-strand break repair Mre11 nuclease and Rad50-ATPase.Cell 2001;105:473-85.
[20]Tsubouchi H,Ogawa H.A novel mre11 mutation impairs processing of double-strand breaks of DNA during both mitosis and meiosis.Mol Cell Biol 1998;18:260-8.
[21]Furuse M,Nagase Y,Tsubouchi H,Murakami-Murofushi K,Shibata T,Ohta K.Distinct roles of two separable in vitro activities of yeast Mre11 in mitotic and meiotic recombination. EMBO J 1998;17:6412-25.
[22]van Noort J,van Der Heijden T,de Jager M,Wyman C,Kanaar R,Dekker C.The coiled-coil of the human Rad50 DNA repair protein contains specific segments of increased flexibility.Proc Natl Acad Sci U S A 2003;100:7581-6.
[23]Lichten M.Rad50 connects by hook or by crook.Nat Struct Mol Biol 2005;12:392-3.
[24]Paull TT.New glimpses of an old machine.Cell 2001;107:563-5.
[25]de Jager M,van Noort J,van Gent DC,Dekker C,Kanaar R,Wyman C.Human Rad50/Mre11 is a flexible complex that can tether DNA ends.Mol Cell 2001;8:1129-35.
[26]Hopfner KP,Craig L,Moncalian G,Zinkel RA,Usui T,Owen BA,et al.The Rad50 zinc-hook is a structure joining Mre11 complexes in DNA recombination and repair.Nature 2002;418: 562-6.
[27]Lee JH,Paull TT.Direct activation of the ATM protein kinase by the Mre11/Rad50/Nbs1 complex.Science 2004;304:93-6.
[28]Haber JE.The many interfaces of Mre11.Cell 1998;95:583-6.
[29]Neale MJ,Pan J,Keeney S.Endonucleolytic processing of covalent protein-linked DNA double-strand breaks.Nature 2005;436:1053-7.
[30]Stracker TH,Petrini JH.The MRE11 complex:starting from the ends.Nat Rev Mol Cell Biol 2011;12:90-103.
[31]Mimitou EP,Symington LS.Sae2,Exo1 and Sgs1 collaborate in DNA double-strand break processing.Nature 2008;455:770-4.
[32]Shibata A,Moiani D,Arvai AS,Perry J,Harding SM,Genois MM,et al.DNA double-strand break repair pathway choice is directedbydistinctMRE11nucleaseactivities.MolCell 2014;53:7-18.
[33]Zhu Z,Chung WH,Shim EY,Lee SE,Ira G.Sgs1 helicase and two nucleases Dna2 and Exo1 resect DNA double-strand break ends.Cell 2008;134:981-94.
[34]Gravel S,Chapman JR,Magill C,Jackson SP.DNA helicases Sgs1 and BLM promote DNA double-strand break resection. Genes Dev 2008;22:2767-72.
[35]Stracker TH,Morales M,Couto SS,Hussein H,Petrini JH.The carboxy terminus of NBS1 is required for induction of apoptosis by the MRE11 complex.Nature 2007;447:218-21.
[36]Williams BR,Mirzoeva OK,Morgan WF,Lin J,Dunnick W,Petrini JH.A murine model of Nijmegen breakage syndrome. Curr Biol 2002;12:648-53.
[37]Zhu J,Petersen S,Tessarollo L,Nussenzweig A.Targeted disruption of the Nijmegen breakage syndrome gene NBS1 leads to early embryonic lethality in mice.Curr Biol 2001;11:105-9.
[38]Tauchi H,Kobayashi J,Morishima K,van Gent DC,Shiraishi T,Verkaik NS,et al.Nbs1 is essential for DNA repair by homologous recombination in higher vertebrate cells.Nature 2002;420:93-8.
[39]Dong Z,Zhong Q,Chen PL.The Nijmegen breakage syndrome protein is essential for Mre11 phosphorylation upon DNA damage.J Biol Chem 1999;274:19513-6.
[40]Paull TT,GellertM.Nbs1 potentiatesATP-drivenDNA unwinding and endonuclease cleavage by the Mre11/Rad50 complex.Genes Dev 1999;13:1276-88.
[41]Williams RS,Dodson GE,Limbo O,Yamada Y,Williams JS,Guenther G,et al.Nbs1 flexibly tethers Ctp1 and Mre11-Rad50 to coordinate DNA double-strand break processing and repair.Cell 2009;139:87-99.
[42]Schaeper U,Subramanian T,Lim L,Boyd JM,Chinnadurai G. Interaction between a cellular protein that binds to the C-terminal region of adenovirus E1A(CtBP)and a novel cellular protein is disrupted by E1A through a conserved PLDLS motif.J Biol Chem 1998;273:8549-52.
[43]Wong AK,Ormonde PA,Pero R,Chen Y,Lian L,Salada G,et al.Characterization of a carboxy-terminal BRCA1 interacting protein.Oncogene 1998;17:2279-85.
[44]You Z,Shi LZ,Zhu Q,Wu P,Zhang YW,Basilio A,et al.CtIP links DNA double-strand break sensing to resection.Mol Cell 2009;36:954-69.
[45]Sartori AA,Lukas C,Coates J,Mistrik M,Fu S,Bartek J,et al. Human CtIP promotes DNA end resection.Nature 2007;450: 509-14.
[46]Takeda S,Nakamura K,Taniguchi Y,Paull TT.Ctp1/CtIP and the MRN complex collaborate in the initial steps of homologous recombination.Mol Cell 2007;28:351-2.
[47]Chen L,Nievera CJ,Lee AY,Wu X.Cell cycle-dependent complex formation of BRCA1.CtIP.MRN is important for DNA double-strand break repair.J Biol Chem 2008;283:7713-20.
[48]Akamatsu Y,Murayama Y,Yamada T,Nakazaki T,Tsutsui Y,Ohta K,et al.Molecular characterization of the role of the Schizosaccharomyces pombe nip1+/ctp1+gene in DNA doublestrand break repair in association with the Mre11-Rad50-Nbs1 complex.Mol Cell Biol 2008;28:3639-51.
[49]Makharashvili N,Tubbs AT,Yang SH,Wang H,Barton O,Zhou Y,et al.Catalytic and noncatalytic roles of the CtIP endonuclease in double-strand break end resection.Mol Cell 2014;54:1022-33.
[50]Wang H,Li Y,Truong LN,Shi LZ,Hwang PY,He J,et al.CtIP maintains stability at common fragile sites and inverted repeats by endresection-independentendonucleaseactivity.MolCell 2014;54:1012-21.
[51]Makharashvili N,Paull TT.CtIP:A DNA damage response protein at the intersection of DNA metabolism.DNA Repair(Amst)2015;32:75-81.
[52]Szankasi P,Smith GR.A DNA exonuclease induced during meiosisofSchizosaccharomycespombe.JBiolChem 1992;267:3014-23.
[53]Szankasi P,Smith GR.A role for exonuclease I from S.pombe in mutationavoidanceandmismatchcorrection.Science 1995;267:1166-9.
[54]Shim EY,Chung WH,Nicolette ML,Zhang Y,Davis M,Zhu Z,et al.Saccharomyces cerevisiae Mre11/Rad50/Xrs2 and Ku proteins regulate association of Exo1 and Dna2 with DNA breaks.EMBO J 2010;29:3370-80.
[55]Krasner DS,Daley JM,Sung P,Niu H.Interplay between Ku and replication protein A in the restriction of Exo1-mediated DNA break end resection.J Biol Chem 2015;290:18806-16.
[56]Cannavo E,Cejka P,Kowalczykowski SC.Relationship of DNA degradation by Saccharomyces cerevisiae exonuclease 1 and its stimulation by RPA and Mre11-Rad50-Xrs2 to DNA end resection.Proc Natl Acad Sci U S A 2013;110:E1661-8.
[57]Eid W,Steger M,El-Shemerly M,Ferretti LP,Pena-Diaz J,Konig C,et al.DNA end resection by CtIP and exonuclease 1 prevents genomic instability.EMBO Rep 2010;11:962-8.
[58]Nimonkar AV,Ozsoy AZ,Genschel J,Modrich P,Kowalczykowski SC.Human exonuclease 1 and BLM helicase interact to resect DNA and initiate DNA repair.Proc Natl Acad Sci U S A 2008;105:16906-11.
[59]Cejka P,Cannavo E,Polaczek P,Masuda-Sasa T,Pokharel S,Campbell JL,et al.DNA end resection by Dna2-Sgs1-RPA and its stimulation by Top3-Rmi1 and Mre11-Rad50-Xrs2.Nature 2010;467:112-6.
[60]Niu H,Chung WH,Zhu Z,Kwon Y,Zhao W,Chi P,et al. Mechanism of the ATP-dependent DNA end-resection machinery from Saccharomyces cerevisiae.Nature 2010;467:108-11.
[61]Nimonkar AV,Genschel J,Kinoshita E,Polaczek P,Campbell JL,Wyman C,et al.BLM-DNA2-RPA-MRN and EXO1-BLMRPA-MRN constitute two DNA end resection machineries for human DNA break repair.Genes Dev 2011;25:350-62.
[62]Unal E,Arbel-Eden A,Sattler U,Shroff R,Lichten M,Haber JE,et al.DNA damage response pathway uses histone modification to assemble a double-strand break-specific cohesin domain.Mol Cell 2004;16:991-1002.
[63]van Attikum H,Gasser SM.The histone code at DNA breaks:a guide to repair?Nat Rev Mol Cell Biol 2005;6:757-65.
[64]Cao LL,Shen C,Zhu WG.Histone modifications in DNA damage response.Sci China Life Sci 2016;59:257-70.
[65]Rogakou EP,Pilch DR,Orr AH,Ivanova VS,Bonner WM.DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139.J Biol Chem 1998;273:5858-68.
[66]van Attikum H,Fritsch O,Hohn B,Gasser SM.Recruitment of the INO80 complex by H2A phosphorylation links ATP-dependent chromatin remodeling with DNA double-strand break repair.Cell 2004;119:777-88.
[67]Morrison AJ,Highland J,Krogan NJ,Arbel-Eden A,Greenblatt JF,Haber JE,et al.INO80 and gamma-H2AX interaction links ATP-dependent chromatin remodeling to DNA damage repair. Cell 2004;119:767-75.
[68]Shim EY,Ma JL,Oum JH,Yanez Y,Lee SE.The yeast chromatin remodeler RSC complex facilitates end joining repair of DNA double-strand breaks.Mol Cell Biol 2005;25:3934-44.
[69]Shim EY,Hong SJ,Oum JH,Yanez Y,Zhang Y,Lee SE.RSC mobilizes nucleosomes to improve accessibility of repair machinery to the damaged chromatin.Mol Cell Biol 2007;27:1602-13.
[70]Chambers AL,Downs JA.The RSC and INO80 chromatinremodeling complexes in DNA double-strand break repair.Prog Mol Biol Transl Sci 2012;110:229-61.
[71]Costelloe T,Louge R,Tomimatsu N,Mukherjee B,Martini E,Khadaroo B,et al.The yeast Fun30 and human SMARCAD1 chromatin remodellers promote DNA end resection.Nature 2012;489:581-4.
[72]Lazzaro F,Sapountzi V,Granata M,Pellicioli A,Vaze M,Haber JE,et al.Histone methyltransferase Dot1 and Rad9 inhibit singlestranded DNA accumulation at DSBs and uncapped telomeres. EMBO J 2008;27:1502-12.
[73]Chen X,Cui D,Papusha A,Zhang X,Chu CD,Tang J,et al.The Fun30 nucleosome remodeller promotes resection of DNA double-strand break ends.Nature 2012;489:576-80.
[74]Durand-Dubief M,Will WR,Petrini E,Theodorou D,Harris RR,Crawford MR,et al.SWI/SNF-like chromatin remodeling factor Fun30 supports point centromere function in S.cerevisiae.PLoS Genet 2012;8:e1002974.
[75]Eapen VV,Sugawara N,Tsabar M,Wu WH,Haber JE.The Saccharomyces cerevisiae chromatin remodeler Fun30 regulates DNA end resection and checkpoint deactivation.Mol Cell Biol 2012;32:4727-40.
[76]Johnston H,Kneer J,Chackalaparampil I,Yaciuk P,Chrivia J. Identification of a novel SNF2/SWI2 protein family member,SRCAP,which interacts with CREB-binding protein.J Biol Chem 1999;274:16370-6.
[77]Hood RL,Lines MA,Nikkel SM,Schwartzentruber J,Beaulieu C,Nowaczyk MJ,et al.Mutations in SRCAP,encoding SNF2-related CREBBP activator protein,cause Floating-Harbor syndrome.Am J Hum Genet 2012;90:308-13.
[78]Dong S,Han J,Chen H,Liu T,Huen MS,Yang Y,et al.The humanSRCAPchromatinremodelingcomplexpromotes DNA-end resection.Curr Biol 2014;24:2097-110.
[79]Bernstein KA,Rothstein R.At loose ends:resecting a doublestrand break.Cell 2009;137:807-10.
27 April 2016;revised 4 May 2016;accepted 5 May 2016
*Corresponding authors.
E-mail:liuting518@zju.edu.cn(LiuT),jhuang@zju.edu.cn(Huang J).
aORCID:0000-0002-7956-3396.
bORCID:0000-0002-7837-653X.
Peer review under responsibility of Beijing Institute of Genomics,Chinese Academy of Sciences and Genetics Society of China.
http://dx.doi.org/10.1016/j.gpb.2016.05.002
1672-0229ⓒ2016 The Authors.Production and hosting by Elsevier B.V.on behalf of Beijing Institute of Genomics,Chinese Academy of Sciences and Genetics Society of China.
This is an open access article under the CC BY license(http://creativecommons.org/licenses/by/4.0/).
杂志排行

Genomics,Proteomics & Bioinformatics的其它文章
- Functions of PARylation in DNA Damage Repair Pathways
- UFMylation:A Unique&Fashionable Modification for Life
- Maintenance of Genome Stability
- Endogenous DNA Damage and Repair Enzymes—A short summary of the scientific achievements of Tomas Lindahl,Nobel Laureate in Chemistry 2015
- DNA Damage Response in Hematopoietic Stem Cell Ageing
- Connecting Malfunctioning Glial Cells and Brain Degenerative Disorders