边坡先锋植物木豆的抗旱性
2016-09-08许翩翩王建柱
许翩翩, 王建柱
(三峡大学生物与制药学院,湖北宜昌 443002)
边坡先锋植物木豆的抗旱性
许翩翩, 王建柱*
(三峡大学生物与制药学院,湖北宜昌 443002)
[目的]探究干旱胁迫对木豆光合及生理生化指标的影响,为水电边坡生态修复中的物种筛选、植被群落的重建提供参考。[方法]采用盆栽控水法进行干旱胁迫,测定木豆叶片净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、水分利用效率(Pn/Tr)、丙二醛(MDA)含量、脯氨酸(Pro)含量和可溶性糖含量8个抗旱指标的变化。[结果]木豆幼苗受水分变化的影响显著,在干旱胁迫下Pn、Gs、Tr、Ci、Pn/Tr等光合指标均显著下降,表现出不利的负向生理生态学响应;木豆叶片MDA、Pro和可溶性糖含量均显著增加以有效应对干旱胁迫。[结论]木豆的抗旱能力较强,适应于干热地区的生态环境,是水电边坡治理与开发的优选灌木。
水电工程;干旱胁迫;光合特性;生理特性;木豆
出于对清洁能源的巨大需求,我国目前在金沙江的干热河谷地区规划或建设白鹤滩、乌东德等一系列水电工程[1]。这些水电工程所在地位于亚热带季风气候区,具有云南高原气候向四川盆地气候过渡的气候特点,兼具干旱河谷的生态特征[2]。调查发现大型水电工程扰动区的生态环境十分恶劣,因大规模的开挖,其土壤物理结构、土壤肥力、地下水路径、地表覆盖都受到严重破坏,植被的生态恢复难度较大[3-4]。一些抗逆性强的植物由于能在短时间内形成植物群落,起到稳固坡地、保持水土、改善生态环境的作用[5-7],常被用作生态恢复工程中的先锋物种。缺水和土地贫瘠是边坡生态修复的限制问题,尤其是针对西南干热河谷地区,因此,科学筛选抗旱植物是水电边坡生态修复成功的关键和前提。植物的抗旱性是其在干旱环境下生长和繁殖的必要条件,是多种因素综合作用的结果[8],虽然目前有众多的研究成果,但不同植物的抗旱机制不同,植物的自身形态和生理生化特征都与植物抗旱性有关[9],对于干热河谷地区边坡生态修复植物,目前尚不能全面准确地描述各种植物的抗旱特征。
木豆(Cajanuscajan),又称豆蓉、扭豆,为豆科木豆属灌木,株高可达3 m。木豆主要分布于我国云南、四川、广东、广西,热带和亚热带地区广为栽培。木豆喜温暖,耐干旱贫瘠,易繁殖,易于种植在乡村及边坡空地,既可增加生产,又可改良土壤,可作为先锋恢复物种应用到水电边坡的生态恢复工程中。水分是重要的生态因子,植物的光合及水分生理特性可反映其生长状况及其与生境的协调适应情况[10-11],可为研判先锋群落的稳定性和群落演替提供有益的信息[12]。鉴于此,笔者通过模拟水电边坡不同水分特征,探究了木豆在干旱胁迫下的光合及生化指标变化,揭示了木豆的耐旱机理,并结合植株形态学变化特征对其适生性进行了综合评价,以期为水电边坡的先锋植物筛选、植被恢复建设提供技术和理论支撑。
1 材料与方法
1.1材料试验地点为湖北省宜昌市三峡大学的生态学试验中心(111 18′64″ E、30°43′44″ N),试验所用木豆种子采购于四川省成都苗木基地。
1.2试验设计挑选出籽粒饱满、大小均匀且无损伤的种子,采用盆栽方式在恒温透明的塑料温棚进行试验。供试土壤为边坡典型黄棕壤,粉碎混匀后装入盆中,每盆播种10粒种子,播种40盆。定期浇水,予以正常管理,培养一段时间后,每盆留取生长良好且高度相近的2株植株幼苗(苗长约20 cm)用于后续试验。将40盆幼苗随机分为4组进行4个水平处理。其中,CK组为对照组,土壤含水量为田间持水量的75%~80%,T1、T2、T3三组为干旱胁迫处理,分别控制土壤含水量为田间持水量的60%~75%(轻度干旱胁迫)、50%~55%(中度干旱胁迫)、35%~40%(重度干旱胁迫)。采取称重法控制土壤含水量,每天定时对盆栽进行称重、补水直至形成干旱胁迫水分梯度。保持设置的水分梯度15 d后,测定幼苗叶片的各光合参数值及相应的生理生化指标。
1.3测定方法采用Li-6400便携式光合分析仪分别测定木豆叶片Pn、Gs、Tr、Ci,并计算植物Pn/Tr。选取植株顶部从上到下的第3或第4片完全展开的健康成熟叶片,使用Li-6400便携式光合分析仪的2 cm×3 cm标准红蓝光源叶室测定各处理组植株叶片的Pn、Tr、Gs、Ci等各项光合参数,每个处理重复测定10株植物。测定前,先用饱和光对植物进行30 min的光诱导,测定时将叶室温度设为25 ℃,饱和光强控制在1 000 μmol/(m2·s),CO2浓度控制在400 μmol/mol,所有测定均于9:00~11:00在室外25 ℃的环境下完成。生理生化指标的测定:采用硫代巴比妥酸(TBA)法测定各处理水平的丙二醛含量;采用茚三酮法测定各处理水平的脯氨酸含量;采用蒽酮法测定各处理水平的可溶性糖含量。每一指标的测定均重复3次,且每一处理水平随机选取5株与光合作用测定位置和成熟度近似的新鲜叶片进行测定。
1.4数据处理采用单因素方差分析法分析木豆种子光合及生理生化指标在不同干旱胁迫条件下的差异性,并进行LSD检验比较各组间差异显著性,揭示木豆的光合作用对土壤干旱胁迫的响应,并分析土壤干旱胁迫下木豆生理生化特征的变化规律。所有统计分析采用SPSS 17.0和Excel 2003软件进行。
2 结果与分析
2.1土壤干旱胁迫对木豆光合特性的影响
2.1.1木豆叶片Pn的变化。木豆叶片的Pn随干旱胁迫程度的加剧逐渐下降,干旱胁迫越剧烈,木豆叶片Pn降低的幅度越大(图1)。在正常供水条件下,木豆叶片的Pn值为14.36 μmol/(m2·s);在T1、T2和T3组Pn值显著降低且与CK组存在显著差异,分别较CK组下降了39.55%、69.99%和79.39%。

注:柱上不同字母表示不同处理间在0.05水平差异显著。 Note:Different letters indicated significant differences at 0.05 level between treatments. 图1 不同干旱胁迫条件下木豆叶片Pn的变化Fig.1 Changes of Pn of C. cajan under different drought stresses
2.1.2木豆叶片Gs的变化。木豆在不同干旱胁迫条件下叶片的Gs均显著降低,但T2和T3组差异不显著(图2)。正常土壤水分条件下木豆的叶片Gs为0.321 mol/(m2·s),Gs随着土壤含水量的减少而明显下降,T1、T2和T3分别较CK下降了17.45%、48.60%和44.50%。不同干旱条件均使木豆气孔活动程度明显降低,以避免自身营养物质过多损失。
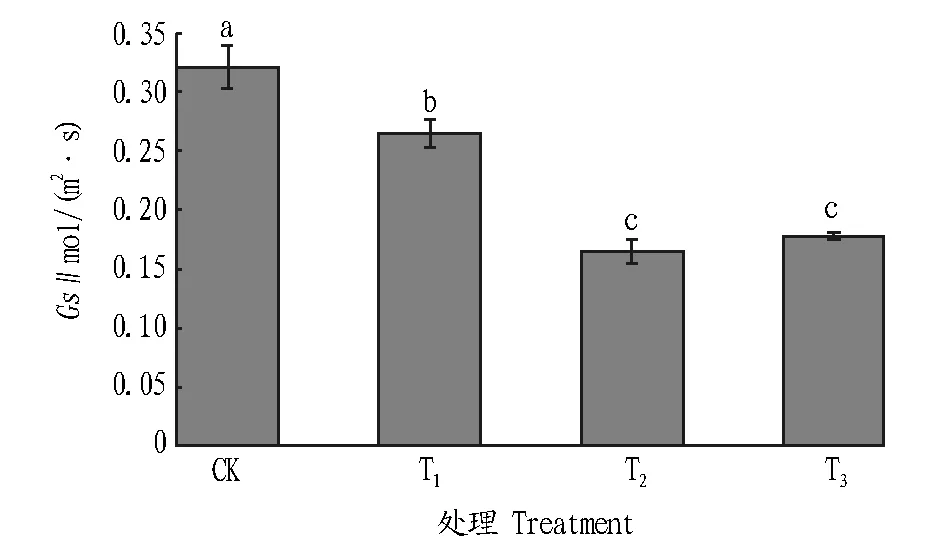
注:柱上不同字母表示不同处理间在0.05水平差异显著。 Note:Different letters indicated significant differences at 0.05 level between treatments.图2 不同干旱胁迫条件下木豆叶片Gs的变化Fig.2Changes of Gs of C. cajan under different drought stresses
2.1.3木豆叶片Tr的变化。随着土壤干旱胁迫程度的加剧,木豆叶片的Tr呈显著下降趋势,T2和T3组Tr差异不显著。当土壤干旱胁程度增加,为有效减少叶片失水,木豆的叶片Tr均出现不同程度的下降,与Gs的变化趋势基本一致。CK组叶片Tr是4.45 mmol/(m2·s);T1、T2和T3组叶片的Tr分别为3.32、2.05、1.76 mmol/(m2·s)。随着干旱胁迫程度的加重,木豆Tr有较大幅度的下降,但不同处理间Tr下降趋势有差异,其中T3组木豆叶片Tr较CK下降了60.45%(图3)。
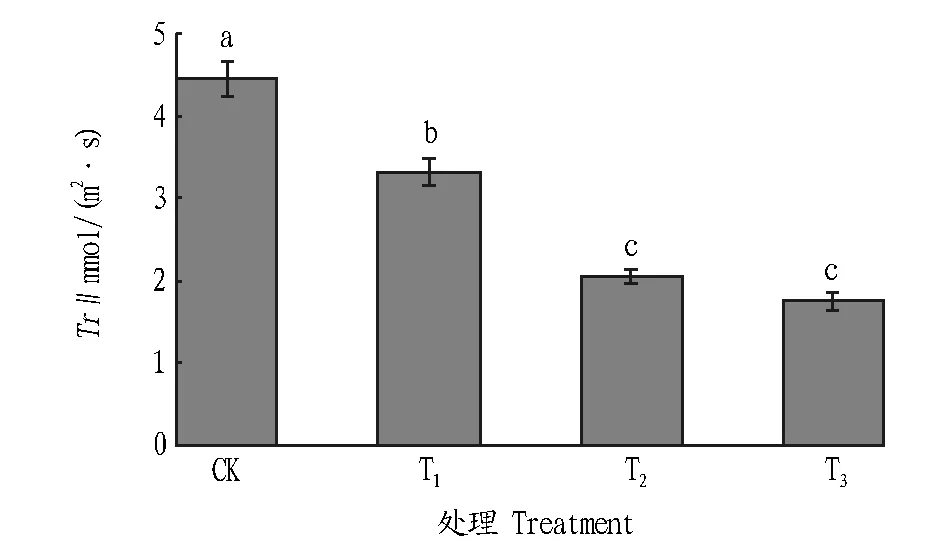
注:柱上不同字母表示不同处理间在0.05水平差异显著。 Note:Different letters indicated significant differences at 0.05 level between treatments.图3 不同干旱胁迫条件下木豆叶片Tr的变化Fig.3 Changes of Tr of C. cajan under different drought stresses
2.1.4木豆叶片Ci的变化。T1、T2组Ci随着土壤干旱胁迫程度的加剧而显著下降,T3组Ci较T2组显著上升,且与T1组无显著性差异。说明T1、T2组木豆光合作用可能受光合作用限制,而T3组非气孔限制占主导。CK组Ci值为246.32 μmol/(m2·s);T1、T2和T3组木豆叶片Ci值分别是212.45、185.83、208.95 μmol/(m2·s)。木豆Ci随着干旱胁迫的加剧呈先下降后上升的趋势,T1、T2组明显下降,在T3组Ci值又明显上升,T1、T2和T3组木豆Ci较CK分别下降了13.75%、24.56%和15.17%(图4)。
2.1.5干旱胁迫对木豆水分利用效率的影响。木豆叶片受干旱胁迫其水分利用效率均显著降低。随着土壤含水量的降低木豆叶片水分利用效率逐渐减小(图5)。在正常土壤水分条件下,木豆叶片水分利用效率为4.15 μmol/mmol,随着干旱胁迫的加重,木豆叶片水分利用效率在各干旱胁迫处理时均明显下降,T1、T2和T3分别较CK下降了27.23%、52.30%和72.77%。

注:柱上不同字母表示不同处理间在0.05水平差异显著。 Note:Different letters indicated significant differences at 0.05 level between treatments.图4 不同干旱胁迫条件下木豆叶片Ci的变化Fig.4 Changes of Ci of C. cajan under different drought stresses

注:柱上不同字母表示不同处理间在0.05水平差异显著。 Note:Different letters indicated significant differences at 0.05 level between treatments.图5 不同干旱胁迫条件下木豆叶片水分利用效率的变化Fig.5 Changes of water use efficiency of C. cajan under different drought stresses
2.2土壤干旱胁迫对木豆生理生化特性的影响木豆叶片在T1、T2和T3组MDA含量与CK组差异显著。随着土壤干旱胁迫程度的加大,木豆叶片的MDA含量逐渐上升。在CK组木豆叶片的MDA含量为17.28 mmol/g,木豆叶片在T1、T2和T3组MDA含量分别为CK组的1.20倍、1.47倍和2.02倍。当土壤含水量逐渐减小时,木豆细胞质膜过氧化反应程度加大,细胞膜受损程度不断加深,严重干旱使得木豆植物严重受损,使其耐旱效果较差。
随着干旱胁迫的增加,木豆叶片Pro含量显著上升,而T2和T3组Pro含量差异不显著。CK组木豆叶片Pro含量为31.54 μg/g,木豆叶片Pro含量在各胁迫处理中均远高于CK组,而中度和重度干旱处理(T1、T2)间Pro含量相当,在T1、T2和T3组Pro含量分别是CK组的2.23倍、5.81倍和6.03倍。在T1和T2组,木豆叶片内Pro含量显著上升来调节植物渗透平衡,而T3组叶片Pro增加减缓其渗透调节能力降低,对干旱环境适应性减弱。
木豆叶片在不同干旱胁迫下的可溶性糖含量均呈显著上升趋势。CK组木豆叶片可溶性糖含量为2.38 mg/g,T1、T2和T3组木豆叶片积累的可溶性糖含量分别是对照的1.67倍、2.29倍和3.10倍。通过对可溶性糖含量积累程度比较,发现不同程度的干旱胁迫均使木豆叶片可溶性糖含量增加;中度和重度干旱胁迫会导致木豆可溶性糖含量显著升高。通过不同程度上可溶性糖的积累,可实现体内渗透调节的平衡(表1)。
3 结论与讨论
在诸多环境因素当中,水分是影响植物生长发育和生存最重要因子之一。水分利用效率是植物物质生产和水分消耗之间的重要综合指标,不仅反映植物生理功能及其对环境的适应性,也是植物对水分条件响应的基础研究指标[13],蒸腾速率低、光合速率和水分利用效率高是抗旱物种的特性之一[14]。在不同的干旱胁迫条件下,木豆的净光合速率、气孔导度、蒸腾速率等光合指标值均呈现出不同的下降趋势。净光合速率是植物光合作用强弱的直接指标,其高低反映了植物积累营养物质能力的大小,正常的水分条件有助于木豆的生长发育,水分不足会影响木豆净光合产物的积累,且在T2和T3组更为显著,木豆叶片的水分利用效率变化趋势与净光合速率基本一致。木豆的气孔导度和蒸腾速率随干旱胁迫的增加均减小,且T2和T3组无显著性差异。木豆的生理代谢活动因氧气的减少而降低,从而造成气孔导度和蒸腾速率均明显下降。在T1水平植株为阻止水分过度散失降低了气孔导度水平使得蒸腾速率降低,在T2和T3水平则可能是非气孔限制导致气孔导度和蒸腾速率下降且两水平无显著差异。胞间CO2浓度由CK到T2呈降低趋势,但到T3增加,且T1和T3无显著性差异。有研究认为,如果气孔因素限制占主导,那么植物净光合速率的下降伴随着胞间CO2浓度的下降,如果非气孔因素限制占主导,那么植物净光合速率的下降就会伴随着胞间CO2浓度的上升[15]。由CK到T2组由于气孔导度降低致使环境中CO2浓度进入叶肉细胞减少,从而使胞间CO2浓度降低进而使净光合速度降低。而T3组水分胁迫造成叶肉细胞光合活性的降低,从而导致光合作用利用的CO2量减少,而呼吸作用产生的CO2量不变,所以使胞间CO2增加,而净光合速度始终呈现下降的趋势。
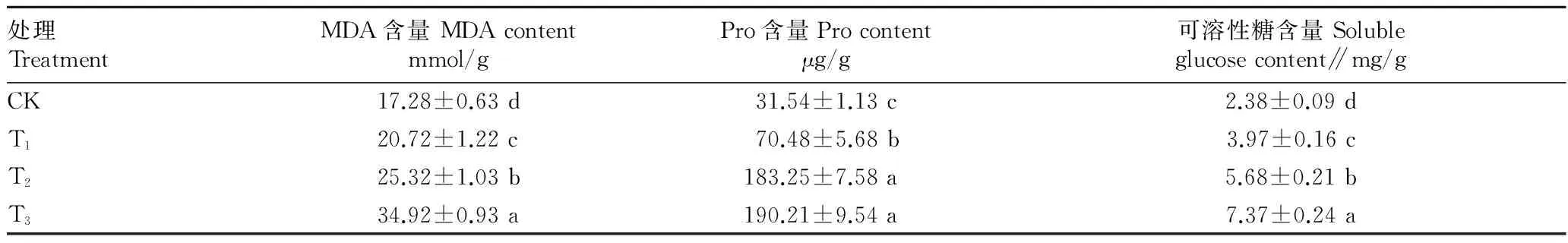
表1 不同干旱胁迫条件下木豆叶片生理生化指标变化
注:同列数据后不同字母表示不同处理间在0.05水平差异显著。
Note:Different letters indicated significant differences at 0.05 level between treatments.
丙二醛是植物细胞膜脂质过氧化作用的产物,当植株处于干旱胁迫条件下时,植株体内丙二醛含量就会增加,丙二醛含量的积累能破坏细胞膜的结构和功能,引起细胞膜功能紊乱,其含量的高低可以反映植株生理代谢活动的强弱和植株遭受伤害的程度[16]。木豆在各干旱胁迫下叶片丙二醛积累含量都显著增加,膜系统受伤害程度不断加深,但在T1、T2水平增加幅度不大,在T3水平则显著增加,表明木豆在一定干旱情况下膜脂过氧化程度变动不大,具备一定的抗旱性。脯氨酸是一种调节渗透的物质,许多植物能通过渗透调节物质来维持细胞一定的含水量和膨压,从而维持细胞的正常生理功能[17]。研究表明,水分胁迫条件下植物渗透调节物质可溶性糖的增加可以降低植物体内的渗透势,有利于植物体在干旱逆境中维持体内正常的所需水分,提高植物的抗逆适应性[18]。 随着干旱胁迫的增加木豆叶片脯氨酸含量和可溶性糖均显著上升,这与李渡等[19]对苜蓿的研究结果一致。干旱胁迫条件下,木豆叶片内游离脯氨酸和可溶性糖含量的大量积累是进行渗透调节的一种反应,以维持细胞一定的含水量和膨压势,提高其自身抗逆适应性。
通过对木豆光合及生理生化指标的分析发现其幼苗具有一定的抗旱能力,在轻度和中度干旱胁迫条件下,各项生化指标反应良好,表现出较强的干旱适应特征,适合栽种于西南干热河谷地区,可作为水电边坡生态修复的备选物种。但因地质环境的差异及物种适应性等因素还需结合其他物种进一步对比研究。
[1] 王传华,曾春函,沈德嵩,等.云南干热河谷水库气候效应对车桑子幼苗生长发育的影响及其作用机制[J].生态学报,2016,36(17):1-10.
[2] 中国水电顾问集团中南勘测设计研究院. 金沙江向家坝水电站水土保持方案报告书[Z].2006.
[3] 曹永强,倪广恒,胡和平.水利水电工程建设对生态环境的影响分析[J].人民黄河,2005,27(1):56-58.
[4] 董哲仁.水利工程对生态系统的胁迫[J].水利水电技术,2003,34(7):1-5.
[5] 杨修,高林.德兴铜矿矿山废弃地植被恢复与重建研究[J].生态学报,2001,21(11):1932-1941.
[6] 李西,罗承德,陈其兵.岩石边坡植被护坡植物选择初探[J].中国园林,2004(9):52-53.
[7] ZHOU Z C,SHANGGUAN Z P.Effect of ryegrasses on soil runoff and sediment control[J].Pedosphere,2008,18(1):131-136.
[8] 孙彩霞,沈秀瑛.作物抗旱性鉴定指标及数量分析方法的研究进展[J].中国农学通报,2002,18(1):49-51.
[9] 庄丽,陈亚宁,陈明,等.模糊隶属法在塔里木河荒漠植物抗旱性评价中的应用[J].干旱区地理,2005,28(3):367-372.
[10] 蔡锡安,孙谷畴,赵平,等.土壤水分对单性木兰幼苗光合特性的影响[J].热带亚热带植物学报,2004,12(3):207-212.
[11] ZHANG S Y,XIA J B,ZHOU Z F,et al.Photosynthesis responses to various soil moisture in leaves ofWisteriasinensis[J].Journal of forestry research,2007,18(3):217-220.
[12] 赵平.退化生态系统植被恢复的生理生态学研究进展[J].应用生态学报,2003,14(11):2031-2036.
[13] 蒋高明,常杰,高玉葆,等.植物生理生态学[M].北京:高等教育出版社,2004:9-57.
[14] 熊伟,王彦辉,于澎涛.树木水分利用效率研究综述[J].生态学杂志,2005,24(4):417-421.
[15] FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].Annual review of plant physiology,1982,18(2):1-3.
[16] 汤章城.植物对水分胁迫的反应和适应性、抗逆性的一般概念和植物的抗涝性[J].植物生理学通讯,1983(3):24-29.
[17] 丁玉梅,马龙海,周晓罡,等.干旱胁迫下马铃薯叶片脯氨酸、 丙二醛含量变化及与耐旱性的相关性分析[J].西南农业学报,2013,26(1):106-110.
[18] 柴成林,李绍华,徐迎春.水分胁迫期间及胁迫解除后桃树叶片中的碳水化合物代谢[J].植物生理学通讯,2001,37(6):495-498.
[19] 李渡,贾秀峰,白庆武,等.干旱胁迫对苜蓿脯氨酸积累的影响[J].植物研究,2003,23(2):189-191.
Drought Resistance Research for Slope Pioneer PlantCajanuscajan
XU Pian-pian,WANG Jian-zhu*
(College of Biopharmaceutis,China Three Gorges University,Yichang,Hubei 443002)
[Objective] To explore the effects of drought stress on photosynthesis,physiological and biochemical indices ofCajanuscajan,and to provide scientific references for species screening and community reconstruction in ecological restoration of hydropower slope. [Method] Pot water control method was used for drought stress ofC.cajan. Changes of net photosynthetic rate (Pn),stomatal conductance (Gs),transpiration rate (Tr),intercellular CO2concentration (Ci),water use efficiency (Pn/Tr),malondialdehyde (MDA),proline (Pro) and soluble glucose content were detected inC.cajan. [Result]C.cajanseedlings were greatly affected by the moisture change. Under drought stress,thePn,Gs,Tr,CiandPn/Trall significantly decreased,showing a negative photosynthetic response to the drought stress. MDA,Pro and soluble glucose content significantly increased,so as to effectively respond to drought stress. [Conclusion]C.cajanhas strong draught tolerance,can adapt to the ecological environment of dry-heat area. It is the optimal shrub to control degraded mountain land.
Hydropower engineering; Drought stress; Photosynthetic characteristic; Physiological characteristics;C.cajan
国家自然科学基金项目(51179094)。
许翩翩(1987-),女,湖北松滋人,硕士研究生,研究方向:恢复生态学。*通讯作者,副教授,博士,从事生态学研究。
2016-05-23
Q 945.78
A
0517-6611(2016)20-001-03
