氧化应激中ROS对FOXO3a转录因子的调控作用研究进展
2016-09-28邢雅琪李晓霞章吉威郭会彩
李 昂,邢雅琪,李晓霞,章吉威,郭会彩
(河北医科大学卫生毒理学教研室,河北 石家庄 050017)
氧化应激中ROS对FOXO3a转录因子的调控作用研究进展
李昂,邢雅琪,李晓霞,章吉威,郭会彩
(河北医科大学卫生毒理学教研室,河北 石家庄050017)
活性氧(reactive oxygen species,ROS)介导的氧化应激参与多种细胞信号转导过程,FOXO3a转录因子是氧化应激中多个信号通路的交汇点,ROS对FOXO3a存在着复杂的调控作用。由于FOXO3a在细胞增殖、细胞周期阻滞、ROS清除和诱导细胞凋亡中发挥复杂而重要的作用,已经成为氧化应激损伤的研究热点之一。该文对氧化应激损伤中FOXO3a的活性调节机制及其靶基因进行了综述,为FOXO3a靶向调控氧化应激和临床治疗相关疾病开辟了新的思路。
氧化应激;ROS;FOXO3a;调控作用;磷酸化作用;抗氧化酶
1 前言
活性氧(reactive oxygen species,ROS)是正常细胞代谢的一种副产物,参与多种细胞信号转导过程。当ROS产生过多或清除能力下降,机体氧化还原水平失衡,导致机体发生氧化应激(oxidative stress)损伤[1]。ROS介导的氧化应激参与众多病理生理学过程,是导致组织损伤、机体衰老、疾病和细胞凋亡的一个重要因素。如何通过内源性途径有效地维持细胞内ROS稳态,恢复机体抗氧化平衡已成为许多领域的研究热点。
FOXO3a转录因子属于叉头框转录因子家族(forkhead box,FOX)的O亚族,参与细胞内包括调节细胞周期、促进凋亡、抗氧化应激损伤等在内的多种生物学调控。研究表明,FOXO3a广泛表达于成人的各种组织器官中,包括骨骼肌、心血管系统、神经系统、肾、肝、胰腺和胸腺中。基于FOXO3a维持活性氧稳态方面的重要作用及其广泛表达、内源性等特点,为抗氧化应激损伤的研究开辟了新的途径。
2 FOXO3a的结构基础
Kenyon等[2]研究秀丽线虫(C.elegans)突变体时发现,DAF-16基因在高度保守的转导通路中发挥抗氧化应激和促进长寿的作用。而DAF-16基因与哺乳动物FOXO转录因子同源,人们猜想,FOXO转录因子可能同样具有抗氧化应激和促进长寿的作用,引发了对FOXO广泛而深入的研究(Fig 1)。
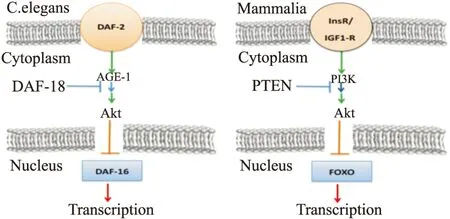
Fig 1 Insulin signaling in mammalian cells and in C. elegans
See text for further details.C.elegans DAF-2 and DAF-16 gene homologous to PI3K/Akt and FOXO respectively.DAF-2 possessing DAF-16 inhibition is similar to the PI3K/Akt suppression of FOXO
哺乳动物的FOXO转录因子家族包含4个成员:FOXO1、FOXO4、FOXO3a和FOXO6,它们都具有由110个氨基酸组成的DNA结合结构域,其中以FOXO3a的分布和功能最为广泛[3]。FOXO3a上的Thr32、Ser253、Ser 315、Ser 644残基等磷酸化位点[4],Lys242、 Lys245氨基酸残基等乙酰化位点[5],可与细胞内的多种信号分子交互作用,从而影响FOXO3a的活性与功能。
3 氧化应激对FOXO3a的调控作用
当机体发生氧化应激时,累积的ROS通过激活多种信号通路,使FOXO3a发生磷酸化/去磷酸化,乙酰化/去乙酰化等翻译后修饰,从而调控FOXO3a的活性与功能。而这一过程是相当复杂,有些甚至是矛盾的。以磷酸化为例,PI3K-AKT通路与p38 MAPK(p38 mitogen-activated kinase)、ERK(extracellular-signal-regulated kinase)、SGK(serum/glucocorticoid regulatedkinase 1)、IKKβ(IkB kinase β)等激酶可使FOXO3a发生磷酸化,抑制其转录活性;而AMPK、JNK(c-Jun N-terminal kinase)、MST1等蛋白激酶则可通过磷酸化作用,增强FOXO3a的转录活性(Fig 2)。
3.1ROS抑制FOXO3a
3.1.1FOXO3a的磷酸化与泛素化Coskun等[6]研究发现,在HCT 116和HT-29结肠癌细胞中,PI3K-AKT信号通路在Thr32、Ser253、Ser315位点使FOXO3a发生磷酸化,磷酸化的FOXO3a与14-3-3蛋白结合形成复合物,由胞核转移到胞质,减少其转录活性,抑制细胞凋亡。Dey等[7]在乳腺癌细胞的研究中发现,使用Akt的抑制剂Iturin A,处理MDA-MB-231和 MCF-7 乳腺癌细胞后,减少了FOXO3a的磷酸化,导致FOXO3a的核内聚集,增强了其转录活性。紫铆因(butein)是一种多酚类化合物。在小鼠子宫颈癌模型中,紫铆通过抑制AKT、ERK以及p38 MAPK信号通路,减少FOXO3a的磷酸化,从而激活FOXO3a入核,增强子宫颈癌细胞对铂化合物的敏感性,发挥抗肿瘤活性[8]。p66Shc蛋白在多种组织中介导氧化应激,有研究表明,激活p66Shc 造成的氧化应激反而可增加ERK-1/2、Akt表达,增强对FOXO3a磷酸化作用,抑制FOXO3a的转录活性[9]。
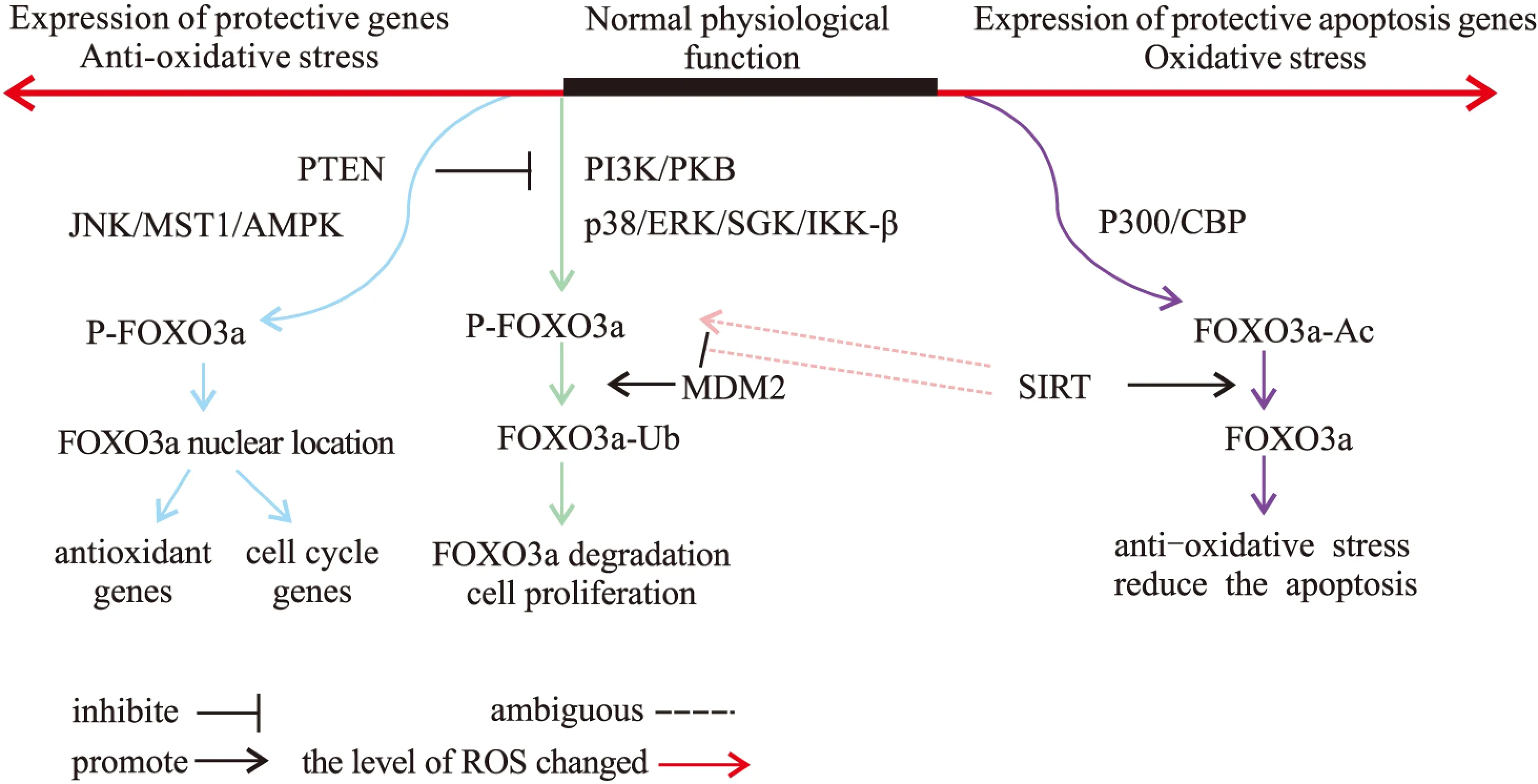
Fig 2 Activation or inhibition of FOXO3a through ROS-induced posttranslational modifications
FOXO3a蛋白分子经多种蛋白激酶的磷酸化作用后,与泛素连接酶E3 MDM2结合,作为MDM2的底物泛素化进而降解[10]。以上过程使FOXO3a与下游靶基因隔绝,导致FOXO3a的转录活性受到抑制。
3.1.2FOXO3a的乙酰化CBP(cAMP binding protein, CBP)/p300(cAMP binding protein p 300)乙酰基转移酶可在叉头区Lys242、Lys245、Lys262等位点直接乙酰化FOXO3a蛋白的赖氨酸,两者以二硫键共价结合,通过这种直接的乙酰化修饰影响FOXO3a转录能力[11]。ROS可通过介导p300/CBP乙酰化作用,促使FOXO3a半胱氨酸与p300/CBP乙酰基转移酶的硫醇形成含二硫键的复合物,从而调节FOXO3a生物学功能[12]。这种氧化还原反应对FOXO3a的直接调节,表明FOXO3a蛋白作为ROS 感受器,也是氧化应激反应的中介者,对于维持胞内氧化还原稳态具有重要作用。
3.2ROS增强FOXO3a
3.2.1FOXO3a的磷酸化与去磷酸化心脏微血管内皮细胞的研究表明,高糖刺激引起ROS积累,可通过抑制Akt激活FOXO3a,导致FOXO3a核定位,并与下游基因结合,促进细胞凋亡[13]。PTEN是PKB/Akt抑制因子,在氧化还原反应中具有重要调控作用。研究表明,在CRC(colorectal cancer)细胞中,ROS激活通过PTEN (phosphatase and tensin homologue),抑制PI3K/AKT通路,导致FOXO3a磷酸化减少,FOXO3a转录活性增强,从而诱导Bim等靶基因表达,介导细胞凋亡[14]。
AMPK、JNK(c-Jun N-terminal kinase)、MST1等蛋白激酶可通过磷酸化作用增强FOXO3a功能。ROS积累引起的氧化应激中,JNK、MST1(mammalian sterile 20-like kinase 1)和肿瘤坏死因子(tumor necrosis factor-α ,TNF-α) 等信号分子可使FOXO3a发生磷酸化,使FOXO3a发生核转位,提高其转录活性[15]。在亚砷酸导致的氧化应激中,过量的ROS激活MST1,抑制Akt对FOXO3a的磷酸化作用,促使FOXO3a核定位并进行转录[16]。
3.2.2FOXO3a的去乙酰化Sirtuins是NAD+依赖的脱乙酰酶家族,其中SIRT1、SIRT3、SIRT5均可通过去乙酰化作用激活FOXO3a,增加FOXO3a的转录活性,在抑制细胞凋亡、抗氧化应激损伤、细胞寿命延长方面发挥重要作用[17]。香烟提取物导致肺上皮细胞的应激中,SIRT5去乙酰化FOXO3a,减少细胞凋亡[18]。同样小胶质细胞中,Sirt3通过去乙酰化作用激活FOXO3a,降低细胞内ROS水平[19],SIRT3介导的FOXO3去乙酰化进一步减少FOXO3磷酸化、泛素化和降解,以此使FOXO3分子稳定[20],但也有研究指出内皮细胞中,SIRT1对FOXO3a的去乙酰化作用导致FOXO3a泛素化及降解,以此拮抗氧化应激介导的细胞凋亡[21]。
4 FOXO3a调控的靶基因
FOXO3a被激活后可诱导下游靶基因表达,发挥多种生物学作用。FOXO3a的靶基因包括抗氧化基因、细胞凋亡和细胞周期阻滞基因等。
4.1FOXO3a的靶基因:抗氧化基因正常生理状态下,细胞通过其内的解毒系统来避免内源性ROS产生过多,维持细胞内ROS的稳态,保护细胞免受损伤。ROS产生后的细胞解毒依靠抗氧化酶系统(antioxidant enzymes system)调控作用。抗氧化酶系统包括:超氧化物歧化酶(superoxide dismutases,SODs)、过氧化氢酶(catatlase)、过氧化物酶(peroxidases)等(Fig 3)。

Fig 3 FOXO3a target genes coding for antioxidant proteins
正常状态下,细胞代谢活动中产生的氧自由基能被SODs歧化为氧气(O2)和过氧化氢(H2O2)。SODs在哺乳动物体内广泛存在,包括位于线粒体基质中的MnSOD,位于胞液中的Cu-ZnSOD[22]。研究表明,暴露于过氧化氢的静止期细胞通过激活FOXO3a,直接上调MnSOD的信使RNA和蛋白表达[23]。同时,当FOXO3a的转录活性受到抑制时,MnSOD的表达减少[24]。说明MnSOD的表达受到FOXO3a的调控作用。糖尿病周围神经病变(DPN )小鼠实验中,通心络胶囊可通过抑制MAPK通路提高SOD、GSH-Px活性,有效减低小鼠的氧化应激损伤[25]。Marinkovic等[26]研究发现,FOXO3a具有调控小鼠有核红细胞胞质中的Cu-ZnSOD表达的作用,进一步证明了转录因子FOXO3a在抵抗过氧化物方面发挥着重要的作用。
机体中的多种酶系统参与到了过氧化氢的分解过程,包括过氧化氢酶、谷胱甘肽过氧化物酶(GSH-Px)、抗氧化蛋白(Prxs)等,其中一些也能被FOXO3a调控。强有力的证据证明,PI3K-Akt/PKB信号通路通过调节FOXO3a的活性来增加过氧化氢酶的表达,使过氧化氢(H2O2)分解成水(H2O)和氧气(O2)[27]。值得注意的是过氧化物酶比过氧化氢酶有更高的亲和性,在低水平的H2O2时也能产生同样的效果。谷胱甘肽过氧化物酶(GSH-Px)能催化GSH变为GSSG,使一分子H2O2还原成1分子H2O。Shalini等[28]的研究显示,在caspase-2(-/-)的小鼠肝脏细胞中,FOXO3a的活性被抑制,导致机体的谷胱甘肽过氧化物酶GSH-Px、超氧化物歧化酶SOD表达减少,ROS的清除减少,机体的抗氧化能力减弱,提示FOXO3a可通过调控GSH-Px来发挥抗氧化作用。位于过氧化物酶体、胞液和线粒体上的抗氧化蛋白(Prxs)可将过氧化氢分解成氧气和水。阿霉素(doxorubicin)预处理的H9C2心肌细胞中,ROS表达增加,对FOXO3a磷酸化减少,促进FOXO3a的定位到细胞核中,激活FOXO3a的转录活性,导致Prx Ⅲ表达增加[29]。
4.2FOXO3a的靶基因:细胞凋亡与细胞周期阻滞基因FOXO3a能增加细胞凋亡基因的表达,如Bim、FasL、TRAIL、PUMA。在结肠癌祖/干细胞中,小分子ONC201/TIC10能够抑制Akt活性。此种情况下,Akt对FOXO3a的磷酸化将减少,可激活FOXO3a的转录活性,FOXO3a定位到细胞核中,促进TRAIL的表达,从而诱导细胞凋亡[30]。同样FOXO3a还能激活细胞周期阻滞蛋白(如p21、p27)表达。p21和p27能与周期蛋白依赖性激酶(cyclin-dependent protein kinases, CDKs)结合,并抑制其活性,从而阻止细胞由G1期向S期转变。S-雌马酚可抑制cyclin B1和CDK1的表达,并通过增加FOXO3a的表达上调,增加p21和p27蛋白表达,使前列腺癌细胞发生细胞周期阻滞,同时可激活Bim和FasL基因表达,从而发挥抗肿瘤活性[31]。FOXO3a参与的细胞周期阻滞和细胞凋亡对于抑制肿瘤细胞具有重要作用,这提示FOXO3a的病理改变可能参与到细胞癌变,使其获得了不死性。
4.3FOXO3a调控下游靶基因的“长寿理论”de Keizer等[32]认为,鉴于FOXO3a作为一种“长寿基因”, FOXO3a的调控作用需要作出微调以适应不同的ROS水平。不难想象,当细胞遇到较低水平的ROS产生时,倘若FOXO3a直接调控细胞凋亡,随着时间的积累,细胞再生的资源就会消耗殆尽,这显然与长寿不符。另一方面,对于严重的ROS损伤,如果FOXO3a依然调控损伤修复,那么这些受损的细胞就可能出现突变体,引发癌症,从而限制长寿。因此,FOXO3a作为“长寿基因”发挥其功能,不在于单纯的被激活或抑制,而在于调整其能力以适应特定的细胞氧化应激环境。
5 氧化应激相关疾病与FOXO3a
在心血管系统中,内皮细胞H2O2积聚导致内皮细胞功能紊乱和动脉粥样硬化。通过激活SIRT1-FOXO3a/PGC-1α信号通路,使锰超氧化物歧化酶(MnSOD)、过氧化氢酶、抗氧化蛋白3和5(Prx3,Prx5)、硫氧还蛋白2(TRX2)、硫氧还蛋白还原酶2(TR2)及解偶联蛋白2(UCP-2)等抗氧化基因表达增加。敲除FOXO3a基因后,即使过表达SIRT1也不能使抗氧化酶表达增加[33]。心脏瓣膜手术后再狭窄主要与抑制心脏微血管内皮细胞(cardiac microvascular endothelial cell,CMECs)增殖和迁移有关,雷帕霉素治疗24 h抑制CMECs的增殖、迁移和一氧化氮(NO)的分泌,同时促进细胞凋亡和活性氧(ROS)的产生。他汀类药物可通过Akt/mTOR(mammalian target of rapamycin)/p70S6K(p70S6 Kinase)/FOXO3a信号通路,降低ROS产生,抑制细胞凋亡,促进增殖和迁移,改善内皮功能,促进血管生成过程[34]。研究发现[35],在衰老的CMECs中过表达FOXO3a,使过氧化氢酶和SOD表达增加,可显著抑制ROS的生成和p27基因的激活,但是这一过程并不能逆转衰老表型。
越来越多的证据表明,氧化应激在骨质疏松的发生发展中起到举足轻重的作用[36]。当骨组织发生氧化应激后,JNK和Mst1结合FOXO3a使其发生磷酸化,促进FOXO3a的核定位。Mst1/FOXO3a可直接诱导细胞凋亡,而JNK可通过磷酸化Mst1增强其介导的前凋亡信号,导致骨质疏松[37]。与之激活方式不同,在Bcl2(-/-)的小鼠体内,Akt的活性受到抑制,导致对FOXO3a的磷酸化减少,激活成骨细胞p53基因表达增加,介导骨质疏松的发生[38]。长期使用糖皮质激素,可诱导机体产生氧化应激,同时诱发骨质疏松[39]。Moriishi等[40]研究发现, FOXO3a既参加了糖皮质激素抑制成骨细胞分化诱发骨质疏松的过程,又能通过细胞凋亡通路增强成骨细胞分化,这两者之间似乎是矛盾的。与以往认为FOXO3a在骨质疏松中诱导细胞凋亡不同,Kim等[41]研究发现,通过激活SIRT1转录因子可使FOXO3a发生去乙酰化,使其下游抗氧化基因表达(如氧化氢酶和HO-1),同时可减少破骨细胞生成和增殖,从而抑制骨质疏松。这提示我们,FOXO3a在骨质疏松中的作用可能是复杂的,有的甚至是矛盾的。
阿尔兹海默症的发生发展也与氧化应激有关。研究发现[42],起初Cdk5(cyclin-dependent kinase 5)可直接磷酸化FOXO3a,促进FOXO3a的核定位,诱导抗氧化酶MnSOD的表达,保护细胞免受氧化应激损伤。进一步暴露于ROS后,FOXO3a则通过上调Bim和FasL基因表达,诱导细胞凋亡。提示在阿尔兹海默症中,FOXO3a介导的抗氧化调控是细胞早期暴露于ROS的结果,当机体的氧化还原反应失调后,FOXO3a会引发凋亡基因的表达。
6 问题与展望
明确调控FOXO3a转录因子信号转导通路的组成、作用及其相互关系,无疑是揭示FOXO3a氧化还原调控本质并进行精确干预的重要突破口。FOXO3a作为一种重要的核转录因子,是氧化还原调控多个信号通路的交汇点,在何种内环境、何种机体状态下,FOXO3a发挥何种调控作用尚未得到明确回答。通过深入研究不同病理生理状态下FOXO3a的调控作用及其作用机制,加大基于此通路药物的研发,将会为临床有效治疗机体氧化应激损伤提供新的途径。
[2]Kenyon C. The first long-lived mutants: discovery of the insulin/IGF-1pathway for ageing[J].PhilosTransRSocLondBBiolSci,2011,366(1561): 9-16.
[3]Zheng F, Wu J, Zhao S,et al.Baicalein increases the expression and reciprocal interplay of RUNX3 and FOXO3a through crosstalk of AMPKα and MEK/ERK1/2 signaling pathways in human non-small cell lung cancer cells[J].JExpClinCancerRes,2015,34(1): 41-54.
[4]Simon Taylor, Lam M, Pararasa C,et al.Evaluating the evidence for targeting FOXO3a in breast cancer: a systematic review[J].CancerCellInt,2015,15(1):1-9.
[5]Consolaro F, Ghaem-Maghami S, Bortolozzi R,et al.FOXO3a and posttranslational modifications mediate glucocorticoid sensitivity in B-ALL[J].MolCancerRes,2015,13(12):1578-90.
[6]Coskun D, Obakan P, Arisan E D,et al. Epibrassinolide alters PI3K/MAPK signaling axis via activating FOXO3a-induced mitochondria-mediated apoptosis in colon cancer cells[J].ExpCellRes,2015,338(1):10-21.
[7]Dey G, Bharti R, Dhanarajan G,et al.MarinelipopeptideIturin A inhibits Akt mediated GSK3β and FOXO3a signaling and triggers apoptosis in breast cancer[J].SciRep,2015,5:10316.
[8]Zhang L, Yang X, Li X,et al.Butein sensitizes HeLa cells to cisplatin through the AKT and ERK/p38 MAPK pathways by targeting FOXO3a[J].IntJMolMed,2015 ,36(4):957-66.
[9]Perrini S, Tortosa F, Natalicchio A,et al.The p66Shc protein controls redox signaling and oxidation-dependent DNA damage in human liver cells[J].AmJPhysiolGastrointestLiverPhysiol,2015 ,309(10):G826-40.
[10]Yang Y C, Tang Y A, Shieh J M, et al.DNMT3B overexpression by deregulation of FOXO3a-mediated transcription repression and MDM2 overexpression in lung cancer[J].JThoracOncol,2014,9(9):1305-15.
[11]Hagenbuchner J, Ausserlechner M J. Mitochondria and FOXO3: breath or die[J].FrontPhysiol,2013,4:147.
[12]Dansen T B, Smits L M, van Triest M H,et al.Redox-sensitive cysteines bridge p300/CBP-mediated acetylation and FoxO4 activity[J].NatChemBiol,2009,5(9):664-72.
[13]Peng C, Ma J, Gao X,et al.High glucose induced oxidative stress and apoptosis in cardiac microvascular endothelial cells are regulated by FOXO3a[J].PLoSOne,2013,8(11):e79739.
[14]Luo H, Yang Y, Duan J, et al.PTEN-regulated AKT/FOXO3a/Bim signaling contributes to reactive oxygen species-mediated apoptosis in selenite-treated colorectal cancer cells[J].CellDeathDis,2013,4:e481.
[15]Wilk A, Urbanska K, Yang S,et al.Factor 3a counteracts high glucose/tumor necrosis factor-α-mediated neuronal damage: implications for human immunodeficiency virus encephalitis[J].JNeurosciRes,2011,89(2):183-98.
[16]Yamaguchi Y,Madhyastha H, Madhyastha R,et al.Arsenic acid inhibits proliferation of skin fibroblasts, and increases cellular senescence through ROS mediated MST1-FOXO signaling pathway[J].JToxicolSci,2016,41(1):105-13.
[17]Zhang F, Li Z L, Xu X M,et al.Protective effects of icariin-mediated SIRT1/FOXO3 signaling pathway on intestinal ischemia/reperfusion-induced acute lung injury[J].MolMedRep,2015,1852(11):2442-55.
[18]Wang Y, Zhu Y, Xing S,et al.SIRT5 prevents cigarette smoke extract-induced apoptosis in lung epithelial cells via deacetylation of FOXO3[J].CellStressChaperones,2015,20(5):805-10.
[19]Rangarajan P, Karthikeyan A, Lu J,et al.Sirtuin 3 regulates FOXO3a-mediated antioxidant pathway in microglia[J].Neuroscience,2015,311:398-414.
[20]Tseng A H, Wu L H, Shieh S S,et al. SIRT3 interactions with FOXO3 acetylation, phosphorylation and ubiquitinylation mediate endothelial cell responses to hypoxia[J].BiochemJ,2014,464(1):157-68.
[21]Sionov R V, Vlahopoulos S A, Granot Z.Regulation of bim in health and disease[J].Oncotarget,2015,6(27):23058-134.
[22]Holzmeister C,Gaupels F,Geerlof A,et al.Differentialinhibition of Arabidopsis superoxidedismutases by peroxynitrite-mediated tyrosine nitration[J].JExpBot,2015, 66(3):989-99.
[23]Kops G J, Dansen T B, Polderman P E,et al.Forkhead transcription factor FOXO3a protects quiescent cells from oxidative stress[J].Nature,2002,419(6904):316-21.
[24]Callaway D A,Riquelme M A,Sharma R,et al.Caspase-2 modulates osteoclastogenesis through down-regulating oxidative stress[J].Bone,2015,76:40-8.
[25]王超,张会欣,邢邯英,等.通心络胶囊抑制 p38 MAPK 磷酸化抑制糖尿病周围神经病变小鼠氧化应激[J].中国药理学通报,2015, 31(5):726-30.
[25]Wang C,Zhang H X,Xing G Y.Tongxinluo capsule inhibits oxidative stress in diabetic peripheral neuropathy mice by inhibiting the activity of p-p38 MAPK[J].ChinPharmacolBull,2015,31(5):726-30.
[26]Marinkovic D,Zhang X,Yalcin S,et al.Foxo3 is required for the regulation of oxidative stress in erythropoiesis[J].JClinInvest,2007,117(8):2133-44.
[27]Glorieux C,Zamocky M,Sandoval J M,et al.Regulation of catalase expression in healthy and cancerous cells[J].FreeRadicBiolMed,2015,87:84-97.
[28]Shalini S,Dorstyn L,Wilson C, et al. Impaired antioxidant defence and accumulation of oxidative stress in caspase-2-deficient mice[J].CellDeathDiffer,2012,19(8):1370-80.
[29]Liu M H, Zhang Y, He J, et al. Upregulation of peroxiredoxin Ⅲ in doxorubicin-induced cytotoxicity and the FOXO3a-dependent expression in H9c2 cardiac cells[J].ExpTherMed,2015,10(4):1515-20.
[30]Prabhu V V,Allen J E,Dicker D T, et al. Small-molecule ONC201/TIC10 targets chemotherapy-resistant colorectal cancer stem-like cells in an Akt/FOXO3a/TRAIL-dependent manner[J].CancerRes,2015,75(7):1423-32.
[31]Lu Z,Zhou R,Kong Y, et al.S-equol, a secondary metabolite of natural anticancer isoflavone daidzein, inhibits prostate cancer growthinvitroandinvivo, though activating the Akt/FOXO3a pathway[J].CurrCancerDrugTargets,2015,Epub ahead of print.
[32]de Keizer P L,Burgering B M,Dansen T B.Forkhead box o as a sensor,mediator, and regulator of redox signaling[J].AntioxidRedoxSignal,2011,14(6):1093-106.
[33]Olmos Y,Sánchez-Gómez F J,Wild B, et al. SirT1 regulation of antioxidant genes is dependent on the formation of a FOXO3a/PGC-1α complex[J].AntioxidRedoxSignal,2013,19(13):1507-21.
[34]Pan Q,Xie X,Guo Y, et al.Simvastatin promotes cardiac microvascular endothelial cells proliferation,migration and survival byphosphorylation of p70 S6K and FOXO3a[J].CellBiolInt,2014,38(5):599-609.
[35]Qi X F,Chen Z Y,Xia J B, et al.FOXO3a suppresses the senescence of cardiac microvascular endothelial cells by regulating the ROS-mediated cell cycle[J].JMolCellCardiol,2015,81:114-26.
[36]Johnson S A,Feresin R G,Soung D Y, et al.Vitamin E suppressesexvivoosteoclastogenesis in ovariectomized rats[J].FoodFunct,2016,7(3):1628-33.
[37]Almeida M.Unraveling the role of FoxOs in bone-insights from mouse models[J].Bone,2011,49(3):319-27.
[38]Moriishi T,Kawai Y,Komori H,et al.Bcl2 deficiency activates FoxO through Akt inactivation and accelerates osteoblast differentiation[J].PLoSOne,2014,9(1):e86629..
[39]Frenkel B,White W,Tuckermann J. Glucocorticoid-induced osteoporosis[J].AdvExpMedBiol,2015,872:179-215.
[40]Moriishi T,Komori T. Glucocorticoid and Bone. The inhibition of osteoblast differentiation and induction of osteocyte apoptosis through the regulation of Bcl-2 by glucocorticoids[J].ClinCalcium,2014,24(9):1329-36.
[41]Kim H N, Han L,Iyer S, et al.Sirtuin1 suppresses osteoclastogenesis by deacetylatingFoxOs[J].MolEndocrinol,2015,29(10):1498-509.
[42]Shi C,Viccaro K,Lee H G, et al.Cdk5-FOXO3a axis: initially neuroprotective, eventually neurodegenerative in Alzheimer’s disease models[J].JCellSci,2016,pii: jcs.185009.[Epub ahead of print]
Redox regulation of FOXO3a transcription factor
LI Ang,XING Ya-qi,LI Xiao-xia,ZHANG Ji-wei,GUO Hui-cai
(DeptofToxicology,HebeiMedicalUniversity,Shijiazhuang050017,China)
ROS-mediated oxidative stress involved in a variety of cellular signal transduction,FOXO3a transcription factor is an intersection in regulating a variety of cellular oxidative stress.FoxO3a has been extensively studied in regulating oxidative stress because of its rather complex and pivotal regulation of cell proliferation, cell cycle arrest, ROS scavenging and apoptosis.This review will elucidate the FOXO3a’s regulatory mechanisms and describe the target genes involved.It will also provide the clinical significance and strategies to target FOXO3a to regulate oxidative stress.
oxidative stress;ROS;FOXO3a;regulation;phosphorylation;antioxidant
2016-05-30,
2016-06-27
国家自然科学基金资助项目(No 81473292);2016年河北省“大学生创新性实验计划”项目(No USIP2016020)
李昂(1993-),男,本科生,研究方向:心血管药物毒理学,E-mail:anglimail@126.com;
郭会彩(1976-),女,博士,副教授,硕士生导师,研究方向:心血管药物毒理学,通讯作者,E-mail:huicaiguo@hotmail.com
10.3969/j.issn.1001-1978.2016.09.005
A
1001-1978(2016)09-1203-05
R-05;R329.2;R349.1;R394.2;R977.3;R977.6
网络出版时间:2016-8-23 14:29:00网络出版地址:http://www.cnki.net/kcms/detail/34.1086.R.20160823.1429.010.html
