基于ArcGIS的南麂列岛潮间带大型底栖藻类研究
2016-09-24姚启学宋伟华蔡厚才
姚启学, 宋伟华, 蔡厚才, 王 飞
(浙江海洋学院水产学院,浙江省海洋渔业装备技术研究重点实验室,浙江舟山 316022)
基于ArcGIS的南麂列岛潮间带大型底栖藻类研究
姚启学, 宋伟华*, 蔡厚才, 王 飞
(浙江海洋学院水产学院,浙江省海洋渔业装备技术研究重点实验室,浙江舟山 316022)
根据 2013 年 11月(秋)和2014年2 月(冬)、5 月(春)、8 月(夏)在南麂列岛海洋自然保护区潮间带调查所获大型底栖藻类数据,对南麂列岛海洋自然保护区潮间带大型底栖藻类的种类组成、数量分布、优势种以及多样性进行研究。将调查数据整理并录入到南麂列岛生物多样性地理信息系统,使用ArcGIS软件绘制南麂列岛潮间带调查断面分布图、藻类生物量分布图和调查断面藻类多样性指数专题图。使用ArcGIS软件进行距离分析和地形分析,并分别结合藻类调查数据进行研究。结果表明,调查断面与南麂岛之间的距离和调查断面春季生物量之间存在弱相关关系,而与生物多样性指数之间不存在相关关系。除在10°~20°坡度范围内调查断面的生物量异常外,生物量随着坡度的增加而减少,并且藻类的群落结构随着坡度的改变而发生变化;就全年潮间带大型底栖藻类生物量而言,阴坡坡向藻类生物量>阳坡坡向藻类生物量>半阳坡坡向藻类生物量>半阴坡坡向藻类生物量,并且不同坡向的大型底栖藻类生物量随季节的变化程度存在明显差异。
地理信息系统;大型底栖藻类;生物多样性
南麂列岛国家级海洋自然保护区是海洋生物“南种北移,北种南移”的资源库,在海洋生态中有着极其重要的研究价值。南麂列岛潮间带大型藻类资源丰富,国内相关单位和科研人员也做了大量研究。结合历史研究资料可知,近20年来随着全球气候变暖,加上人为活动的影响,南麂列岛潮间带大型藻类自然资源已经出现衰退现象,海藻生物多样性下降明显。针对这种情况,通过科学而全面的资源调查掌握南麂列岛潮间带大型藻类的时空分布格局,并以高效合理的方式存储调查数据和直观灵活的表现形式展示调查成果,为其资源修复和管理决策提供科学依据已成为当前迫切需要解决的问题。
笔者根据2013年11月(秋)、2014年2月(冬)、2014年5月(春)和2014年8月(夏)在南麂列岛主要岛礁区(大擂岛、大山脚、国姓岙、后麂山、火焜岙、龙船礁、马祖岙、破屿、三盘尾、山角寮、上马鞍东南、上马鞍西北、下马鞍、小柴屿、朱屿)潮间带15个断面调查数据,定量分析南麂列岛潮间带大型藻类的种类组成、数量分布、优势种、多样性等,并运用ArcGIS软件对调查数据进行存储和制图,以了解潮间带大型藻类时空分布上的变化,为资源修复和管理决策提供科学依据;此外,结合南麂列岛物多样性地理信息系统(NIBGIS)中基础地理数据库的数据,运用ArcGIS软件计算各调查断面和南麂岛的距离,分析潮间带大型藻类的生物量及多样性与距离南麂岛位置远近的相关关系,由于南麂列岛人类的活动主要集中在南麂岛及其周围海域,如果有明显的相关关系则可以初步判断人类活动是影响南麂列岛潮间带大型藻类空间分布的重要因素;借助南麂列岛等高线、等深线和高程点图层数据,建立DEM(数字高程模型),获取各调查断面的地形信息,提取各调查断面的坡度和坡向数据,研究坡度和坡向对南麂列岛潮间带大型藻类的影响。
1 研究方法
1.1数据来源及数据支持藻类数据源自2013年11月(秋)、2014年2月(冬)、2014年5月(春)和2014年8月(夏)在南麂列岛海洋自然保护区开展的潮间带资源调查中获得的潮间带藻类资源资料。
距离量算、制图以及建立DEM(数字高程模型)所用的地理信息图层数据来源于南麂列岛生物多样性地理信息系统中的地理空间数据库,数据库设计开发单位为国家海洋局第二海洋研究所,使用及更新管理单位为南麂列岛国家海洋自然保护区管理局;距离量算,制图及建立DEM提取坡度、坡向数据所使用的软件为ArcGISDesktop9.3,提供单位为南麂列岛国家海洋自然保护区管理局。
1.2调查断面分别在南麂列岛的大擂岛、大山脚、大山脚砾石滩、国姓岙、后麂山、火焜岙、龙船礁、马祖岙、破屿、三盘尾、三角寮、上马鞍东南、上马鞍西北、下马鞍、小柴屿、竹屿各布设1条断面(图1)。
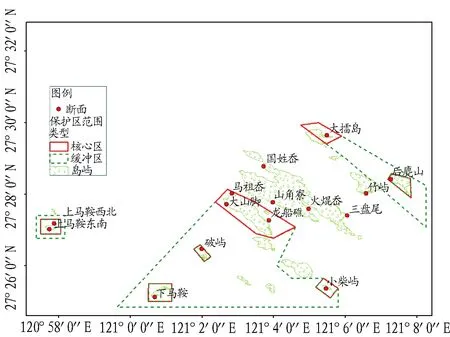
图1 南麂列岛国家海洋自然保护区潮间带调查断面Fig.1 Sampling sectionsof intertidal zone in Nanji islands marine nature reserve
1.3数据处理多样性指数采用Shannon-Wiener多样性指数的计算公式[1-3],均匀度指数采用Pielou指数的计算公式[3-4],丰富度指数采用Margalef指数的计算公式[3,5],各计算公式如下:
Shannon-Wiener多样性指数:
(2)
Pielou均匀度指数:J′=H′lnS
(3)
Margalef丰富度指数:D=(S-1)lnN
(4)
式中,H′为藻类种类多样性,S为藻类总种类数,ni为大型藻类物种i的生物量,N为各断面中所有大型藻类的生物量,J′为藻类种类均匀度,D为藻类丰富度。
2 地理信息系统中距离定义及坡度、坡向提取
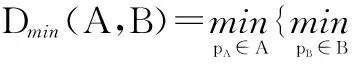

2.2坡度和坡向的计算方法坡度(Slope)是地表单元陡缓的程度,其计算公式为:
α=arctan(h/l)
(5)
式中,α为坡度值,h为坡面的垂直距离,l为坡面的水平距离。
坡向(Aspect)是指坡面法线在水平面上投影的方向。坡向的描述共有定量和定性2种方式。定量描述方式是以坡面法线在水平面上的投影与正东方的水平夹角,其中正北方为0°(360°),顺时针递增,正东方为90°,正南方为180°,正西方为270°。定性描述又可以分为四方向法和八方向法。四方向法包括阳坡(定量135°~225°)、半阳坡(定量90°~135°和225°~270°)、阴坡(定量45°~315°)和半阴坡(定量45°~90°和270°~315°);八方向法则可以分为东、东南、西、西北、南、西南、北、东北8个方向。
3 基于GIS的藻类时空分布分析
3.1生物量的时空变化从图2可以看出,各断面大型底栖藻类生物量分布具有明显的差异,且季节性变化明显。大型底栖藻类生物量最高的断面是大擂岛,其平均生物量接近670g/m2;其次分别是竹屿、三盘尾和小柴屿,平均生物量介于300~400g/m2;断面生物量明显少于其他断面的主要有国姓岙、火焜岙和处于核心区的破屿,平均生物量少于20g/m2,其中国姓岙平均生物量不足3g/m2,这3个断面的秋冬季平均生物量大多为0。整体来看,根据各断面位置和平均生物量可以将断面分为3个部分:南麂列岛国家海洋自然保护区东部潮间带断面(大擂岛、后麂山、竹屿、三盘尾、小柴屿)、中部潮间带断面(国姓岙、火焜岙、山角寮、龙船礁、马祖岙、大山脚、破屿)、西部潮间带断面(上马鞍东南、上马鞍西北、下马鞍),3个部分断面藻类的平均生物量从大到小依次为东部潮间带藻类生物量、西部潮间带藻类生物量、中部潮间带生物量。

图2 南麂列岛潮间带不同断面大型底栖藻类的生物量分布Fig.2 Biomass distribution of macroalgae in different sections in Nanji islands marine nature reverse
从季节来看,春季南麂列岛国家海洋自然保护区潮间带大型底栖藻类生物量最高,15条断面春季的大型底栖藻类平均生物量为328.74g/m2;其次是秋季,平均生物量为211.15g/m2;冬季断面大型底栖藻类平均生物量接近秋季,为210.74g/m2;夏季断面大型底栖藻类平均生物量最少,为151.28g/m2,不到春季平均生物量的50%。
3.2多样性分析从图3可以看出,Shannon-Winner多样性指数(H′)春季最高值出现在大擂岛断面,最低值出现在大山脚断面。南麂列岛国家海洋自然保护区潮间带春季H′最高的4个断面为大擂岛、后麂山、龙船礁和马祖岙,都在核心保护区;夏季H′的最大值出现在小柴屿,并且小柴屿夏季H′成为该断面4个季节中的最大值,这与其他14个断面(春季为最大值)是不同的;秋季H′的最大值同样出现在大擂岛,龙船礁、上马鞍西北和后麂山H′值同样高于其他断面;冬季H′的最大值出现在上马鞍西北,其次是山角寮,大山脚、国姓岙、火焜岙和破屿由于定量采集中没有采集到藻类,因此H′为0。
Margalef丰富度指数(D)的时空分布的主要特点是在大多数断面春季的Margalef丰富度指数明显高于其他季节,但是在小柴屿和破屿夏季Margalef丰富度指数高于其他季节,并且明显高于夏季其他断面。南麂列岛国家海洋自然保护区潮间带藻类Margalef丰富度指数(D)的时空分布如图4所示。南麂列岛国家海洋自然保护区潮间带藻类Pielou均匀度指数(J′)的时空分布如图5所示。

图3 南麂列岛国家海洋自然保护区潮间带藻类群落Shannon-Wiener多样性指数(H′)的时空分布Fig.3 The spatial and temporal distribution of marco-algae Shannon-Wiener index (H′ ) of Nanji islands marine nature reserve

图4 南麂列岛国家海洋自然保护区潮间带藻类群落Margalef丰富度指数(D)的时空分布Fig.4 The spatial and temporal distribution of marco-algae Margalef index (D) of Nanji islands marine nature reserve

图5 南麂列岛国家海洋自然保护区潮间带藻类群落Pielou均匀度指数(J′)的时空分布Fig.5 The spatial and temporal distribution of marco-algae Pielou index (J′)of Nanji islands marine nature reserve
4 人类活动对大型藻类的影响分析
4.1最近距离和质心距离的求解由于南麂列岛国家海洋自然保护区人类活动(生活生产和旅游)主要集中在南麂岛上的行政村及其周围,大擂岛和竹屿岛内行政村的村民大多生活在内陆,但户口在行政村内,因此在研究时需要将这2个岛内村庄的影响去除。在计算最近距离和质心距离时,在行政村点图层中将大擂岛内的大擂山村和竹屿岛内的竹屿村删除。任一调查断面到南麂岛内行政村的欧氏空间距离的最小值就是该调查断面和行政村之间的最近距离;任一调查断面到所有行政村的质量中心(几何中心)之间的欧氏空间距离就是该调查断面和行政村之间的质心距离。该部分数据处理及距离量算使用ArcGIS软件完成,各调查断面和行政村之间的最近距离和质心距离结果如表1所示。

表1 调查断面和行政村之间的距离
4.2最近距离与大型底栖藻类的相关性为了定量分析南麂列岛国家海洋自然保护区各潮间带调查断面大型底栖藻类与人类活动的相关性,选取春季各调查断面大型底栖藻类生物量以及生物多样性指数(春季生物量以及藻类种类数明显高于其他季节,更具代表性),进行双变量相关性分析,相关性关系采用Pearson相关系数(r)进行量化。Pearson相关系数(r)的取值范围为-1~1;当r大于1时,2个变量之间呈正相关关系,当r小于1时,2个变量之间呈负相关关系;当r等于-1或1时,2个变量之间呈完全相关;当r等于0时,2个变量之间完全不相关;当0<丨r丨≤0.3时,2个变量之间呈微弱相关,可以认为不相关;当0.3<丨r丨≤0.5时,2个变量之间呈低度相关;当0.5<丨r丨≤0.8时,2个变量之间呈显著相关;当0.8<丨r丨<1时,2个变量之间高度相关[6]。
各调查断面到行政村最近距离与春季各调查断面大型底栖藻类生物量的Pearson相关系数(r)为0.444,与春季各调查断面Shannon-Wiener多样性指数之间的Pearson相关系数(r)为0.052,与春季各调查断面Pielou均匀度指数之间的Pearson相关系数(r)为0.110,与春季各调查断面Margalef丰富度指数之间的Pearson相关系数(r)为-0.204。调查断面到行政村的最近距离与春季调查断面大型底栖藻类生物量之间存在低度相关,而与群落多样性指数之间存在微弱相关关系。
4.3质心距离与大型底栖藻类的相关性分析南麂列岛国家海洋自然保护区各潮间带调查断面大型底栖藻类与质心距离之间的相关性分析表明,各调查断面到行政村质心距离与春季各调查断面大型底栖藻类生物量之间的Pearson相关系数(r)为0.442,与春季各调查断面Shannon-Wiener多样性指数之间的Pearson相关系数(r)为0.036,与春季各调查断面Pielou均匀度指数的Pearson相关系数(r)为0.068,与春季各调查断面Margalef丰富度指数之间的Pearson相关系数(r)为-0.208。此结果与最近距离和春季各调查断面大型底栖藻类生物量之间的Pearson相关系数(r)计算结果高度相似,说明在南麂列岛国家海洋自然保护区分别使用最近距离和质心距离进行人类活动对大型底栖藻类的影响的研究结果相同。
5 地形因素对大型底栖藻类空间分布的影响
5.1TIN的建立及坡度、坡向提取在陆地上,地形通过地貌过程直接对植被产生作用,并可以通过地形起伏变化控制资源因子的空间再分配,间接影响陆地植物群落[7]。国内学者对地形因素与陆地植物群落之间的关系进行了广泛研究[8-14],但是地形因素对潮间带大型底栖藻类影响的研究在国内较少报道。该研究使用ArcGIS软件根据现有高程数据创建南麂列岛国家海洋自然保护区不规则三角网(TIN)数字高程模型,根据数字高程模型生成坡度和坡向专题图,提取不同断面的坡度和坡向数据,分析坡度、坡向对南麂列岛国家海洋自然保护区潮间带大型底栖藻类的影响。
研究地形因素对大型底栖藻类空间分布的影响要尽可能去除无关因素对研究结果的影响,分布在南麂岛周围潮间的国姓岙、三角寮和火焜岙调查断面带受附近行政村人类活动的影响明显,因此在研究地形因素对大型底栖藻类空间分布的影响时将不考虑这3个调查断面的数据。
根据坡度和坡向栅格图层分别提取调查断面所在地理位置的坡度、坡向数据,结果如表2所示。坡度使用角度表示,在存储坡度的栅格图层上进行坡度等级划分时,坡度为10°为1个等级,共分为4个等级: 0°~10°、10°~20°、20°~30°、30°~40°;ArcGIS软件生成的坡向栅格数据使用定量法描述,并使用八方向法根据栅格数值进行分类,根据调查断面所处栅格的坡向数值使用四方向法定性描述。该研究调查断面所处地理位置的坡度和坡向不是现场调查时测量获取,而是通过手持GPS接收机记录调查断面坐标值,后期结合ArcGIS软件空间分析功能求出对应调查断面的坡度和坡向。手持GPS接收机误差范围通常在10m左右,加之部分调查断面周围地形(比如陡峭的山坡)对卫星信号的阻碍作用,造成获取的部分调查断面地理位置误差较大。为了减小误差对研究的影响,分别将坡度图层、坡向图层与岛屿图层、等高线图层、潮间带图层、调查断面图层进行叠加分析,判断调查断面地理位置数据的准确性。对于存在明显地理位置错误的调查断面坡度和坡向数据,使用该调查断面临近海岸线的多个栅格单元的平均值代替。

表2 各调查断面的坡度和坡向
5.2大型底栖藻类对坡度的响应由表2可知,坡度范围在0°~10°的调查断面分别为大擂岛、龙船礁、三盘尾和竹屿,除了龙船礁调查断面生物量较低(龙船礁4个季度的生物量平均值为207.83g/m2)外,其余3个调查断面生物量均超过300g/m2,大擂岛生物量更是超过了600g/m2,这4个调查断面的四个季度生物量平均值为383.65g/m2;坡度范围在10°~20°的调查断面分别为后麂山、马祖岙、破屿和上马鞍东南共4个调查断面,生物量平均值为152.51g/m2,其中上马鞍东南调查断面生物量明显高于其他3个调查断面的生物量,其4个季度生物量的平均值接近300g/m2,破屿调查断面生物量显著低于其他3个调查断面生物量,4个季度生物量的平均值不足20g/m2;坡度范围在20°~30°的调查断面分别为上马鞍西北、下马鞍和小柴屿共3个调查断面,4个季度生物量平均值为276.07g/m2;大山脚调查断面所处位置的地形比较陡峭,其坡度范围为30°~40°,4个季度生物量的平均值为176.46g/m2。整体来看,大型底栖藻类生物量随着坡度的增加呈现递减趋势,但是坡度范围在10°~20°内的调查断面生物量都不是很丰富,造成这一现象的原因还有待进一步研究。
春季,在0°~10°坡度范围内的调查断面共定性定量采集到大型底栖藻类31种,该范围内4个调查断面采集到大型底栖藻类种类数的平均值为16。定量采集到的大型底栖藻类主要是珊瑚藻,生物量占总生物量的46.81%;其次是瓦氏马尾藻,生物量占总生物量的10.78%;再次是厚网藻,生物量占总生物量的8.06%,其他大型底栖藻类生物量明显不如以上3种藻类。在10°~20°坡度范围内,4个调查断面共定性定量采集到大型底栖藻类33种,该范围内调查断面采集到大型底栖藻类种类数的平均值为18。定量采集到的大型底栖藻类主要为珊瑚藻,生物量占总生物量的44.96%;其次为鼠尾藻,生物量占总生物量的16.20%;再次为厚网藻和羊栖菜,生物量分别占总生物量的9.46%和8.15%。在20°~30°坡度范围内共定性定量采集到大型底栖藻类38种,该范围内3个调查断面采集到大型底栖藻类种类数的平均值为19。其中砺菜生物量最多,占总生物量的34.30%;其次是鼠尾藻,生物量占总生物量的17.66%;再次是厚网藻,生物量占总生物量的14.31%,扁节荚藻在该坡度范围内也有一定的生物量,占总生物量的11.38%。在30°~40°坡度范围内只有大山脚调查断面,共定性定量采集到大型底栖藻类6种,鼠尾藻生物量占据绝对优势,占总生物量的93.62%,其余定量采集到的大型底栖藻类是少量的半丰满鞘丝藻和石莼。
夏季在0°~10°坡度范围内共定性定量采集到大型底栖藻类8种,鼠尾藻成为夏季该坡度范围内的绝对优势种,生物量占总生物量的75.53%;其次分别为铁钉菜和鸡毛菜,生物量各占总生物量的16.15%和6.02%。在10°~20°坡度范围内,4个调查断面共定性定量采集到大型底栖藻类8种,其中定量采集到的藻类只有3种,分别为鼠尾藻、铁钉菜和荆刺藻,生物量分别占总生物量的82.32%、17.51%和0.17%。在20°~30°坡度范围内,共定性定量采集到大型底栖藻类19种,主要大型底栖藻类生物量由多到少依次为鼠尾藻、瓦氏马尾藻、小衫藻、岗村凹顶藻、苔垢菜,生物量分别占总生物量的42.31%、14.97%、7.48%、7.46%、5.70%;在30°~40°坡度范围内,共定量采集到2种大型底栖藻类,分别为珊瑚藻和鼠尾藻,生物量分别占总生物量的92.52%和7.48%。
秋季在0°~10°坡度范围内共定性定量采集到大型底栖藻类13种,其中主要为珊瑚藻,生物量占总生物量的67.21%;其次是鼠尾藻,生物量占总生物量的23.63%;再次为坛紫菜和铁钉菜,生物量均不足10%;秋季在10°~20°坡度范围内共定性定量采集到大型底栖藻类12种,其中珊瑚藻为绝对优势种,生物量占总生物量的73.99%;其次为铁钉菜,生物量占总生物量的24.58%。在20°~30°坡度范围内,共定性定量采集到大型底栖藻类9种,珊瑚藻为秋季该坡度范围内的绝对优势种,生物量占总生物量的92.33%;其次为鼠尾藻,生物量仅占总生物量的6.44%。秋季在30°~40°坡度范围内共定性采集到大型底栖藻类2种,定量则没有采集到大型藻类。
冬季在0°~10°坡度范围内共定性定量采集到大型底栖藻类29种,主要为珊瑚藻、坛紫菜和鼠尾藻,生物量分别占总生物量的55.01%、32.45%和10.34%;冬季在10°~20°坡度范围内共定性定量采集到大型底栖藻类11种,其中珊瑚藻成为冬季该坡度范围内的绝对优势种,生物量占总生物量的93.84%;其次为鼠尾藻和坛紫菜,生物量占总生物量比例不足4%;在20°~30°坡度范围内共定性定量采集到大型底栖藻类18种,其中主要为坛紫菜、珊瑚藻、鼠尾藻和石莼,生物量分别占总生物量的50.57%、21.69%、15.26%、7.25%;在30°~40°坡度范围内共定性定量采集到大型底栖藻类3种,只定量采集到珊瑚藻一种大型底栖藻类。
根据上述4个季度分坡度大型底栖藻类统计数据,发现在:30°~40°坡度范围内潮间带生物多样性较差,夏季20°~30°坡度范围内大型底栖藻类多样性明显优于其他坡度范围内物种多样性,冬季0°~10°坡度范围内大型底栖藻类多样性优于其他坡度范围内物种多样性。从南麂列岛潮间带大型底栖藻类群落结构分析,0°~10°坡度范围和10°~20°坡度范围内藻类群落结构相似,2个坡度范围内各个季节位于主导地位的藻类完全相同,20°~30°坡度范围内潮间带藻类除了秋季群落结构和以上2个坡度范围内藻类群落结构相似外,其他3个季节有明显差异性。
5.3大型底栖藻类对坡向的响应坡向为阳坡的调查断面有大山脚、三盘尾、竹屿、下马鞍、小柴屿、上马鞍东南共6个调查断面,这6个调查断面的4个季节生物量平均值为290.19g/m2;半阳坡坡向的调查断面只有后麂山调查断面,生物量为199.93g/m2;属于半阴坡坡向的调查断面分别为龙船礁、马祖岙、破屿,共3个调查断面,该坡向的调查断面4个季节生物量平均值为111.76g/m2;属于阴坡坡向的调查断面共有2个,分别为大擂岛和上马鞍西北,该坡向调查断面4个季节生物量的平均值为436.48g/m2。就全年潮间带大型底栖藻类生物量而言,阴坡坡向藻类生物量>阳坡坡向藻类生物量>半阳坡坡向藻类生物量>半阴坡坡向藻类生物量。
春季大型底栖藻类生物量,半阳坡坡向生物量(433.97g/m2)>阴坡坡向生物量(411.85g/m2)>阳坡坡向生物量(409.56g/m2)>半阴坡坡向生物量(222.79g/m2);夏季,阴坡坡向生物量(532.6g/m2)>阳坡坡向生物量(134.19g/m2)>半阳坡坡向生物量(73.67g/m2)>半阴坡坡向生物量(64.76g/m2);秋季,阴坡坡向生物量(479.79g/m2)>阳坡坡向生物量(260.08g/m2)>半阳坡坡向生物量(184.04g/m2)>半阴坡坡向生物量(86.89g/m2);冬季,阳坡坡向生物量(356.52g/m2)>阴坡坡向生物量(321.66g/m2)>半阳坡坡向生物量(108.07g/m2)>半阴坡坡向生物量(72.6g/m2)。由此可见,不同坡向的大型底栖藻类生物量随季节的变化程度是不同的,阴坡坡向生物量随季节的变化不是特别明显,4个季节的藻类生物量数值都较高,其他3个坡向的藻类生物量变化则比较明显。
6 结论
笔者根据 2013 年 11月(秋)和1014年2 月(冬)、5 月(春)、8 月(夏)4个季节在南麂列岛海洋自然保护区潮间带调查所获得的大型底栖藻类数据,结合ArcGIS软件对南麂列岛海洋自然保护区潮间带大型底栖藻类的种类组成、数量分布、优势种以及多样性进行了研究,得出以下结论:
(1)生物量总体上呈现出南麂列岛自然保护区东部藻类生物量>西部藻类生物量>中部生物量的趋势,各季节平均生物量从高到低依次为春季、冬季、秋季、夏季。
(2)Shannon-Winner指数(H′)春季最高,冬季次之,夏季略低于冬季,秋季最低;Margalef丰富度指数(D)的季节变化与Shannon-Winner指数(H′)相同;各季节Pielou均匀度指数(J′)从大到小依次为春季、夏季、冬季、秋季。
(3)调查断面与南麂岛之间的距离和调查断面春季生物量之间存在弱相关关系,与生物多样性指数之间不存在相关关系。
(4)除在10°~20°坡度范围内调查断面的生物量异常外,生物量随着坡度的增加而减少,并且藻类的群落结构随着坡度的改变而发生变化;就全年潮间带大型底栖藻类生物
量而言,阴坡坡向藻类生物量>阳坡坡向藻类生物量>半阳坡坡向藻类生物量>半阴坡坡向藻类生物量,并且不同坡向的大型底栖藻类生物量随季节的变化程度存在明显差异。
[1] 马克平.生物群落多样性的测度方法Ⅰα多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168.
[2] 马克平,刘玉明.生物群落多样性的测度方法Ⅰα多样性的测度方法(下)[J].生物多样性,1994,2(4):131-139.
[3] 沈国英,施并章.海洋生态学[M].北京:科学出版社,2006:162.
[4]PIELOUEC.Mathematicalecology[M].NewYork:Wiley,1977.
[5]MARGALEFDR.Informationtheoryinecology[J].Geneneralsystem, 1957, 3: 36-71.
[6] 孙逸敏.利用SPSS软件分析变量间的相关性[J].新疆教育学院学报,2007,23(2):120-123.
[7] 张旭,国庆喜.地形对天然次生林空间格局的影响[J].东北林业大学学报,2007,35(1):68-75.
[8] 马旭东,张苏峻,苏志尧,等.车八岭山地常绿阔叶林群落结构特征与微地形条件的关系[J].生态学报,2010,30(19):5151-5160.
[9] 马建梅,刘金福,郑世群.戴云山自然保护区不同地形植物群落的物种α多样性研究[J].武夷科学,2013,29(00):49-55.
[10] 侯兆疆,赵成章,李钰,等.高寒退化草地狼毒种群地上生物量空间格局对地形的响应[J].生态学杂志,2013,32(2):253-258.
[11] 刘海丰,薛达元,桑卫国.地形因子对暖温带森林群落物种丰富度-地上生物量关系的影响[J].生态环境学报,2012,21(8):1403-1407.
[12] 方楷,宋乃平,魏乐,等.荒漠草原不同地形条件下土壤水分和地上生物量的时空分异[J].干旱区研究,2012,29(4):641-647.
[13] 栾忠平,吴湘菊.森林在不同坡度上分布特征的研究[J].吉林林业科技,2012,41(1):23-26.
[14] 刘金根,薛建辉.坡度对香根草护坡植物群落早期特征的影响[J].草原与草坪,2010,30(2):56-61.
StudyontheMacroalgaeBasedontheArcGISinNanjiIslandsMarineNature
YAOQi-xue,SONGWei-hua*,CAIHou-caietal
(KeyLaboratoryofMarineFisheryEquipmentandTechnologyofZhejiang,FisherySchoolofZhejiangOceanUniversity,Zhoushan,Zhejiang316022)
AccordingtoNovember2013 (autumn)andFebruary2014 (winter),May(spring)andAugust(summer)amongNanjiIslandsMarineReserve,macroalgaedatawasinvestigated,thespecies,quantitydistribution,dominantspeciesanddiversitywasstudied.ThesurveydatawasreorganizedandenteredintoNanjiIslandsbiodiversityGIS,ArcGISsoftwarewasusedtodrawsurveysectiondistributionmaps,mapsofalgalbiomass,algaediversityindexsectionalsurveymap.UsingArcGISsoftwarefordistanceanalysisandterrainanalysisandtheresultswerecombinedwiththealgaeresearchsurveydata.TheresultsshowedthattherewasaweakcorrelationbetweendistanceofsectionandNanjiIslands,andbiomassinspring,andtherewasnocorrelationamongthebiodiversityindex.Exceptintherangeof10°-20°slopesectionofthesurveybiomassabnormalities,biomassdecreasedwithincreasingslope,algalcommunitystructurechangedwithslope;onthewholeyearofmacroalgaebiomass,theshadyslopeofalgalbiomass>sunnyslopeofalgalbiomass>semi-sunnyslopeofalgalbiomass>semi-shadyslopeofalgalbiomass,andtherewassignificantdifferenceinvariationdegreeofdifferentaspectsmacroalgaebiomasswithseason.
Geographicinformationsystems;Macroalgae;Biologicaldiversity
姚启学(1987- ),男,山东菏泽人,硕士研究生,研究方向:渔具渔法与渔业工程技术。*通讯作者,教授,博士,硕士生导师,从事海洋渔业研究。
2016-05-23
S932.7
A
0517-6611(2016)18-011-05
