大豆孢囊线虫大豆和烟草群体寄生性比较分析
2016-09-14刘海勤刘泉成赵洪海
刘海勤, 刘泉成, 赵洪海
(青岛农业大学农学与植物保护学院,山东省植物病虫害防控重点实验室,青岛 266109)
大豆孢囊线虫大豆和烟草群体寄生性比较分析
刘海勤,刘泉成,赵洪海*
(青岛农业大学农学与植物保护学院,山东省植物病虫害防控重点实验室,青岛266109)
通过盆栽试验,测定大豆孢囊线虫(Heteroderaglycines,soybean cyst nematode, SCN)大豆群体(SCNS)和烟草群体(SCNT)对5个大豆品种(SCN生理小种鉴别寄主)和9个烟草栽培品种的寄生性。结果发现,SCNS在5个大豆品种上的平均繁殖系数(Rf=Pf/Pi)为0.60(0.14~1.90),在感病对照品种‘Lee’上平均Rf高达1.90;SCNT在5个大豆品种上的平均Rf为0.18(0.06~0.44),在感病对照品种‘Lee’上Rf为0.44。SCNS对9个烟草品种几乎无寄生性,平均Rf为0.06(0.00~0.10);SCNT则能寄生9个烟草品种,平均Rf为1.26(0.45~1.95)。接种35 d后,在不同大豆品种根内SCNS和SCNT的幼虫数量均极少,侵染率基本相同;在不同烟草品种根内SCNS的幼虫数量均为0,而SCNT的幼虫数量平均为26.5(14.6~57.0)。研究表明,供试SCNS属于14号生理小种,但供试SCNT与之寄生性差异大,属于新的SCN致病类型。
大豆孢囊线虫;大豆;烟草;繁殖系数;寄生性;新致病类型
大豆孢囊线虫(Heteroderaglycines,soybean cyst nematode, SCN)属于线虫门、垫刃目、异皮科,是世界上危害大豆最严重的植物病原线虫之一[1-2]。SCN在我国黑龙江、吉林、辽宁、内蒙古、北京、河北、山西、山东、河南、江苏、安徽等16省(直辖市、自治区)有发生,每年大豆受害面积超过150万hm2,严重发病地块减产30%以上,甚至绝收[3]。SCN在我国除危害大豆外,还可侵染烟草、地黄、泡桐等作物[4-6]。
SCN是严格的专性寄生物,种内群体间存在明显的生理分化现象,而寄生大豆的SCN存在16个生理小种[7-8]。作者在调查中多次发现,在同一地点大豆和烟草上同时发生SCN,而且研究初步表明SCN大豆群体和烟草群体在田间侵染特性和寄主适应性上存在差异[9-10],但其致病类型及其对烟草不同品种的致病性尚不明确。本试验测定同一地点来源的SCN大豆和烟草群体的寄生性差异,将为SCN综合治理中的抗性利用提供依据。
1 材料与方法
1.1材料来源
SCN大豆群体和烟草群体均采自山东省诸城市辛兴农场,分别用SCNS和SCNT表示。供试大豆品种5个,为‘Lee’、‘Pickett’、‘Peking’、‘PI88788’和‘PI90763’,即SCN生理小种鉴别寄主,由沈阳农业大学北方线虫研究所提供。供试烟草品种9个,为‘中烟100’、‘中烟103’、‘中烟201’、‘中烟202’、‘安烟2号’、‘龙江911’、‘K326’、‘NC89’和‘NC95’,由中国农业科学院烟草研究所提供。大豆和烟草种植钵为塑料水杯,杯口直径6.5 cm、杯底直径4.5 cm、杯高9.0 cm。
1.2线虫接种和培育
2014年6月,每杯装灭菌砂壤土200 g后,移栽胚根长4~5 cm豆苗或3~4片真叶烟苗,每杯1株。同时进行线虫接种:在幼苗四周土壤上用玻璃棒打4个孔,孔深至根系水平,每孔置入4个压破的孢囊,用水淋冲后覆盖灭菌土封严,每杯共接种孢囊16个(约含1 500个卵),即最初孢囊群体密度(Pi)为8.0个/100 g土壤。将种植杯置温室内在22~32 ℃环境下培育。大豆和烟草每个品种重复5次。
1.3寄生性调查方法
线虫接种35 d后,扣钵取根,检查根系上和土壤中白色雌虫并计数,并对根系中的线虫进行染色、计数。记录各品种处理形成的雌虫(Female)及每株根内2龄幼虫(J2)、3龄和4龄幼虫(J3+J4)的数量。在此试验中雌虫群体密度即为最终孢囊群体密度(Pf)。
SCNS生理小种鉴定方法参考雌虫指数(female index,FI)法:FI=(供试品种每株根上平均孢囊数/感病对照品种‘Lee’上平均孢囊数)×100,当FI≥10时为感病,反之抗病。根据各鉴别寄主对SCNS的抗感反应,确定其生理小种类型[3, 7]。
SCNS和SCNT对大豆和烟草品种寄生性大小的测定采用繁殖系数(reproduction factor,Rf)法:Rf=最终孢囊群体密度(Pf)/最初孢囊群体密度(Pi)。本试验中Pf即接种35 d后形成的雌虫群体密度。Rf值越大,寄生性越强[11]。
利用DPS数据处理系统软件(DPS 14.50)中的Duncan新复极差法对试验数据进行统计分析。
2 结果与分析
2.1SCNS和SCNT对大豆的致病性
SCNS在5个SCN生理小种鉴别寄主上的致病性测定结果(表1)表明, SCNS在5个鉴别寄主上的Rf为0.14~1.90,平均为0.60,其中在感病对照品种‘Lee’上的Rf高达1.90,品种‘Lee’表现为高感。参照Riggs等[7],‘Pickett’、‘Peking’和‘PI90763’表现为感病,‘PI88788’表现为抗病,由此推断SCNS属于大豆孢囊线虫14号生理小种[3,7]。

表1 SCN鉴别寄主对SCNS的抗感反应1)Table 1 The resistance (-) and susceptibility (+) reactions of the SCN host differentials to SCNS
1) 表中雌虫数和Rf数值为5次重复的平均值±标准误,同列不同大写字母表示0.01水平差异显著性,下同。
The values for female number andRfare means of five replicates ± SE; different capital letters indicate significant difference at 0.01 level. The same below.
SCNT在5个SCN生理小种鉴别寄主上的致病性测定结果(表2)表明,SCNT在5个鉴别寄主上的Rf为0.06~0.44,平均为0.18。SCNT在感病对照品种‘Lee’上繁殖最好,但也仅产生6.4个孢囊,Rf为0.44(表明繁殖1代后群体密度下降56%),即感病对照品种‘Lee’对SCNT亦具有一定程度的抗性[3]。SCNT在其他鉴别寄主上的繁殖系数均不到0.20,只有轻微寄生性。由此得知,SCNT对SCN鉴别寄主虽然均能侵染,但相对于SCNS其致病性明显较差。由于感病对照品种‘Lee’对SCNT并非表现高感,故而用现有的这套SCN鉴别寄主无法鉴定其生理小种类型。

表2 SCNT在SCN鉴别寄主上的繁殖结果Table 2 The reproduction of SCNT on the host differentials of SCN
2.2SCNS和SCNT对烟草的致病性
SCNS和SCNT在9个烟草品种上的繁殖情况分别见表3和表4。
由表3可见,SCNS在9个烟草品种上的繁殖虽然存在极显著差异,但在繁殖最好的‘中烟103’上的Rf也仅为0.10,在9个烟草品种上的Rf平均为0.06。

表3 SCNS在不同烟草品种上的繁殖结果Table 3 The reproduction of SCNS on different tobacco cultivars
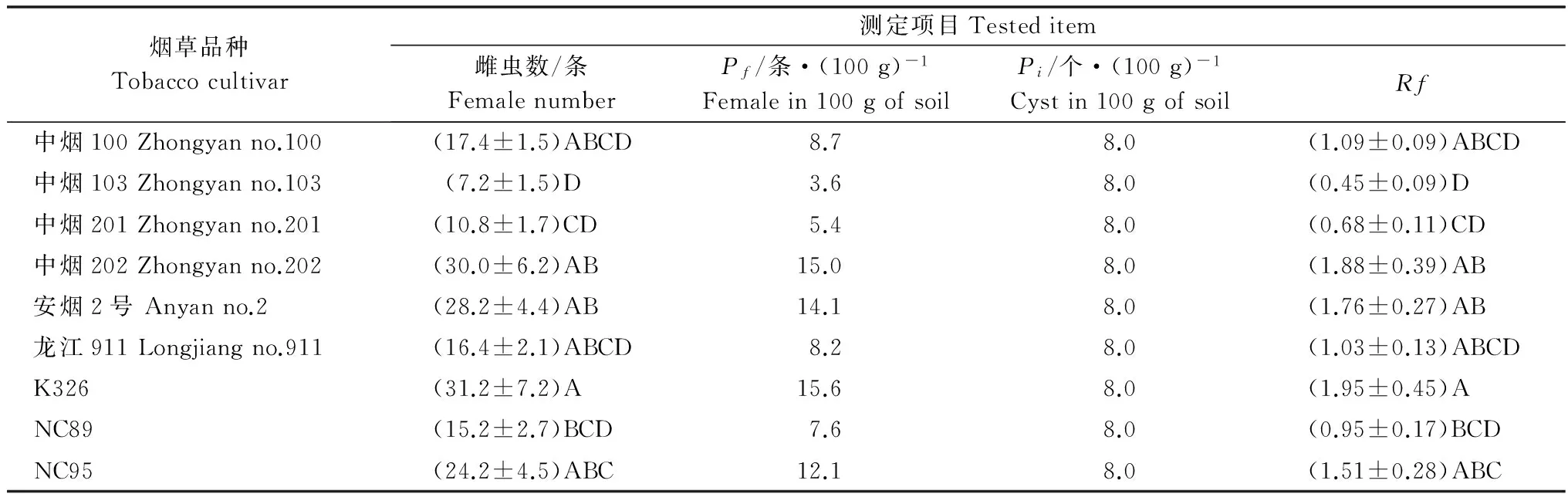
表4 SCNT在不同烟草品种上的繁殖结果Table 4 The reproduction of SCNT on different tobacco cultivars
由表4可见,SCNT在9个烟草品种上均能繁殖,但在‘K326’、‘中烟202’和‘安烟2号’上的繁殖最好,在‘中烟103’和‘中烟201’上的繁殖最差,两者Rf的差异达极显著水平(P<0.01)。SCNT在9个烟草品种上的Rf为0.45~1.95,平均为1.26,是SCNS的21倍。
由此表明,SCNS和SCNT对烟草的寄生性存在明显差异。SCNT对烟草的寄生性强,烟草是SCNT的寄主,有些品种甚至是其好寄主;但SCNS对烟草的寄生性极差,烟草是SCNS的极差寄主,甚至是其非寄主。
2.3SCNS和SCNT对大豆和烟草根系的侵染观察
大豆孢囊线虫鉴别寄主分别接种SCNS和SCNT35 d后(表5),除‘PI90763’根内有少量J2外,其他品种根内均未见J2,差异不显著(P<0.05);接种SCNS后在感病品种‘Lee’上有较多发育中的J3+J4,与其他品种存在显著性差异,而接种SCNT后不同品种根内J3+J4数量差异不显著。表明接种35 d后,SCNS和SCNT在5个鉴别寄主品种上均已完成1代,侵染率基本相同。

表5 不同大豆品种单株根内SCNS和SCNT幼虫数量1)Table 5 The juvenile number of SCNS and SCNT in single roots of different soybean cultivars
1) 表中数值为5次重复的平均值±标准误,同列不同小写字母表示0.05水平差异显著性,下同。
The values are means of five replicates±SE; different lowercase letters indicate significant difference at 0.05 level.The same below.
接种SCNS35 d后,9个烟草品种根内均无任何虫态。接种SCNT35 d后,9个烟草品种单株根系中J2数量为12.1(5.6~28.2)条,J3+J4数量为14.4(8.8~28.8)条,全部幼虫(J2+J3+J4)数量为26.5(14.6~57.0)条,其中‘中烟201’中幼虫数量最多,与其他品种存在显著性差异(P<0.05),而其他品种中的J2数量也存在显著性差异,但J3+J4数量差异不显著(表6)。

表6 不同烟草品种每株根内SCNS和SCNT幼虫数量Table 6 The juvenile numbers of SCNS and SCNT in single roots of different tobacco cultivars
3 讨论
SCN是植物的专性内寄生线虫,寄主范围相对较宽,但大多数属于豆科植物,包括大豆(Glycinemax)、赤豆(Vignaangularis)、菜豆(Phaseolusvulgaris)、绿豆(Phaseolusaureus)、豌豆(Pisumsativum)等作物,其差寄主或非寄主则主要有小麦、燕麦、大麦、黑麦、玉米、水稻、高粱、棉花、花生、甘薯、烟草、甘蔗、甜菜、番茄、甜瓜等作物[12-14]。然而,某种植物对于SCN的寄主地位并非一成不变,例如,在试验条件下SCN的一个群体在番茄等非寄主上适应多代后,可具备在这些非寄主上繁殖的能力[15]。本研究则表明,SCNT在烟草上繁殖良好,在大豆上繁殖差,烟草是其好寄主,而大豆是其差寄主;SCNS在大豆品种‘Lee’上繁殖良好,在所有烟草品种上几乎不能繁殖,大豆是其好寄主,而烟草是其极差寄主或非寄主。
SCN群体间存在明显的寄生性分化,根据其对大豆特定品种的寄生性差异,可分成不同的生理小种或HG类型[7, 16]。根据Riggs等的生理小种鉴别体系,SCN理论上存在16个生理小种,目前除12号和16号小种未被发现外,世界上已发现14个小种。其中美国SCN小种类型最多,存在全部14个已被发现的小种,优势小种为3号;其次是中国,存在1、2、3、4、5、6、7和14号共8个小种,其中黄淮大豆产区SCN的优势小种是4号,而东北大豆产区SCN的优势小种是3号[13, 17-18]。山东省是重要的夏大豆产区,SCN小种类型较为复杂,已报道有1、2、4、5和7号共5个小种,其中1号小种分布在禹城、济南、潍坊、昌邑、东营、山东半岛(胶州等),2号分布在聊城、德州,4号分布在菏泽、垦利,5号分布在东营,7号分布在山东半岛西部的潍坊、高密、昌邑[17-19]。本研究中将采自诸城市(潍坊市管辖的县级市)的SCNS群体鉴定为14号小种,属该小种在山东省的首次发现。在寄主抗性的选择压力下,SCN生理小种的变异常有发生[18, 20-27]。例如,SCN 3号小种在具备不同抗性基因的大豆上繁殖10余代后,能够变为4、6或14号小种[21, 25-26];种植大豆历史相对较短的美国却拥有世界上最多的SCN小种类型,可能是普遍种植抗线虫大豆品种所致[13]。14号小种在我国属劣势小种,之前仅在黑龙江省安达市大豆田自然发生,与优势小种3号相比其毒性较强[28-29]。
目前,侵染烟草的SCN仅发生在我国的山东和河南两省,在其他省及国外尚无SCN侵染烟草的报道[4, 9, 30]。SCNT与同一地点来源的SCNS在对大豆和烟草的寄生性上均存在明显差异,究其原因,可能与其对各自寄主的长期适应有关。波多黎各发生SCN历史很短,对于波多黎各SCN 2号小种多数木豆(Cajanuscajan)品种是免疫的,只有少数品种轻微感病,而在之28年之前已有美国SCN群体严重和中等侵染木豆的报道[31-32]。山东和河南两省位于黄淮流域,该流域可能是大豆的起源地,大豆栽培历史已有5000年上下[33]。从大豆-SCN协同进化角度看,黄淮流域SCN原本应具较为丰富的遗传多样性。中国植烟历史较短,约400年上下时间,山东和河南两省在清朝开始植烟,至今仅有350~400年时间[36]。由此可见,SCNT可能是大豆上SCN某个原始或特有种群(针对美国等国外地理区域)经过多年对烟草的适应演变而来。此外,在黄淮流域SCN侵染泡桐和地黄现象均有报道[6, 10],亦可说明该区域SCN种群存在较为多样的寄主专化性分化。
SCNS和SCNT在形态、ITS序列及SSR标记的遗传结构上(另文发表)没有区别,但寄主和环境适应性及寄生性差异明显[4, 9-10, 37],可能与它们能否克服烟草和大豆的某种/些抗性机制有关。大豆抗SCN的机制主要包括抗孵化、抗侵入和抗发育及相关的生化机制[38]。烟草中的烟碱具有杀线虫活性,长期生活在烟草根围的线虫种群能够应对烟碱高剂量处理[39]。烟草对SCNS的几乎免疫及大豆对SCNT的较高抗性(Rf≤0.44)发生在哪个线虫发育阶段,以及烟草和大豆中的某些线虫抑制物质(诸如烟碱和大豆素Ⅰ等)是否与抗孢囊线虫有关,均需要深入研究。目前,利用抗病性是控制SCN的最经济有效措施,但SCN生理小种的变异和新毒性小种的出现为抗病品种的选育和合理布局带来了难题[3, 12-14]。因而,有必要加强SCN生理小种田间监测工作,为品种抗性的科学利用提供依据。
[1]Jones J T, Haegeman A, Danchin E G J, et al. Top 10 plant-parasitic nematodes in molecular plant pathology [J]. Molecular Plant Pathology, 2013, 14(9): 946-961.
[2]Wrather J A, Anderson T R, Arsyad D M, et al. Soybean disease loss estimates for the top ten soybean-producing countries in 1998[J]. Canadian Journal of Plant Pathology, 2001, 23(2): 115-121.
[3]段玉玺. 植物线虫学[M]. 北京: 科学出版社, 2011.
[4]程子超, 赵洪海, 李建立, 等. 山东省寄生烟草的孢囊线虫种类鉴定及种内群体间rDNA-ITS-RFLP分析[J]. 植物病理学报, 2012, 42(4): 387-395.
[5]陈金堂, 李知. 为害地黄的大豆孢囊线虫的初步研究[J]. 植物病理学报, 1981, 11(1): 37-44.
[6]张东升. 大豆胞囊线虫侵染泡桐和豌豆的研究[J]. 植物病理学报, 1995, 25(3): 275-278.
[7]Riggs R D, Schmitt D P. Complete characterization of the race scheme forHeteroderaglycines[J]. Journal of Nematology, 1988, 20(3): 392-395.
[8]Winter S M J, Rajcan I, Shelp B J. Soybean cyst nematode: Challenges and opportunities[J]. Canadian Journal of Plant Science,2006,86(1):25-32.
[9]赵洪海,丁海燕,王凤龙.大豆和烟草上大豆孢囊线虫田间侵染特征比较分析[J].中国农业科学,2014,47(22):4417-4425.
[10]王慧敏,赵洪海, 王凤龙. 大豆孢囊线虫不同寄主种群的形态变异及对大豆寄主的适应性差异分析[J]. 植物保护, 2014, 40(6): 106-111.
[11]Koenning S R. Resistance of soybean cultivars to field populations ofHeteroderaglycinesin North Carolina [J]. Plant Disease, 2004, 88(9): 942-950.
[12]Noel G R.Heteroderaglycinesin soybean[J]. Nematologia Brasileira, 1993, 17(2): 103-121.
[13]Yu Q. Soybean cyst nematode (HeteroderaglycinesIchinohe)[M] ∥ El-Shemy H A. Soybean Physiology and Biochemistry. Rijeka, Croatia: InTech Europe, 2011: 461-474.
[14]Niblack T L, Tylka G L. Soybean cyst nematode management guide. Fifth ed [EB/OL]. Plant Health Initiative, 2010: 1-16[2012-04-05]. http:∥www. manitobapulse.ca/wp-content/uploads/SCN_Management.pdf.
[15]Niblack T L, Colgrove A L. Adaptability ofHeteroderaglycinesto new hosts and resistant soybean cultivars [J]. Phytopathology, 2006, 96(6): S84.
[16]Niblack T L, Arelli P R, Noel G R, et al. A revised classification scheme for genetically diverse populations ofHeteroderaglycines[J]. Journal of Nematology, 2002, 34(4): 279-288.
[17]Lu Weiguo, Gai Junyi, Li Weidong. Sampling survey and identification of races of soybean cyst nematode (HeteroderaglycinesIchinohe) in Huang-Huai Valleys[J]. Agricultural Sciences in China, 2006, 5(8): 615-621.
[18]陈品三,齐军山,王寿华,等.我国大豆胞囊线虫生理分化动态的鉴定和监测研究[J].植物病理学报,2001,31(4):336-341.
[19]陈贵省,颜清上,阎淑荣,等.大豆孢囊线虫的危害及其防治[J].作物杂志,2000(1):6-9.
[20]Cloud G L, Giggs R D, Caviness C E. Variability in host preference among field populations ofHeteroderaglycines[J]. Journal of Nematology, 1988, 20(3): 417-420.
[21]刘维志, 刘晔, 段玉玺, 等. 抗病品种对大豆孢囊线虫的选择作用[J]. 植物保护学报, 1993, 20(2): 135-137.
[22]Young L D. Changes in theHeteroderaglycinesfemale index as affected by ten-year cropping sequences [J]. Journal of Nematology, 1994, 26(4): 505-510.
[23]马书君. 黑龙江省安达地区大豆孢囊线虫生理小种动态监测[J]. 大豆科学, 1996, 15(3): 254-257.
[24]李云辉, 李肖白, 田中艳, 等. 连续种植大豆抗孢囊线虫品种胁迫线虫生理小种变异研究[J]. 大豆科学, 1998, 17(4): 370-372.
[25]于佰双,王家军,崔丽伟,等.连种抗线大豆品种(系)对大豆孢囊线虫的选择作用[J].黑龙江农业科学,1999(3):4-6.
[26]田中艳,高国金,周长军,等.大豆胞囊线虫生理小种变异研究[J].大豆科学,2007,26(2):290-292.
[27]Yu Baishuang, Duan Yuxi, Wang Jiajun, et al. Selection of virulence phenotypes from an original race 3 population ofHeteroderaglycinesby resistant cultivars [J]. Soybean Science, 2009, 28(3): 491-494.
[28]董丽民,许艳丽,李春杰,等.黑龙江省大豆胞囊线虫胞囊密度和生理小种鉴定[J].中国油料作物学报,2008,30(1):108-111.
[29]陈双,潘凤娟,周长军,等.黑龙江省安达地区大豆孢囊线虫生理小种遗传分化现象[J].土壤与作物,2015,4(1):42-47.
[30]Shi H, Zheng J. First report of soybean cyst nematode (Heteroderaglycines) on tobacco in Henan, central China[J]. Plant Disease, 2013, 97(6): 852.
[31]Smith J R, Chavarria-Carvajal J A. First report of soybean cyst nematode (Heteroderaglycines) on soybean in Puerto Rico [J]. Plant Disease, 1999, 83: 591.
[32]Smith J R, Chavarria-Carvajal J A. Host suitability of pigeon pea forHeteroderaglycinesrace 2 [J]. Nematropica, 2002, 32(2): 125-130.
[33]赵团结, 盖钧镒. 栽培大豆起源与演化研究进展[J]. 中国农业科学, 2004, 37(7): 954-962.
[36]刘国顺. 烟草栽培学[M]. 北京: 中国农业出版社, 2003: 4-5.
[37]赵洪海, 程子超, 王凤龙. 山东省烟草孢囊线虫的群体动态和世代发生特点[J]. 植物保护学报, 2013, 40(6): 529-532.
[38]吴海燕, 远方, 陈立杰, 等. 大豆胞囊线虫病与大豆抗胞囊线虫机制的研究[J]. 大豆科学, 2001, 20(4): 285-289.
[39]Yu Qing, Potter J W. Selective nematicidal activity of nicotine[J]. Journal of Food, Agriculture & Environment, 2008, 6: 428-432.
(责任编辑:田喆)
Comparative analysis of the parasitism between soybean and tobacco populations ofHeteroderaglycines
Liu Haiqin,Liu Quancheng,Zhao Honghai
(College of Agronomy and Plant Protection, Qingdao Agricultural University; Key Laboratory of Integrated Crop Pest Management of Shandong Province, Qingdao266109, China)
The parasitism of the soybean population (SCNS) and tobacco population (SCNT) ofHeteroderaglycines(soybean cyst nematode, SCN) was tested separately on five soybean cultivars (SCN host differentials) and nine tobacco cultivars through pot experiments. It was found that, on the five soybean cultivars, the mean reproduction factor (Rf=Pf/Pi) of the SCNSwas 0.60 (0.14-1.90) with a highRfof 1.90 on the control susceptible cultivar ‘Lee’, and the meanRfof the SCNTwas only 0.18 (0.06-0.44) with a meanRfof 0.44 on ‘Lee’. The SCNScould hardly parasitize the nine tobacco cultivars with a meanRfof 0.06 (0.00-0.10), but the SCNTinfected well with the tobacco cultivars with a meanRfof 1.26 (0.45-1.95). In 35 d after the inoculation, the juveniles of SCNSand SCNTwere both rare in the roots of different soybean cultivars with nearly the same infection ratios, but in the roots of different tobacco cultivars, none of SCNSjuveniles was found and the mean juvenile number of SCNTwas 26.5 (14.6-57.0). The results indicated that the SCNSwas the physiologic race No.14, and SCNTwas a new SCN pathogenic type which greatly differed in parasitism from SCNS.
Heteroderaglycines;Glycinesmax;Nicotianatabacum;reproduction factor;parasitism;new pathogenic type
2015-03-08
2015-03-27
公益性行业(农业)科研专项(201503114);中国烟草总公司科技项目(110200902065);山东省“泰山学者”建设工程专项;山东省普通本科高校应用型人才培养专业发展支持计划项目
E-mail:hhzhao@qau.edu.cn
S 435. 651
A
10.3969/j.issn.0529-1542.2016.02.011
致谢:沈阳农业大学北方线虫研究所段玉玺教授提供大豆孢囊线虫鉴别寄主种子,中国农业科学院烟草研究所提供烟草品种,在此表示感谢。
