甘蓝型油菜分枝角度主基因+多基因混合遗传模型及遗传效应
2016-08-27汪文祥梅德圣李云昌周日金成洪涛付
汪文祥 胡 琼 梅德圣 李云昌 周日金 王 会 成洪涛付 丽 刘 佳
中国农业科学院油料作物研究所 / 农业部油料作物生物学与遗传育种重点实验室, 湖北武汉430062
甘蓝型油菜分枝角度主基因+多基因混合遗传模型及遗传效应
汪文祥 胡 琼 梅德圣 李云昌 周日金 王 会 成洪涛付 丽 刘 佳*
中国农业科学院油料作物研究所 / 农业部油料作物生物学与遗传育种重点实验室, 湖北武汉430062
分枝角度是油菜株型重要性状, 是油菜品种高产及适合机械化收获理想株型的基本组成之一。为明确油菜分枝角度的遗传, 本研究选用油菜分枝角度大的松散型材料6098B和分枝角度小的紧凑型材料Purler配制杂交组合,采用主基因+多基因混合遗传模型方法对该组合6世代(P1、P2、F1、F2、BCP1和BCP2)的分枝角度进行了遗传分析。结果表明, 上部第一分枝(顶枝)和基部第一分枝(基枝)角度的最适合遗传模型均为 D-0 (1对加性-显性主基因+加性-显性-上位性多基因)。顶枝角的主基因加性效应值为4.939º, 显性效应值为-4.156º, 主基因遗传率在BCP1、BCP2和F2中分别是 34.08%、1.40%和14.99%, 多基因遗传率分别为24.43%、61.72%和 63.98%; 而基枝角的主基因加性效应值为 2.217º, 显性效应值为-1.941º, 主基因遗传率在BCP1、BCP2和 F2中分别是 7.86%、1.24%和4.84%, 多基因遗传率分别为66.46%、58.49%和73.96%。结果发现油菜分枝角度明显存在主效基因, 为油菜分枝角度的遗传改良奠定了基础。
油菜; 分枝角度; 理想株型; 主基因+多基因; 遗传效应
油菜是我国种植面积最大的油料作物, 年种植面积约 730万公顷(2015中国统计年鉴)。油菜籽不仅仅用来榨油, 菜籽饼粕也是重要的工业原料, 并与畜牧和水产养殖业发展高度关联。近年来,油料供给短缺的状态一直未得到有效解决, 甚至日趋紧张[1]。单产水平低和机械化生产普及率低是限制我国油菜产业发展的主要瓶颈。培育适合高效机械化生产油菜品种, 构建油菜理想株型, 改良油菜在密植环境下的生长发育情况, 是提高油菜产量的基础, 也是促进机械化收获提高油菜生产效益的关键。
株高、分枝部位高、分枝数和主花序有效长是油菜株型重要的组成部分, 相关的研究较多[2-7], 但关于油菜分枝角度的研究较少。分枝角度主要指有效分枝与主茎之间所成的夹角。分枝角度大, 植株松散, 增加了单个植株的占地面积, 降低了叶面积指数及单位面积上光合作用面积, 相应降低了产量;植株间相互交织, 影响光能透射和通风, 不仅因降低植株中下部绿色组织的光能利用率而降低群体产量, 同时增加了病害的风险及机械化收割时刮碰造成的产量损失。减小分枝角度对增加油菜种植密度,实现高产高效生产意义重大。
油菜分枝角度的遗传研究, 对培育紧凑株型油菜品种具有重要理论指导作用。前人的研究基本局限于分枝角度与产量性状的相关性[8-9], 而对油菜分枝角度基因的效应大小及其作用形式很少报道。张文学等[10]首次研究了甘蓝型油菜紧凑型和松散型杂种一代分枝性状的遗传效应, 结果表明紧凑型与松散型正反交F1代分枝角度无差异, 表现双亲平均值,说明细胞质效应不影响分枝角度。王俊生等[11]利用6个油菜品种构建 30个完全双列杂交组合, 对分枝角度、一次分枝数、分枝长等10多个株型性状进行了遗传效应及其与产量性状的相关性分析, 结果表明分枝角度狭义遗传率较高, 显性效应对分枝角度有减效作用, 并与单株产量特别是单株角果数存在显著的遗传负相关。近几年, 为构建机械化密植条件下的油菜理想株型, 分枝角度的研究也逐渐得到重视[12-13], Liu等[14]利用油菜自然群体, 定位了6个与分枝角度显著关联的基因组区域, 并获得其中 4个区域内候选基因。但迄今为止仍缺少利用遗传群体对该性状进行传统遗传研究的报道。
主基因+多基因混合遗传模型是由盖钧镒[15]提出的数量性状遗传分析方法, 在多种植物的遗传研究中得到应用[16], 在油菜性状的遗传研究中也广泛应用[9,17-21]。本文以6个世代的研究群体(P1、P2、F1、F2、BCP1和 BCP2), 利用数字图像采集法鉴定油菜上部第一分枝角(顶枝角)和基部第一分枝角(基枝角), 运用主基因+多基因混合遗传模型分析分枝角度的遗传模式和遗传效应, 评估单个主基因不同的遗传效应和各分离世代的遗传率, 揭示分枝角度的遗传规律。研究结果不仅为改良油菜株型性状提供理论指导, 也为进一步QTL定位和相关基因的克隆奠定基础。
1 材料与方法
1.1 试验材料
亲本材料分别为6098B (P1)和Purler (P2), 6098B是中国农业科学院油料作物研究所杂种优势利用课题组选育, 属于松散型; Purler是引自澳大利亚的油菜品种, 属于紧凑型。2013年秋, 在中国农业科学院油料作物研究所阳逻试验基地种植亲本, 2014年春手工去雄授粉杂交获得 F1(P1×P2)种子; 同年, 将亲本P1、P2和F1种植于青海大学试验基地, 将F1套袋自交收获F2种子, 再以2个亲本分别与F1回交获得BCP1(6098B/Purler//6098B)和 BCP2(6098B/Purler// Purler)。从而获得6098B/Purler杂交组合的6个基本世代群体, 即P1、P2、F1、F2、BCP1和BCP2群体。亲本材料由中国农业科学院油料作物研究所油菜杂种优势利用课题组保存提供。
1.2 田间试验
2014年9月, 将6个世代群体种子同期播种在中国农业科学院油料作物研究所阳逻试验基地, 行距33 cm, 株距11 cm, 每行15株, 田间管理与普通大田生产基本相同。种植P1、P2和F1群体各6行, F2群体18行, BCP1和BCP2群体各12行。试验地肥力均匀一致, 参照当地大田栽培的施肥水平。2015年5月8日避开行两端的边株分单株收获群体待表型考察。
1.3 分枝角度鉴定方法
2015年6月在室内采用数字图像处理法[22]鉴定6世代群体分枝角度: 油菜成熟后, 剪取连有油菜上部第一分枝(顶枝)和基部第一分枝(基枝)的茎段, 采集图像。将该图像文件导入AutoCAD软件, 利用角度工具标注夹角角度, 将分枝角度结果记录到Excel文档中。
1.4 数据统计分析
利用SAS 9.3统计分析软件进行基本统计量、相关系数及多组数据差异显著性比较。利用植物数量性状主基因+多基因混合遗传模型分析方法[15],并结合软件[23]数据处理进行遗传模型分析(分析软件由南京农业大学章元明教授提供)。通过极大似然分析和 IECM (iterated expectation and conditional maximization)算法对混合分布中的相关分布参数作出估计, 再采用AIC (Akaike’s information criterion)值选择最佳模型,并进行适合性检验, 包括均匀性和检验, Smirnov检验(nW2)和Kolmogorov检验(Dn), 根据检验结果选择最优遗传模型。最后采用最小二乘法依据最优模型的各成分分布参数估计各基因效应值、方差等遗传参数。群体的多基因方差100%, 多基因遗传率,其中和σ2分别表示主基因方差、多基因方差、表型方差和环境方差。
2 结果与分析
2.1 表型数据分析
由表1可知, 亲本P1(6098B)的顶枝角和基枝角分别为71.7°±5.8°和50.6°±6.8°; 亲本P2(Purler)的顶枝角度和基枝角分别为26.2°±4.5°和15.4°±2.7°; 2个亲本 2个部位的分枝角间差异均达到显著水平(P<0.05)。F1的顶枝角和基枝角均值分别为 41.6°±6.2° 和 38.0°±5.4°, 介于双亲之间, 略偏向高值亲本。F2群体的顶枝角和基枝角均值分别为 39.2º±10.9º和34.5º±8.9º, 其分布为偏态分布, 表明分枝角度受到多基因的控制。F1与亲本(P1)回交的BCP1群体的顶枝角和基枝角均值分别为 47.0°±7.8°和 41.0°±8.1°,偏向于P1亲本(图1)。F1与亲本(P2)回交的BCP2群体的顶枝角度和基枝角均值分别为 32.2°±8.2°和26.4°±7.2°, 偏向于亲本P2(图2)。利用所有单株进行统计分析, 结果表明顶枝角与基枝角的相关系数为0.611 (P < 0.01, n = 419)。

表1 6098B×Purler 各世代分枝角度统计及变异系数Table 1 Performance of branch angle in six generations from 6098B×Purler in B. napus
2.2 分枝角度主基因+多基因遗传分析
2.2.1 遗传模型的选择 利用植物数量性状主基因+多基因混合遗传模型的6世代(P1、P2、F1、BCP1、BCP2和F2)联合分析, 估算获得1对主基因(A类模型)、2对主基因(B类模型)、多基因(C类模型)、1对主基因+多基因(D类模型)、2对主基因+多基因(E类模型)共5类 24种遗传模型的极大似然函数值和AIC值(表2)。根据遗传模型的选取原则, 即AIC值最小准则, 选取AIC值最小及与最小AIC值比较接近的一组遗传模型作为备选模型。结果显示24种混合模型中, 油菜顶枝角和基枝角的D-0 (1对加性-显性主基因+加性-显性-上位性多基因)和 E-1 (2对加性-显性-上位性主基因+加性-显性多基因)模型的AIC值相对较小, 可作为备选模型。
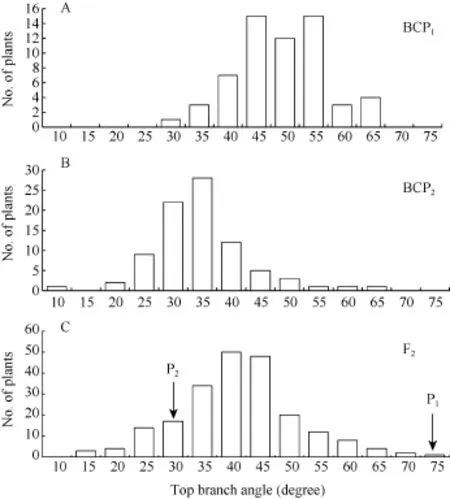
图1 6098B×Purler杂交组合顶枝角的频次分布Fig. 1 Frequency distribution of top branch angle from 6098B×Purler
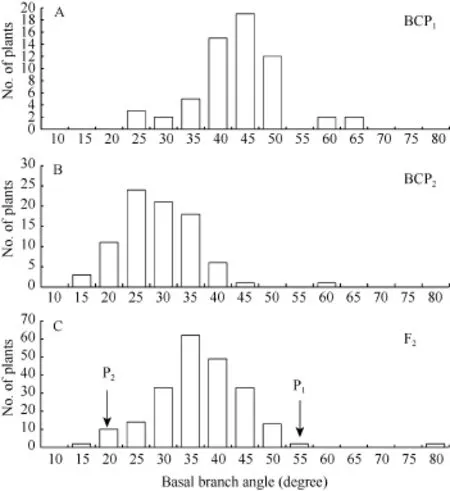
图2 6098B×Purler杂交组合基枝角的频次分布Fig. 2 Frequency distribution of basal branch angle from 6098B×Purler

表2 6098B×Purler组合后代各遗传模型的AIC值Table 2 AIC values of the genetic models from 6098B×Purler
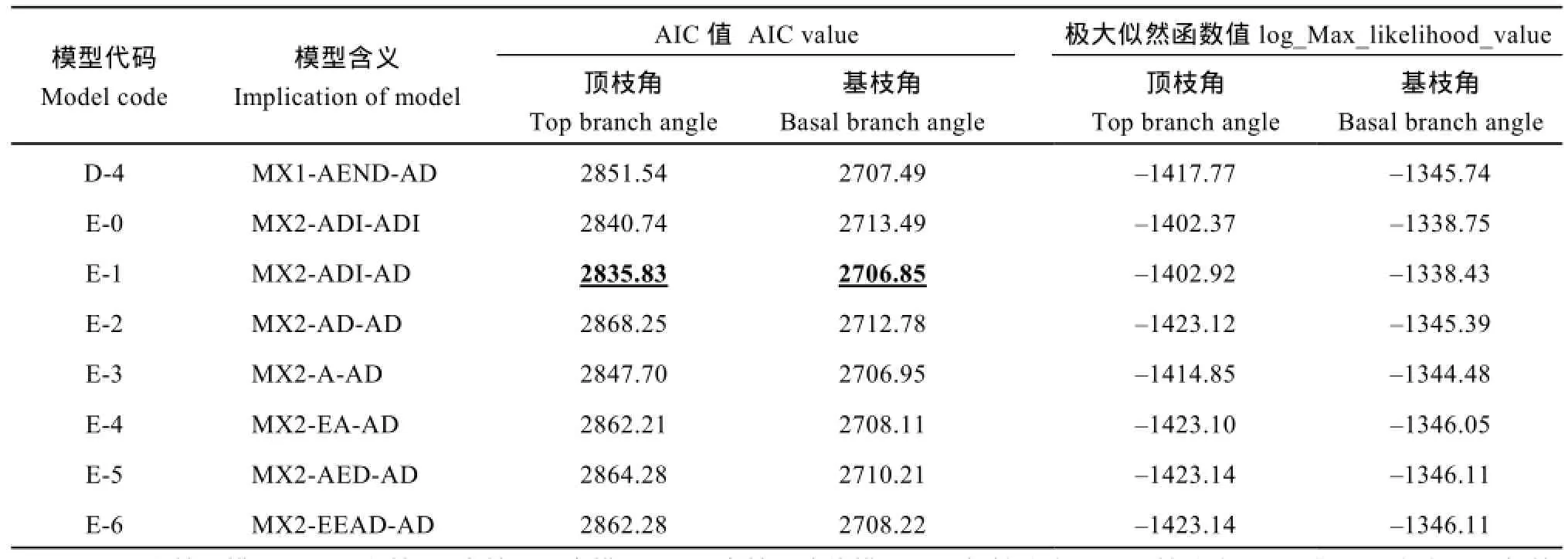
(续表2)
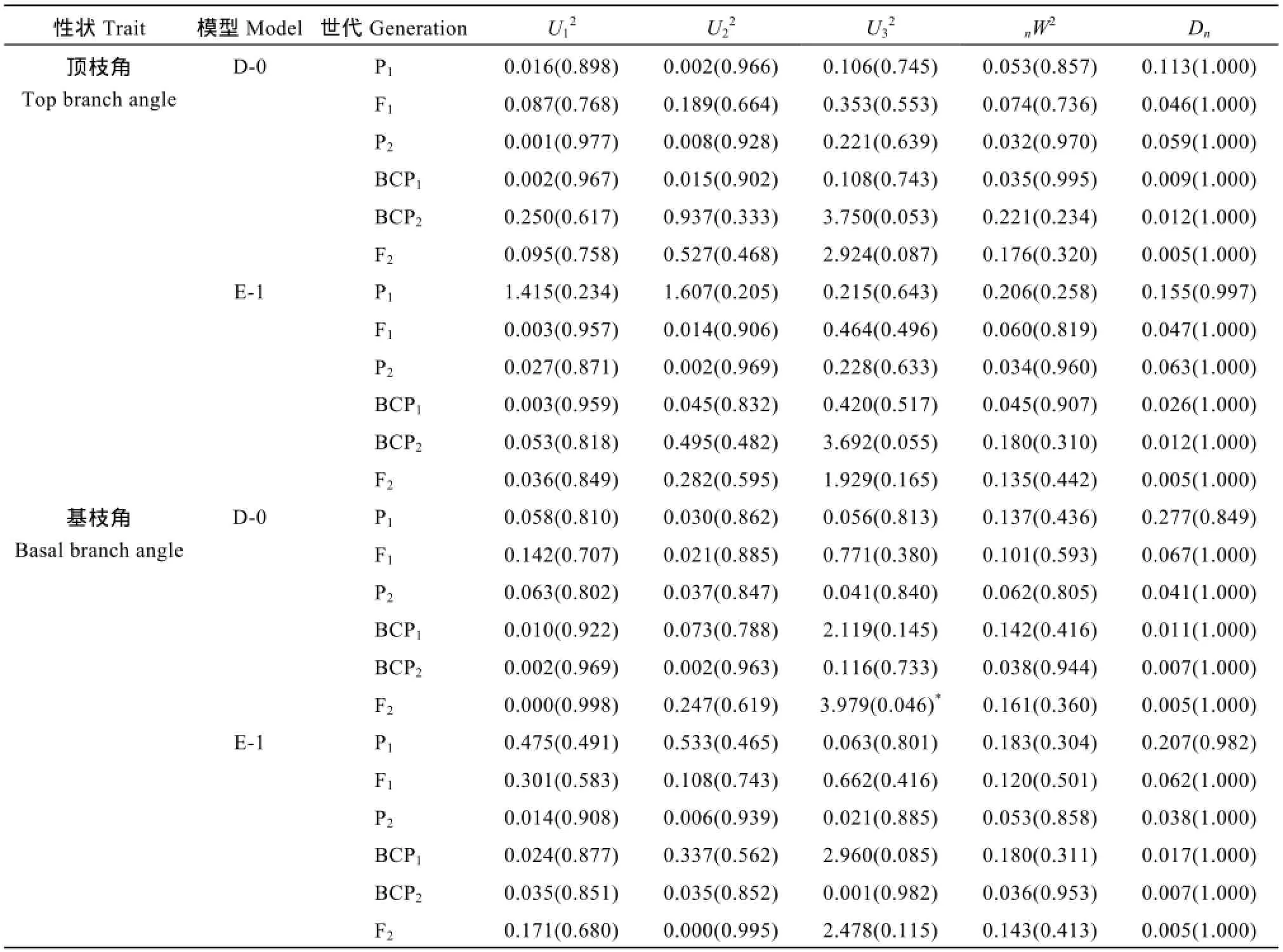
表3 6098B×Purler群体适合性检验Table 3 Fitness tests of selected models in 6098B×Purler
2.2.3 遗传参数估计 根据选择的最优遗传模型估计顶枝角和基枝角的最优遗传模型一阶、二阶遗传参数(表4)。结果表明, 顶枝角和基枝角的主基因加性效应(da)均是正值; 而显性效应(ha)分别为-4.156和-1.941, 均为负值, 主基因显性效应与加性效应的比值均为︱ha/da︱< 1, 即显性效应都小于相应的加性效应, 表明控制分枝角度的1对主基因以加性效应为主。在显性效应的作用方向上, 1对主基因表现为分枝角度大对分枝角度小为部分显性。显性效应增大分枝角度, 表现为部分平生型分枝。
对于油菜顶枝角, BCP1、BCP2和F2分离世代群体遗传率分别为58.51%、63.12%和78.97%, 其中主基因遗传率分别为 34.08%、1.40%和 14.99%, 多基因遗传率分别为24.43%、61.72%和63.98%, 各世代的多基因遗传率都大于相应世代的主基因遗传率,可见油菜分枝角度的遗传体系中, 微效多基因的遗传贡献占主要部分; 3个分离世代群体环境因素引起的变异分别占 11.23%、35.97%和 7.10%; 可见油菜分枝角性状受基因型和环境双重影响。同样, 对于油菜基枝角度, BCP1、BCP2和F2分离世代群体遗传率分别为74.32%、59.73%和78.80%, 其中主基因遗传率分别为 7.86%、1.24%和 4.84%, 多基因遗传率分别为66.46%、58.49%和73.96%, 各世代的多基因遗传率都大于相应世代的主基因遗传率。可见基枝角的遗传体系中, 微效多基因的遗传贡献占主要部分; 3个分离世代群体环境因素引起的变异分别占19.35%、39.54%和17.21%。

表4 6098B×Purler杂交组合分枝角度的一阶遗传参数估计值Table 4 First order genetic parameters estimated for the fitting models of branch angle from 6098B×Purler
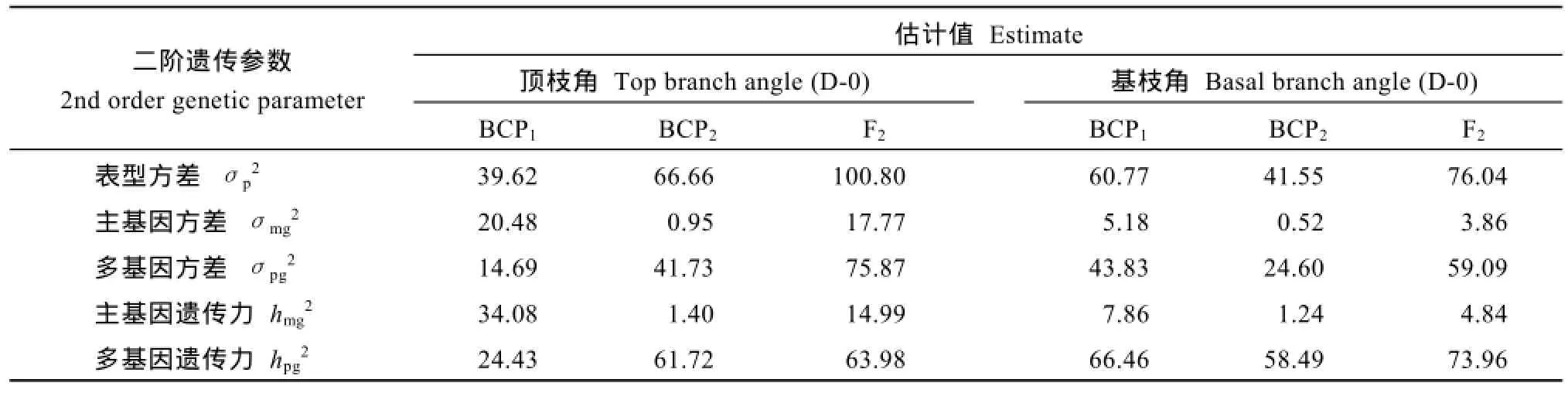
表5 6098B×Purler杂交组合分枝角度的二阶遗传参数估计值Table 5 Second order genetic parameters estimated for the fitting models of branch angle from 6098B×Purler
3 讨论
分枝角度是油菜株型重要的性状, 与产量、耐密性及机械化收获紧密相关。提高种植密度对提高油菜单产和机收适应性都具有非常重要的作用。密植除了增加植株的叶面积指数和光能利用效率外[24],还能提高油菜氮肥利用效率[25], 促进氮素向籽粒转移[26], 起到增产效果。密植还可以提高油菜群体的整齐度, 使茎杆变细、分枝部位提高、分枝长度变短及成熟期相对集中, 有利于机械联合收获, 显著降低机收损失率[27]。密植栽培要求选择株型紧凑、分枝角度较小的品种, 避免植株间相互交织, 有利于光能透射和通风, 提高植株中下部绿色组织的光能利用率, 从而提高群体产量, 也降低了病害的风险。王俊生等[11]的研究发现分枝角度与单株产量特别是单株角果数存在显著的遗传负相关, 说明分枝角度小的植株在生长中后期的物质积累中占有优势,由于中下部光合作用受到的影响较小, 促进了有效分枝数和有效角果数的提高。紧凑型油菜在密植条件下植株间不相互交叉, 可以降低机械收割时刮碰造成的产量损失, 更有利于机械联合收割。因此, 改良油菜株型, 不仅能促进油菜产量的提高也可以推动机械化生产的实施, 符合我国油菜发展的长远目标。
分枝角度形成的直接原因是分枝感受向重力性, 受到植株内部生长素和独角金内酯等激素的调节[28-29]。油菜具多分枝习性, 常规种植密度下(23~30万株 hm-2),油菜的平均分枝数为5~10个。根据田间观察, 油菜植株从下到上的分枝角度一般逐渐加大, 本研究获得的数据证实了这一趋势。从试验结果看, 顶枝和基枝分枝角度相关性极显著, 相关系数为 0.611, 说明顶枝和基枝都能反映油菜品种的分枝角度习性。但顶枝角的遗传效应值(加性效应和显性效应)大于基枝角, 基枝角度因环境引起的变异大于顶枝角度,说明基枝角度受到植株间互相交织和遮蔽的影响更大。因此, 在当前的种植密度下, 顶枝角度更适合作为分枝角度性状指标用于定量遗传分析。
由于顶、基枝角在BCP1、BCP2和F2分离群体中都表现连续正态分布, 选用主基因+多基因模型进行遗传模型和效应分析是恰当的。本研究结果表明, 油菜顶枝角和基枝角均受1对加性-显性主基因+加性-显性-上位性多基因控制(D-0模型), 主基因加性效应使分枝角度增加, 而主基因显性效应使分枝角度减少。其中显性效应为负值与王俊生等[10-11]的显性效应对分枝角度有减效作用的结论相同。主基因遗传率较小, 多基因遗传率较大, 表明油菜分枝角度性状被主基因调控, 但主要受多基因控制,并受环境因素的影响, 与前人研究[10-11]相比, 本研究更进一步获得主基因和微效多基因的遗传效应值,为进一步利用高世代群体克隆相关基因奠定了基础。在选育株型紧凑(分枝角度<30°)油菜品种的过程中需要采用联合选育, 即采用轮回选择或聚合的方法来累积增效基因, 以提高选择效率[30]。同时在油菜杂种优势利用中, 需充分考虑亲本产量性状杂种优势, 同时选取分枝角度均较小的亲本配组, 才能获得杂交种的高产兼顾紧凑株型。
本研究利用6098B和Purler分枝角度差异显著的亲本构建 6个世代遗传群体进行遗传分析, 并且分析样本足够大, 对于油菜分枝角度的鉴定采用数字图像处理分析方法, 从而大大提高结果的准确性[22];利用主基因+多基因混合遗传模型分析油菜分枝角度的遗传, 从研究的油菜组合中检测出 1对主基因的存在与效应, 同时还分析了主基因之外的微效基因的综合表现。下一步需要在不同种植密度环境下,将分枝角度、分枝数、角果数以及单株产量等性状结合分子标记进行遗传定位分析, 从分子水平上阐述分枝角度的基因及其遗传调控网络, 并与产量等相关基因位点作元分析, 发掘密植环境下的与分枝角度及产量相关性状紧密连锁的分子标记, 实现油菜紧凑株型高产稳产的分子标记辅助育种。
4 结论
甘蓝型油菜上分枝角和下分枝角性状均受 1对加性-显性主基因+加性-显性-上位性多基因控制(D-0模型), 显性效应值和加性效应值不同向, 表现为部分显性; 遗传率在 3个分离世代中都表现为中等或者中等偏下、说明在其后代选育相对株型紧凑(分枝角度小)的高产育种品系中, 需要考虑兼顾主基因、多基因和环境因素的影响。References
[1] 王汉中, 殷艳. 我国油料产业形势分析与发展对策建议. 中国油料作物学报, 2014, 36: 414-421
Wang H Z, Yin Y. Analysis and strategy for oil crop industry in China. Chin J Oil Crop Sci, 2014, 36: 414-421 (in Chinese with English abstract)
[2] 唐敏强, 程晓晖, 童超波, 刘越英, 赵传纪, 董彩华, 于景印,马小艮, 黄军艳, 刘胜毅. 甘蓝型油菜株高性状的全基因组关联分析. 作物学报, 2015, 41: 1121-1126
Tang M Q, Cheng X H, Tong C B, Liu Y Y, Zhao C J, Dong C H,Yu J Y, Ma X G, Huang J Y, Liu S Y. Genome-wide association analysis of plant height in rapeseed (Brassica napus). Acta Agron Sin, 2015, 41: 1121-1126 (in Chinese with English abstract)
[3] 王嘉, 荆凌云, 荐红举, 曲存民, 谌利, 李加纳, 刘列钊. 甘蓝型油菜株高、第一分枝高和分枝数的QTL检测及候选基因筛选. 作物学报, 2015, 41: 1027-1038
Wang J, Jing L Y, Jian H J, Qu C M, Chen L, Li J N, Liu L Z. Quantitative trait loci mapping for plant height, the first branch height, and branch number and possible candidate genes screening in Brassica napus L. Acta Agron Sin, 2015, 41: 1027-1038 (in Chinese with English abstract)
[4] 张凤启, 刘越英, 程晓辉, 童超波, 董彩华, 唐敏强, 黄军艳,刘胜毅. 利用高密度SNP标记定位甘蓝型油菜株高QTL. 中国油料作物学报, 2014, 36: 695-700
Zhang F Q, Liu Y Y, Cheng X H, Tong C B, Dong C H, Tang M Q, Huang J Y, Liu S Y. QTL mapping of plant heigh using high desity SNP markers in Brassica napus. Chin J Oil Crop Sci, 2014,36: 695-700 (in Chinese with English abstract)
[5] 谢田田, 陈玉波, 黄吉祥, 张尧锋, 徐爱遐, 陈飞, 倪西源, 赵坚义. 甘蓝型油菜不同发育时期株高 QTL的动态分析. 作物学报, 2012, 38: 1802-1809
Xie T T, Chen Y B, Huang J X, Zhang Y F, Xu A X, Chen F, Ni X Y, Zhao J Y. Dynamic analysis of QTL for plant height of rapeseed at different developmental stages. Acta Agron Sin, 2012, 38:1802-1809 (in Chinese with English abstract)
[6] Xu J S, Song X, Cheng Y, Zou X L, Zeng L, Qiao X, Lu G Y, Fu G P, Qu Z, Zhang X K. Identification of QTLs for branch number in oilseed rape (Brassica napus L.). J Genet Genom, 2014, 41:557-559
[7] Li F, Chen B Y, Xu K, Gao G Z, Yan G X, Qiao J W, Li J, Li H,
Li L X, Zhang T Y, Takeshi Nishio, Wu X M. A genome-wideassociation study of plant height and primary branch number in rapeseed (Brassica napus). Plant Sci, 2015, 242: 169-177
[8] 王俊生, 李少钦, 张耀文. 不同栽培密度对紧凑型油菜产量和主要性状的影响. 耕作与栽培, 2006, (3): 25-26
Wang J S, Li S Q, Zhang Y W. Effects of different planting density on yield and main characters of compact in rapeseed. Gengzuo yu Zaipei, 2006, (3): 25-26 (in Chinese)
[9] 张倩. 甘蓝型油菜主要株型性状的遗传分析和QTL初步定位.西南大学硕士学位论文, 重庆, 2013. pp 3-7
Zhang Q. Genetic Effects Analysis and QTL Mapping of Major Plant-Type Traits in Brassica napus L. MS Thesis of Southwest University, Chongqing, China, 2013. pp 3-7 (in Chinese with English abstract)
[10] 张文学, 李殿荣, 田建华. 甘蓝型油菜紧凑型和松散型杂种一代分枝性状的遗传效应. 中国农学通报, 2000, 16(5): 17-19
Zhang W X, Li D R, Tian J H. Genetic effect of hybrid F1branching character of CCT and LCT in Brassica napus. Chin Agric Sci Bull, 2000, 16(5): 17-19 (in Chinese with English abstract)
[11] 王俊生, 田建华, 张继澍, 李殿荣, 袁志发. 紧凑型油菜株型性状的遗传及其与主要产量性状的相关性研究. 西北农林科技大学学报(自然科学版), 2005, 33: 7-12
Wang J S, Tian J H, Zhang J S, Li D R, Yuan Z F. Study on the heredity of plant-type traits in compact rapeseed lines and the genetic correlation with yield traits. J Northwest Sci-Tech Univ Agric For (Nat Sci Edn), 2005, 33: 7-12 (in Chinese with English abstract)
[12] 唐佳. 甘蓝型油菜株型相关性状的杂种优势及配合力分析.四川农业大学硕士学位论文, 四川成都, 2012. pp 3-7
Tang J. Analyses on Heterosis and Combining Ability of the Traits Related to Plant Types in Brassica napus L. MS Thesis of Sichuan Agricultural University, Sichuan, China, 2014. pp 3-7 (in Chinese with English abstract)
[13] 漆丽萍. 甘蓝型油菜株型与角果相关性状的 QTL分析. 华中农业大学博士学位论文, 湖北武汉, 2014. pp 11-12 Qi L P. QTL Analysis for Traits Associated with Plant Architecture and Silique in Brassica napus L. PhD Dissertation of Huazhong Agricultural University, Wuhan, China, 2014. pp 11-12 (in Chinese with English abstract)
[14] Liu J, Wang W, Mei D, Wang H, Fu L, Liu D, Li Y, Hu Q. Characterising variation of branch angle and genome-wide association mapping in rapeseed (Brassica napus L.). Front Plant Sci, 2016,7: 21
[15] 盖钧镒. 植物数量性状遗传体系. 科学出版社, 2003. pp 224-265, 351-370
Gai J Y. Genetic System of Quantitative Traits in Plants. Beijing:Science Press, 2003. pp 224-265, 351-370 (in Chinese)
[16] 李余生, 朱镇, 张亚东, 赵凌, 王才林. 水稻稻曲病抗性的主基因+多基因混合遗传模型分析. 作物学报, 2008, 34:1728-1733
Li Y S, Zhu Z, Zhang Y D, Zhao L, Wang C L. Genetic analysis of rice false smut resistance using major gene plus polygene mixed genetic model. Acta Agron Sin, 2008, 34: 1728-1733 (in Chinese with English abstract)
[17] 周清元, 崔翠, 阴涛, 陈东亮, 张正圣, 李加纳. 甘蓝型油菜角果长度的主基因+多基因混合遗传模型. 作物学报, 2014, 40: 1493-1500
Zhou Q Y, Cui C, Yin T, Chen D L, Zhang Z S, Li J N. Genetic analysis of silique length using mixture model of major gene plus polygene in Brassica napus L. Acta Agron Sin, 2014, 40:1493-1500 (in Chinese with English abstract)
[18] 金岩, 吕艳艳, 付三雄, 戚存扣. 甘蓝型油菜苗期耐淹性状主基因+多基因遗传分析. 作物学报, 2014, 40: 1964-1972
Jin Y, Lü Y Y, Fu S X, Qi C K. Inheritance of major gene plus polygene of water-logging tolerance in Brassica napus L. Acta Agron Sin, 2014, 40: 1964-1972 (in Chinese with English abstract)
[19] 田露申, 牛应泽, 余青青, 郭世星, 柳丽. 甘蓝型油菜白花性状的主基因+多基因遗传分析. 中国农业科学, 2009, 42:3987-3995
Tian L S, Niu Y Z, Yu Q Q, Guo S X, Liu L. Genetic analysis of white flower color with mixed model of major gene plus polygene in Brassica napus L. Sci Agric Sin, 2009, 42: 3987-3995 (in Chinese with English abstract)
[20] 顾慧, 戚存扣. 甘蓝型油菜(Brassica napus L.)抗倒伏性状的主基因+多基因遗传分析. 作物学报, 2008, 34: 376-381
Gu H, Qi C K. Genetic analysis of lodging resistance with mixed model of major gene plus polygene in Brassica napus L. Acta Agron Sin, 2008, 34: 376-381 (in Chinese with English abstract)
[21] 王会, 桑世飞, 梅德圣, 李云昌, 刘佳, 付丽, 王军, 陈玉峰,胡琼. 甘蓝型油菜DH群体抗裂角性遗传分析. 中国油料作物学报, 2014, 36: 437-449
Wang H, Sang S F, Mei D S, Li Y C, Liu J, Fu L, Wang J, Chen Y F, Hu Q. Inheritance analysis of pod shatter resistance using a DH population in Brassica napus L. Chin J Oil Crop Sci, 2014, 36:437-449 (in Chinese with English abstract)
[22] 汪文祥, 胡琼, 梅德圣, 李云昌, 王会, 王军, 付丽, 刘佳. 基于图像处理的油菜分枝及角果着生角度测量方法. 中国油料作物学报, 2015, 37: 566-570
Wang W X, Hu Q, Mei D S, Li Y C, Wang H, Wang J, Fu L, Liu J. Evaluation of branch and pod angle measurement based on digital images from Brassica napus L. Chin J Oil Crop Sci, 2015, 37:566-570 (in Chinese with English abstract)
[23] 曹锡文, 刘兵, 章元明. 植物数量性状分离分析Windows软件包SEA的研制. 南京农业大学学报, 2013, 36(6): 1-6
Cao X W, Liu B, Zhang Y M. SEA: a software package of segregation analysis of quantitative traits in plants. J Nanjing Agric Univ, 2013, 36(6): 1-6 (in Chinese with English abstract)
[24] Diepenbrock W. Yield analysis of winter oilseed rape (Brassica napus L.): a review. Field Crops Res, 2000, 67: 35-49
[25] 马霓, 张春雷, 李俊, 李光明. 种植密度对直播油菜结实期源库关系及产量的调节. 中国油料作物学报, 2009, 31: 180-184
Ma N, Zhang C L, Li J, Li G M. Regulation of planting density on source-sink relationship and yield at seed-set stage of rapeseed (Brassica napus L.). Chin J Oil Crop Sci, 2009, 31: 180-184 (in Chinese with English abstract)
[26] 朱珊, 李银水, 余常兵, 谢立华, 胡小加, 张树杰, 廖星, 廖祥生, 车志. 密度和氮肥用量对油菜产量及氮肥利用率的影响.中国油料作物学报, 2013, 35: 179-184
Zhu S, Li Y S, Yu C B, Xie L H, Hu X J, Zhang S J, Liao X, Liao X S, Che Z. Effects of planting density and nitrogen application rate on rapeseed yield and nitrogen efficiency. Chin J Oil Crop Sci, 2013, 35: 178-184 (in Chinese with English abstract)
[27] 马霓, 李云昌, 胡琼, 李俊, 李光明, 张春雷. 我国南方冬油菜机械化生产农艺研究进展. 中国油料作物学报, 2010, 32:451-456
Ma N, Li Y C, Hu Q, Li J, Li G M, Zhang C L. Research of agronomic techniques form mechanical production of winter rape (Brassica napus L.). Chin J Oil Crop Sci, 2010, 32: 451-456 (in Chinese with English abstract)
[28] 王玫, 陈洪伟, 王红利, 刘克锋. 独脚金内酯调控植物分枝的研究进展. 园艺学报, 2014, 41: 1924-1934
Wang M, Chen H W, Wang H L, Liu K F. Research progress in regulatory role of strigolactones in shoot branching. Acta Hort Sin, 2014, 41: 1924-1934 (in Chinese with English abstract)
[29] Rameau C, Bertheloot J, Leduc N, Andrieu B, Foucher F, Sakr S. Multiple pathways regulate shoot branching. Front Plant Sci,2015, 5: 741
[30] 付三雄, 戚存扣. 甘蓝型油菜含油量的主基因+多基因遗传分析. 江苏农业学报, 2009, 25: 731-736
Fu S X, Qi C K. Major gene plus polygene inheritance of oil content in Brassica napus L. Jiangsu J Agirc Sci, 2009, 25:731-736 (in Chinese with English abstract)
Genetic Effects of Branch Angle Using Mixture Model of Major Gene Plus Polygene in Brassica napus L.
WANG Wen-Xiang, HU Qiong, MEI De-Sheng, LI Yun-Chang, ZHOU Ri-Jin, WANG Hui, CHENG Hong-Tao, FU Li, and LIU Jia*
Oil Crops Research Institute of Chinese Academy of Agricultural Sciences / Key Laboratory of Biology and Genetic Improvement of Oil Crops of the Ministry of Agriculture, Wuhan 430062, China
Branch angle is one of important plant architectural traits in rapeseed (Brassica napus), which is the basic requirement for high-density cultivation and mechanical harvesting. To reveal the genetic mechanism of branch angle in rapeseed, we produced six generations (P1, P2, F1, F2, BCP1, and BCP2) from a cross between loose-type 6098B and compact-type Purler to analyze genetic effects of branch angle in rapeseed by the mixed major gene plus polygene inheritance model. The branch angle showed a continuous normal distribution, where both the basal branch angle and the top branch angle were dominated by a pair of major gene with additive-dominant effects plus polygenes with additive-dominance-epistasis effects (D-0 model). The additive effect and dominant effect of major gene of the top branch angle were 4.939 and -4.156, the heritability in BCP1, BCP2, and F2was 34.08%, 1.40%, and 14.99%, respectively for the major gene, and 24.43%, 61.72%, and 63.98%, respectively for the polygenes. The additive effect and dominant effect of major gene of the basal branch angle were 2.217 and -1.941, the heritability in BCP1,BCP2, and F2was 7.86%, 1.24%, and 4.84%, respectively for the major gene, and 66.46%, 58.49%, and 73.96%, respectively for the polygenes. Results of this study found that the branch angle traits of 6098B×Purler is controlled by polygenes, but there is a major gene among them. The research may be useful for further understanding and genetic improvement of plant architecture inrapeseed.
Brassica napus L.; Branch angle; Ideal plant architecture; Major gene plus polygene; Genetic effects
10.3724/SP.J.1006.2016.01103
本研究由中国农业科学院科技创新工程项目(Group No. 118), 国家农业现代产业技术体系建设专项(CARS-13), 国家自然科学基金项目(31471535), 湖北省自然科学基金项目(2014CFB156)和湖北省科技创新工程项目资助。
This study was support by the Science and Technology Innovation Project of Chinese Academy of Agricultural Sciences (Group No. 118), the Earmarked Fund for China Agriculture Research System (CARS-13), the Natural Science Foundation of China (31471535), the Natural Science Foundation of Hubei Province (2014CFB156) and the Hubei Agricultural Science and Technology Innovation Center.
*
(Corresponding author): 刘佳, E-mail: liujia02@caas.cn, Tel: 027-86711556
联系方式: E-mail: wangwx8@gmail.com
Received(
): 2016-01-14; Accepted(接受日期): 2016-05-09; Published online(网络出版日期): 2016-05-30.
URL: http://www.cnki.net/kcms/detail/11.1809.S.20160530.0905.014.html
