红鳍东方鲀生长性状的遗传力估计及性状间相关关系的初步研究
2016-09-26于海龙顾晨雷孔德荣闻涛崔军王秀利姜志强仇雪梅刘圣聪李荣张涛
于海龙,顾晨雷,孔德荣,闻涛,崔军,王秀利,姜志强,仇雪梅,刘圣聪,李荣,张涛
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023;2.大连天正实业有限公司,辽宁大连116011;3.大连富谷水产有限公司,辽宁大连116400)
红鳍东方鲀生长性状的遗传力估计及性状间相关关系的初步研究
于海龙1,顾晨雷1,孔德荣1,闻涛1,崔军1,王秀利1,姜志强1,仇雪梅1,刘圣聪2,李荣3,张涛2
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023;2.大连天正实业有限公司,辽宁大连116011;3.大连富谷水产有限公司,辽宁大连116400)
为估计红鳍东方鲀Takifugu rubripes生长性状的遗传力及生长性状间的相关关系,构建了5个全同胞家系,测定了210日龄 (504尾)和360日龄 (500尾)红鳍东方鲀的体质量、体长和体全长等生长性状的表型值,构建固定模型,用全同胞法估计了210日龄和360日龄红鳍东方鲀的体质量、体长和体全长3个生长性状的遗传力。结果表明:红鳍东方鲀的体质量、体长和体全长3个生长性状的遗传力估计值在210日龄时分别为0.54、0.38、0.52,在360日龄时分别为0.46、0.37、0.44,这3个性状的遗传力属于中等和高等遗传力;体质量与体长,体质量与体全长,以及体长与体全长的遗传相关系数在210日龄时分别为0.99、0.99、0.98,在360日龄时分别为0.97、0.97、0.96,这3个性状之间的遗传相关呈高度正相关;测定了102日龄 (296尾)红鳍东方鲀的体质量、体长和体高3个生长性状,利用回归模型建立了体质量 (W)与体长 (L)和体全高 (H)的数学模型为W=e-8.513L2.046H0.742(R2=0.903)。本研究结果可为红鳍东方鲀的遗传育种研究提供参考。
红鳍东方鲀;生长性状;遗传力;遗传相关;回归分析
红鳍东方鲀Takifugu rubripes是中国北方重要的海水养殖鱼类,尽管在其育苗、病害防治、饲料与营养、土池养殖、工厂化循环水养殖、离岸网箱养殖、越冬技术、苗种及商品鱼的长途运输、与对虾的混养技术、标志放流等方面做了大量的工作并取得了一定的进展,但对红鳍东方鲀生长性状相关研究较少。仅见有学者进行了氨氮、温度、形态性状对生长、体质量等影响的分析和表型相关分析[1-6]。此外,万玉美等[7]通过对21个微卫星座位与红鳍东方鲀体长、体质量、体高3个经济性状进行最小二乘方差分析,得到了6个与经济性状显著相关的微卫星标记,即f161、f273与体质量显著相关 (P<0.05),f170、f501与体长显著相关 (P<0.05),f53、f331与体高显著相关 (P<0.05)。刘秀云等[8]利用微卫星标记方法分别对3个家系红鳍东方鲀的基因型进行鉴定,对生长性状进行QTL分析。结果发现,有4个标记的4种基因型与红鳍东方鲀的体长、体质量表型值呈正相关,4个标记的4种基因型与红鳍东方鲀的体长、体质量表型值呈负相关。张丽等[9]对红鳍东方鲀的黑素皮质素受体-4基因 (Melanocortin-4receptor,MC4R)进行了单核苷酸多态性 (Single nucleotide polymorphisms,SNPs)筛选及SNPs与生长性状的相关性分析。结果发现,在 MC4R基因编码区48 nt和264 nt均发生了碱基转换突变 (G→A),但在所研究的红鳍东方鲀群体中,这两个SNPs与生长性状的相关性不强。Wang等[10]克隆了红鳍东方鲀肌肉生长抑制素 (Myostatin,MSTN)基因,发现该基因存在2个SNPs:一个是在由引物M4扩增的片段上,第748个碱基突变G→A,处于第二个外显子中,导致第166个氨基酸发生改变,由GCC(丙氨酸)突变为ACC(苏氨酸),产生的3个基因型为野生型GG、突变型AA和杂合型GA;另一个是在由引物M6扩增的片段上,第1197个碱基突变C→T,发生在第二个内含子上,产生的3个基因型为野生型CC、突变型TT和杂合型CT。方差分析表明,M6的SNP与体长、体高、体质量生长性状显著相关 (P<0.05)。国外对红鳍东方鲀的研究主要集中在基因组测序、性别决定与分化、河豚毒素、部分基因克隆与表达等方面,并取得了一定的成果。
数量性状一般是由微效多基因控制的,其表型值既受遗传效应的影响,也受环境效应的影响,甚至还受遗传与环境相互作用的影响。数量性状的遗传力是动物遗传和育种中最重要的遗传参数。在鱼类的遗传和育种中,生长性状是最重要的数量性状和经济性状。刘永新等[11]对200日龄红鳍东方鲀体质量和体长的遗传力进行了估计,分别为0.16 和0.14。此外,未见国内外其他学者对红鳍东方鲀生长性状进行遗传力估计的文献报道。本研究中,通过构建全同胞家系对210日龄和360日龄红鳍东方鲀体质量、体长、体全长的遗传力进行了估计,并对102日龄红鳍东方鲀早期生长的体质量与体长、体高的相关关系进行了拟合,旨在为进一步丰富红鳍东方鲀不同生长阶段 (或不同日龄)时生长性状的遗传力估计值,为红鳍东方鲀的群体遗传和新品种培育提供理论基础。
1 材料与方法
1.1材料
试验用红鳍东方鲀包括两个群体类型:群体Ⅰ包括5个全同胞家系,在大连天正实业有限公司河北唐山曹妃甸养殖场成功构建了红鳍东方鲀5个全同胞家系,每个家系约3万尾鱼苗,5个家系的养殖、管理等条件基本一致;群体Ⅱ为大连富谷水产有限公司养殖的红鳍东方鲀。
1.2方法
1.2.1生长性状的测定 在群体Ⅰ的5个全同胞家系内,当红鳍东方鲀鱼苗分别达到210日龄和360日龄时,从每一家系中随机选取约100尾红鳍东方鲀,测定其体质量、体长和体全长等3个生长性状。在群体Ⅱ中,随机测定同一群体中102日龄的296尾红鳍东方鲀的体质量、体长和体高等3个生长性状。
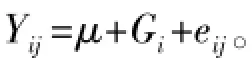
1.2.2遗传力估计 构建固定模型:其中:Yij为生长性状的表型值 (测定值);μ为性状的群体平均值;Gi为遗传效应 (家系效应);eij为随机误差效应。采用全同胞法[11-12]估计生长性状的遗传力 (h2),估计公式为
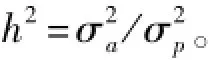
其中:σ2a为遗传方差;σ2p为表型方差。
遗传相关系数的计算公式为
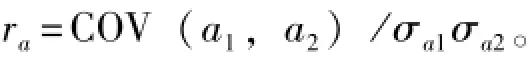
其中:ra为遗传相关系数;COV(a1,a2)为任何两个生长性状之间的遗传协方差;σa1和σa2分别为两个性状的加性遗传标准差。
1.2.3生长性状间相关关系的拟合 根据华元渝等[13]关于鱼类体质量与身体维数之间的关系式W= aLb1Hb2(W为体质量,L为体长,H为体高),利用SPSS 19.0软件进行回归分析,拟合体质量与体长、体高的数学函数。根据对数函数的性质,对关系式两边取自然对数进行变形,得
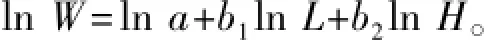
令ln W=y,ln a=A,ln L=x1,ln H=x2,可得y= A+b1x1+b2x2,进行线性回归分析,获得性状间相关关系的数学函数。
2 结果与分析
2.1红鳍东方鲀生长性状的表型值及其统计量
不同群体不同日龄的红鳍东方鲀生长性状的表型值及其统计量如表1所示。从表1可见:随着日龄的增加,群体Ⅰ的体质量、体长和体全长等生长性状的变异系数呈下降趋势;群体Ⅱ在102日龄时体质量、体长的变异系数大于群体Ⅰ在210日龄和360日龄时体质量、体长的变异系数。本研究结果表明,红鳍东方鲀日龄越小其生长性状的变异越大,即随着红鳍东方鲀日龄的增加,其生长性状变异越小。
2.2生长性状的遗传力和遗传相关估计值
以群体Ⅰ中5个全同胞生长性状的2个时间点的测定值为材料,对红鳍东方鲀体质量、体长和体全长等性状进行方差剖分和遗传力估计,结果如表2所示。从表2可见,红鳍东方鲀体质量、体长和体全长3个生长性状的遗传力在210日龄时分别为0.54、0.38、0.52,在 360日龄时分别为 0.46、0.37、0.44,除体长的遗传力属于中等遗传力外,体质量和体全长的遗传力均属于高遗传力。

表1 红鳍东方鲀生长性状的表型值及其统计量Tab.1 Phenotypic values and statistics of growth traits in redfin puffer Takifugu rubripes

表2 红鳍东方鲀生长性状的遗传力估计值Tab.2 Estimation of heritability of growth traits in redfin puffer Takifugu rubripes
2.3 生长性状的表型相关和遗传相关
以群体Ⅰ中5个全同胞生长性状的2个时间点的测定值为材料,对红鳍东方鲀体质量、体长和体全长等性状进行表型相关和遗传相关分析,结果如表3所示。从表3可见,在210日龄和360日龄时,红鳍东方鲀的体质量、体长和体全长3个生长性状间,无论是表型相关还是遗传相关都呈高度正相关。

表3 红鳍东方鲀生长性状的表型相关和遗传相关Tab.3 Phenotypic correlation and genetic correlation of growth traits in redfin puffer Takifugu rubripes
2.4生长性状间的相关关系
由群体Ⅱ所测得的红鳍东方鲀各生长性状值,以体长、体高为自变量,体质量为因变量,结合变形后的关系式,先计算出变形后的数值,然后进行二元线性回归分析,拟合出未知系数A=-8.513,b1=2.046,b2=0.742,决定系数R2=0.903,其关系式为

还原后得出红鳍东方鲀体质量与体长、体高的相关关系式为

3 讨论
3.1红鳍东方鲀遗传力估计
尽管动物分子遗传学取得了令人瞩目的成就,但分子标记辅助育种在海水养殖动物遗传育种上的应用非常成功的案例不多,常规的育种技术在鱼类遗传育种和新品种 (或家系)培育过程中仍然占有重要的地位。鱼类的数量性状大多是经济性状,因此,在鱼类的常规育种中,数量性状遗传力的估计及性状间的相关关系显得尤为重要。
遗传力是数量性状三大遗传参数中最重要的参数之一,遗传力高的性状适合个体选择和家系选择,遗传力低的性状适合家系内选择。本研究中估计的红鳍东方鲀在210日龄和360日龄时体质量、体长和体全长3个生长性状的遗传力为中等遗传力和高遗传力,其遗传力范围分别为0.46~0.54、0.37~0.38、0.44~0.52,而刘永新等[11]估计红鳍东方鲀在200日龄时体质量和体长的遗传力分别为0.16和0.14,属于低遗传力。遗传力估计值的差异如此之大,可能是因采用的估计方法不完全一样和不同群体的样本资料不同造成的。马爱军等[14]采用了4种模型对大菱鲆不同生长阶段的体质量进行了遗传力估计。结果表明,3月龄体质量的遗传力分别为0.52、0.37、0.40和0.25,9月龄体质量的遗传力分别为 0.76、0.32、0.68和0.31,15月龄体质量的遗传力分别为0.88、0.49、0.73和0.34,说明用不同模型估计的遗传力差异很大。另外,因数量性状的表型值一般是由微效多基因控制的,除受遗传因素影响外,还受环境、管理、场年季等因素的影响。本研究中所用的红鳍东方鲀群体与刘永新等[11]所用的群体不同,这可能也是产生遗传力估计值有较大偏差的原因。通常,水生生物的外部形态性状和体质量等数量性状的遗传力大多属于中等偏上的遗传力,如大黄鱼Larimichthys crocea 180日龄体质量和体全长的遗传力估计值分别为0.40、0.31[15],虹鳟360日龄体质量和体长的遗传力估计值分别为0.45和0.28[16],大口黑鲈Micropterus salmoides 4月龄和6月龄体质量遗传力分别为0.29和0.28(体长遗传力分别为0.31、0.26[17]),牙鲆24日龄体质量和体长的遗传力估计值分别为0.27和0.30[18],大西洋鳕200日龄体质量遗传力的估计值为0.29[19],银大马哈鱼130日龄体质量的遗传力估计值为0.26[20],虾夷马粪海胆3月龄和5月龄体质量和壳径的遗传力估计值范围分别为 0.399~0.523和 0.316~0.483[21],凡纳对虾 P.vannamei和红额角对虾P.stylirostris生长率的遗传力估计值分别为0.2、0.7[22],马氏珠母贝壳长、壳高和壳宽的广义遗传力分别为0.563、0.587、0.429[23]。但也有较低遗传力的,如Su等[24]对3个家系的虹鳟体质量遗传力进行估计,遗传力估计值范围为0.01~0.17。因受红鳍东方鲀群体、样本所限,本研究中并未估计红鳍东方鲀主要生长阶段 (如3月龄、6月龄、12月龄、24月龄)的体质量、体长和体全长等生长性状的遗传力。作者认为,本研究中所得到的210日龄和360日龄时红鳍东方鲀体质量、体长和体全长3个生长性状的遗传力估计值可能更接近其该日龄时的真实遗传力值。今后有必要构建并获得红鳍东方鲀的后裔资料、全同胞资料、半同胞资料,利用多种模型,对各生长阶段的生长性状进行全面的遗传力估计,为红鳍东方鲀新品种培育奠定基础。
3.2红鳍东方鲀的遗传相关和表型相关
体长、体高和体质量等生长性状的表型值是鱼类重要的生物学特征。生物作为一个有机整体,它所表现的各种性状之间必然存在着内在的联系,即性状之间存在着一定的相关关系。性状间的遗传相关对于间接选择、多性状选择和比较不同环境下的选择效果具有重要意义。本研究表明,在210日龄和360日龄时,红鳍东方鲀的体质量、体长和体全长3个生长性状间,无论是表型相关还是遗传相关都呈高度正相关,这与其他鱼类体质量、体长、体全长和体高等性状间的高度正相关一致[15-17,25]。体质量是鱼类生长性状中较难测准的性状[25],在实际育种工作中,如果对于多性状进行选择较繁杂的话,因体质量、体长和体高的高度相关,可以针对其中的一个生长性状进行选择,从而间接提高了高度相关的其他性状。另外,如果测得红鳍东方鲀体长、体高等性状,可以根据本研究中拟合的性状间关系式计算出其对应的体质量。
[1] 姜志强,何旭颖,王冰,等.不同家系红鳍东方鲀形态差异研究[J].现代农业科技,2012(17):256-257.
[2] 王新安,马爱军,庄志猛,等.红鳍东方鲀(Takifugu rubripes)形态性状对体重的影响效果[J].海洋与湖沼,2013,44(1):135-140.
[3] 王鑫,李荣,刘东超,等.氨氮对红鳍东方鲀生长及能量收支的影响[J].水产科学,2014,33(7):433-437.
[4] 刘永新,周勤,张红涛,等.温度对红鳍东方鲀早期生长和性别分化的影响[J].南方水产科学,2014,10(5):24-29.
[5] 张云,齐自元,宁丽军,等.温度对红鳍东方鲀幼鱼生长的影响[J].现代渔业信息,2010,25(12):29-34.
[6] 王新安,马爱军,庄志猛,等.红鳍东方鲀体型性状选育指标的综合判定[J].渔业科学进展,2012,33(6):10-15.
[7] 万玉美,王蕾,谭照君,等.红鳍东方鲀两个群体的遗传结构及与经济性状的相关性分析[J].淡水渔业,2011,41(5):9-16.
[8] 刘秀云,康大海,孙效文,等.红鳍东方鲀F1代混养群体的基因型鉴定及QTL分析[J].水产科学,2008,27(12):641-644.
[9] 张丽,仇雪梅,王娟,等.红鳍东方鲀(Takifugu rubripes)MC4R基因的多态性分析[J].生物技术通报,2012(7):97-102.
[10] Wang Ying,Wang Xiuli,Meng Xiangying,et al.Identification of two SNPs in myostatin(MSTN)gene of Takifugu rubripes and their association with growth traits[J].Molecular and Cellular Probes,2014,28(4):200-203.
[11] 刘永新,周勤,张红涛,等.红鳍东方鲀(Takifugu rubripes)生长性状的遗传参数估计[J].渔业科学进展,2014,35(6):39-44.
[12] 高会晓,崔妍,杜杰,等.牙鲆体质量和形态性状异速生长的遗传分析[J].大连海洋大学学报,2015,30(4):369-373.
[13] 华元渝,阮景荣.鱼类的重量-身体维数关系的研究[J].水生生物学集刊,1983,8(1):45-61.
[14] 马爱军,王新安,雷霁霖.大菱鲆(Scophthalmus maximus)不同生长阶段体重的遗传参数和育种值估计[J].海洋与湖沼,2009,40(2):187-194.
[15] 韦信键,刘贤德,王志勇.32个大黄鱼家系早期阶段生长性状比较及遗传参数估计[J].集美大学学报:自然科学版,2013,18(5):321-328.
[16] 刘宗岳,高会江,白秀娟,等.应用不同模型估计虹鳟生长性状的遗传参数[J].水产学杂志,2009,22(1):10-14.
[17] 李镕,白俊杰,李胜杰,等.大口黑鲈生长性状的遗传参数和育种值估计[J].中国水产科学,2011,18(4):766-773.
[18] Liu Yongxin,Wang Guixing,Wang Yufen,et al.Estimation of genetic parameters for growth traits of Japanese flounder Paralichthys olivaceus using an animal model[J].Fisheries Science,2011,77(1):87-93.
[19] Gjerde B,Terjesen B F,Barr Y,et al.Genetic variation for juvenile growth and survival in Atlantic cod(Gadus morhua)[J].Aquaculture,2004,236(1-4):167-177.
[20] Martínez V,Neira R,Gall G A E.Estimation of genetic parameters from pedigreed populations:lessons from analysis of alevin weight in coho salmon(Oncorhynchus kisutch)[J].Aquaculture,1999,180(3-4):223-236.
[21] 刘小林,常亚青,相建海,等.虾夷马粪海胆早期生长发育的遗传力估计[J].中国水产科学,2003,10(3):206-211.
[22] Benzie J A H,Kenway M,Trott L.Estimates for the heritability of size in juvenile Penaeus monodon prawns from half-sib mating [J].Aquaculture,1997,152(1-4):49-53.
[23] 杨创业,吴丹阳,王庆恒,等.马氏珠母贝生长性状遗传力估计[J].中国农学通报,2015,31(23):25-29.
[24] Su Guosheng,Liljedahl L E,Gall G A E.Genetic and environmental variation of body weight in rainbow trout(Oncorhynchus mykiss)[J].Aquaculture,1996,144(1-3):71-80.
[25] 顾宪明,刘永新,王凯,等.应用DF-REML法估计牙鲆性状遗传参数[J].东北农业大学学报,2008,39(11):71-75.
Heritability estimating and correlativity of growth traits in redfin puffer Takifugu rubripes
YU Hai-long1,GU Chen-lei1,KONG De-rong1,WEN Tao1,CUI Jun1,WANG Xiu-li1,JIANG Zhi-qiang1,QIU Xue-mei1,LIU Sheng-chong2,LI Rong3,ZHANG Tao2
(1.Key Laboratory of Mariculture&Stock Enhancement in North China's Sea,Ministry of Agriculture,Dalian Ocean University,Dalian 116023,China;2.Dalian Tianzheng Industrial Corporation Limited,Dalian 116011,China;3.Dalian Fugu Industrial Corporation Limited,Dalian 116400,China)
The phenotypic values of growth traits including body weight,body length and total body length were measured in 504 individuals(210 days old)and 500 individuals(360 days old)of redfin puffer Takifugu rubripes from five full-sib families to estimate the heritability and correlativity of growth traits in the redfin puffer.The results showed that the heritability of 210 days old individuals was 0.54 in body weight,0.38 in body length,and 0.52 in total body length and the heritability of 360 days old fish 0.46 in body weight,0.37 in body length,and 0.44 in total body length,the heritability of these three traits within secondary and higher heritability.There was the genetic correlation of 0.99 between body weight and body length,0.99 between body weight and total body length,and 0.98 between body length and total body length in 210 days old redfin puffer.In the 360 days old fish,the genetic correlation was found to be 0.97 between body weight and body length,0.97 between body weight and total body length,and 0.96 between body length and total body length,with highly positive genetic correlations. The regression analysis of phenotypic values of growth traits including body weight,body length and body height from 296 samples of 102 days old redfin puffer revealed that the mathematical model of growth traits(body weight,W;body length,L and body height,H):W=e-8.513L2.046H0.742(R2=0.903).The findings provide an important foundation for breeding of redfin puffer.
Takifugu rubripes;growth trait;heritability;genetic correlation;regression analysis
S917
A
10.16535/j.cnki.dlhyxb.2016.04.002
2095-1388(2016)04-0357-05
2016-04-28
大连市科技计划项目 (2014B11NC091);国家海洋公益性行业科研专项 (201405003)
于海龙 (1989—),男,硕士研究生。E-mail:1131568827@qq.com
王秀利 (1964—),男,博士,教授。E-mail:xlwang@dlou.edu.cn
