外寄生性花绒寄甲的寄生选择及其发育表现
2016-08-09李广花李保平徐福元郑华英解春霞
李广花,李保平,徐福元,郑华英,解春霞,孟 玲,*
1 南京农业大学植物保护学院,农作物生物灾害综合治理教育部重点实验室,南京 210095 2 江苏省林业科学院森林保护研究所,南京 211153
外寄生性花绒寄甲的寄生选择及其发育表现
李广花1,李保平1,徐福元2,郑华英2,解春霞2,孟玲1,*
1 南京农业大学植物保护学院,农作物生物灾害综合治理教育部重点实验室,南京210095 2 江苏省林业科学院森林保护研究所,南京211153
摘要:“选择-表现”假说认为,成虫应该选择有利于子代发育的高品质寄主,但在寄主选择中,除了寄主品质外,其他因素也可能影响寄主选择决策。寄主选择研究通常以成虫为对象,而对那些初龄幼虫选择寄主的寄生性昆虫很少关注。以1龄幼虫积极搜寻寄主的寄生性花绒寄甲为模式生物,采用双选试验设计,观察了花绒寄甲初孵幼虫在不同体重青杨天牛幼虫之间、在已被寄生与健康的黄粉虫蛹之间的寄生选择性;然后采用回归设计,观察了花绒寄甲寄生若干不同体重的青杨天牛幼虫后的发育表现。研究结果表明,花绒寄甲1龄幼虫对体型较大的青杨天牛幼虫的选择偏好显著大于对体型较小的寄主幼虫的选择,选择大体型幼虫的比值比是选择小体型幼虫的4.55倍;对已被寄生的寄主黄粉虫蛹的选择偏好显著大于对健康寄主蛹的选择,选择已被寄生寄主的比值比是选择健康寄主的 12.57倍。寄生青杨天牛幼虫的花绒寄甲幼虫发育历期平均为11.49 d、蛹历期为26.67 d、幼虫发育至成虫的羽化率>50%, 这些发育表现与寄生时青杨天牛幼虫的体重没有显著关系。但刚羽化寄甲成虫体重与寄生时寄主的体重存在显著的正直线关系:寄生时的寄主体重每增大0.01 g,羽化出的寄甲成虫体重增大近0.08%;方差分析寄甲成虫体重在不同寄主体重水平之间的差异表明,从体型较大寄主中羽化的寄甲成虫体重显著大于从体型较小寄主中羽化的成虫。研究结果说明,花绒寄甲初孵幼虫在寄主选择决策时,在寄主体型大小与被寄生状态之间可能采取折衷对策,而且对体型大小不同的寄主选择与子代发育适合度表现存在一致性,从而支持“选择-表现”假说。
关键词:生活史进化;初龄幼虫;发育表现;折衷;寄主体型大小
昆虫产卵选择与其子代发育、生存和生殖表现的关系,一直是植食性昆虫与植物进化生态学研究的核心问题[1]。“选择-表现”假说认为,当子代不善活动的植食性昆虫选择植物产卵时,会选择使子代发育和存活最优的场所产卵。该假说不仅在许多植食性昆虫中得到了支持[2- 3],而且在诸如食蚜蝇[4]、食蚜瘿蚊[5]以及寄生蜂[6]等肉食性昆虫中也获得了支持,但也有一些研究不支持该假说[1- 2]。说明昆虫在产卵选择时不仅考虑寄主品质,而且参考其他因素。通常,这类研究均以成虫期负责产卵选择的昆虫作为对象,而在有些昆虫中,负责选择的不是成虫,而是初龄幼虫,例如,鞘翅目芫菁科(Meloidae)[7]和穴甲科(Bothrideridae)[8]中有些营寄生生活的种类,依赖于胸足发达的初龄幼虫搜寻和选择寄主,找到寄主后脱皮变成足不发达的幼虫,依赖于该寄主完成幼期发育。迄今,鲜见以幼虫负责搜寻和选择的寄生性昆虫为模式生物来检验“选择-表现”假说。
花绒寄甲(Dastarcushelophoroides)是星天牛类(包括黄斑星天牛Anoplophoranobilis、光肩星天牛A.glabripennis和星天牛A.chinensis) 以及其他天牛(包括松墨天牛Monochamusalternatus、云斑天牛Batocerahorsfieldi、锈色粒肩天牛Aprionaswainsoni等)的寄生性天敌[8- 10],对防治松墨天牛等林木蛀干天牛害虫具有良好的防效[11-15]。花绒寄甲成虫将卵产在树枝、干缝隙中,孵化的1龄幼虫具有很强的搜索能力,依靠发达的胸足积极搜寻虫道中的天牛幼虫;找到寄主后取食、脱皮变成“拟蛆型”幼虫,依赖于找到的寄主完成其后的幼虫发育期[16-18]。所以,负责搜寻的1龄幼虫所选择的寄主与适合度相关发育表现密切相关。根据进化生物学基本原理预测,花绒寄甲1龄幼虫面临强大的自然选择压力,应对寄主具有很强的搜寻和选择性。
本研究以花绒寄甲为模式生物,用青杨天牛(Saperdapopulnea)幼虫和黄粉虫(Tenebriomolitor)蛹为寄主,对以下具体问题进行研究:(1)花绒寄甲1龄幼虫寄生选择时在不同体型大小寄主之间、在已被寄生与未被寄生的寄主之间是否存在选择性?(2)不同体型大小的寄主如何影响花绒寄甲的适合度相关发育表现?对第1个问题的回答,将有助于推测寄甲在选择寄主时是否在寄主品质上采用折中(trade-off)策略;对第2个问题的回答,将有助于明确寄甲在不同体型大小寄主中的发育是否与其寄生选择存在正相关性,以检验“选择-表现”假说是否适用于依靠初孵幼虫选择的昆虫。
1材料与方法
1.1供试昆虫
花绒寄甲为江苏省林业科学研究院森林保护研究所大规模繁殖种群。供试前将卵卡剪成约0.5 cm × 1 cm小块 (约100粒卵)置于指形管(直径0.8 cm、高4.8 cm)中,保存于7 ℃恒温箱内待用。青杨天牛广泛分布于我国北方和西北地区,是当地主栽树种——杨树的重大蛀干害虫[19- 20]。在利用花绒寄甲防治天牛害虫的实践中,业已利用青杨天牛为寄主扩繁花绒寄甲。青杨天牛老熟幼虫采自山西省运城市绛县以毛白杨(Populustomentosa)为主的杨树被害枝条,冬季采集被害枝条,运至繁育室冷藏,于早春从枝条中剖出幼虫供试。黄粉虫购自南京市秦淮区集贸市场,在养虫室用麦麸加土豆片饲养,取24 h内化的蛹供试。
1.2试验方法
1.2.1花绒寄甲初孵幼虫的寄生选择
采用2个双选试验观察花绒寄甲幼虫的寄生选择。第1个试验旨在观察寄甲在体重大、小不同的天牛寄主幼虫之间的选择,大体型寄主的体重为(0.0780±0.0078)g(平均值±标准差),小体型寄主体重为(0.0412±0.0041)g。首先将体型大小不同的青杨天牛幼虫单头称重(METTLER TOLEDO电子天平AL204-IC,精确至0.0001g),从中选取体重相差约2倍的青杨天牛幼虫各1头,并排放入培养皿(直径5 cm、高1.5 cm)中间,然后用软毛笔接入1头当天孵化的花绒寄甲幼虫,于2日后在解剖镜下检查寄甲所寄生的寄主。试验重复50次。
第2个试验旨在观察寄甲幼虫在已被寄生与健康寄主黄粉虫蛹之间的选择。单头称重寄主(METTLER TOLEDO电子天平AL204-IC,实际分度值为0.0001 g),选择体重接近的寄主幼虫供试,健康寄主幼虫体重为(0.1416±0.0210)g,被寄生寄主体重为(0.1418±0.0214)g。首先,把1头寄主放入培养皿(直径5 cm、高1.5 cm)中,释放1头当日孵化的花绒寄甲幼虫,2d后在解剖镜(Nikon SMZ800)下检察是否寄生(以寄甲幼虫开始取食寄主作为寄生表现);当发现寄甲开始寄生后,再放入1头健康的黄粉虫蛹,靠近(但不接触)已被寄生的寄主,然后释放1头当天孵化的花绒寄甲初孵幼虫,两天后观察该寄甲初孵幼虫的寄生选择。试验重复50次。
1.2.2花绒寄甲寄生不同体型大小青杨天牛幼虫的发育表现
为揭示寄甲发育特征与寄主体型大小的关系,采用回归设计,该设计的特点是设置尽可能多的因子处理水平但不强调重复,特别适于揭示自变量与因变量之间的数量关系,从而为建立生态模型提供依据[21-22]。取青杨天牛老熟幼虫称重(METTLER TOLEDO电子天平AL204-IC,精确到0.0001 g),单头放入24孔板的单一孔(直径1.5 cm、高2 cm)中,然后用软毛笔向每孔释放1头当日孵化的花绒寄甲初孵幼虫。每天观察3次,记录寄生与否,如遇寄主死亡,及时补充体重相当的寄主;观察记录花绒寄甲化蛹和羽化情况,在羽化后12 h内取花绒寄甲成虫称鲜重。以上试验均在室内进行,环境温度24—27℃,相对湿度约60%—70%。共观察166头寄主。
1.3数据分析
由于选择性实验的观测变量为二进制数据(选择=1,不选择=0),符合二项分布型,故用logistic回归模型分析花绒寄甲选择某一类型寄主的概率;在分析对黄粉虫蛹的选择试验数据时,将寄主体重作为协变量以排除寄主体重的影响。寄甲幼虫是否存活至成功羽化的观测值为二进制数据,符合二项分布型,符合Poisson分布故用logistic模型拟合羽化概率随寄主体重增大的变化趋势。寄甲幼虫和蛹发育天数为整数数据,故用Poisson模型分析幼虫和蛹历期随寄主体重增大的变化趋势。用似然比测验比较模型进行模型简化。寄甲成虫体重为连续数值数据,符合正态分布,用一般线性回归模型分析寄甲成虫体重随寄主幼虫体重增大的变化趋势;然后将寄主体重平均划分为低、中和高3个水平,用方差分析检验不同水平之间是否存在显著差异,若有显著差异,再用TukeyHSD测验比较不同水平之间的差异。用R软件分析数据[23]。
2结果与分析
2.1花绒寄甲初孵幼虫的寄生选择
花绒寄甲初孵幼虫对不同体重的寄主幼虫表现出显著的偏好性(对数似然比测验:χ2=44.35,P<0.001),对较重寄主的寄生偏好显著大于对较轻寄主的寄生(比值比=4.55,95%置信域=2.32—10.00,WaldZ=5.83,P<0.001)(图1)。花绒寄甲初孵幼虫在被其他花绒寄甲寄生与健康寄主黄粉虫蛹之间表现出显著偏好(对数似然比测验χ2=33.25,P<0.001),对被选择寄生寄主的选择概率远大于对健康寄主的选择(比值比=12.57; 95%置信域=5.05—33.94,WaldZ=5.24,P< 0.001)(图1)。
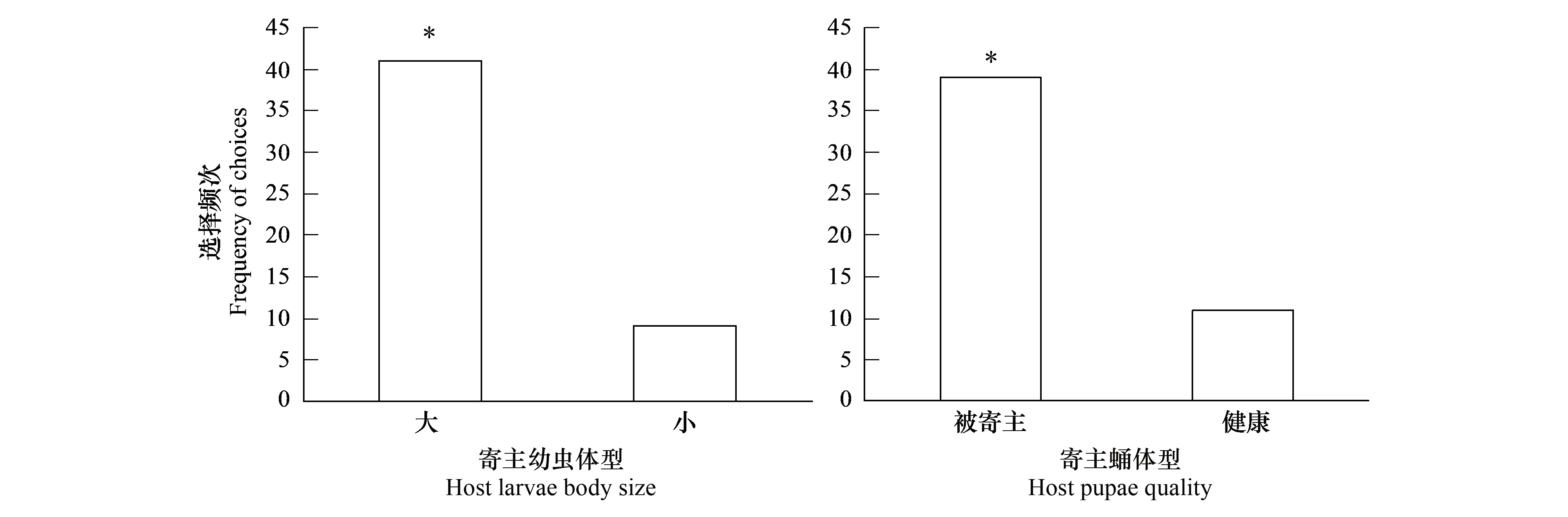
图1 花绒寄甲初孵幼虫对不同体型大小青杨天牛寄主幼虫和寄生与否的黄粉虫寄主蛹的选择频次Fig.1 Frequency of choices between S. populnea larvae differed in body size and between T. molitor pupae differed in parasitism by D. helophoroides 1st instar larvae *表示两选项之间存在显著差异(P<0.05)
2.2青杨天牛幼虫寄主体型大小对花绒寄甲发育和存活的影响
寄主青杨天牛幼虫的体重对花绒寄甲幼虫发育历期没有显著影响(似然比测验χ2=0.09,P=0.77),幼虫历期平均为11.49 d (95%置信域=11.11—11.87 d,n=122) (图2)。寄主幼虫体重对蛹历期没有显著影响(似然比测验χ2=1.33,P=0.25),蛹历期平均为26.67 d (95%置信域=26.32—27.02 d,n=110) (图3)。寄主幼虫体重对寄甲幼虫发育至成虫羽化的存活率没有显著影响(似然比测验χ2=0.016,P=0.90)(图4)。寄主幼虫体重显著影响羽化成虫的体重(F1,108=58.98,P<0.001),刚羽化成虫的体重随寄主体重增大而增大,寄主体重每增大0.01 g寄甲成虫体重增大近0.08%(图5);当根据体重把寄主体型大小划分为小(0.084—0.131 g)、中等(0.131—0.178 g)和大(0.178—0.225 g)等3个水平时,方差分析表明寄甲成虫体重在寄主大小

图2 花绒寄甲幼虫历期与寄生时青杨天牛寄主幼虫体型大小的关系Fig.2 Larval development time of D. helophoroides in relation to S. populnea larval body weight at parasitism

图3 花绒寄甲蛹期与寄生时青杨天牛寄主幼虫体型大小的关系Fig.3 Pupal duration of D. helophoroides in relation to S. populnea larval body weight at parasitism

图4 幼虫至成虫羽化概率与寄生时青杨天牛寄主幼虫体型大小的关系Fig.4 Larva-to-adult emergence success of D. helophoroides in relation to S. populnea larva body weight at parasitism绘图时采用颤抖技术以避免数据点重叠

图5 羽化成虫体重与寄生时青杨天牛寄主幼虫体型大小的关系Fig.5 Adult body weight at emergence of D. helophoroides in relation to S. populnea larval body weight at parasitism拟合线模型为:寄甲成虫体重=0.017 (± 标准误=0.0017)+ 寄主体重 × 0.079 (± 0.010); 一般线性模型拟合度:R2=0.35,F1,108=58.98,P<0.001
不同水平间存在显著差异(F2,107= 28.85,P< 0.0001),TukeyHSD多重比较测验表明,各寄主水平间均存在显著差异(P<0.05),从大体型寄主中羽化的寄甲成虫比从小体型寄主中羽化的成虫重0.008 g,比从中等体型寄主中羽化的成虫重0.002 g。
3讨论
生活史的进化受制于折衷:当生物个体因某一生活史特征的改变而获益时,会因另一特征的相应变化而付出代价[24]。探究哪些生活史特征之间在影响生物适合度上存在折衷,是进化生态学研究的核心问题之一[25]。本研究的选择性试验结果表明,花绒寄甲初孵幼虫不仅偏好寄生体型较大的青杨天牛幼虫,而且偏好已被其他幼虫寄生的寄主。根据该寄生选择行为可推测,花绒寄甲初孵幼虫在寄主选择中可能采取折衷的选择对策,即在寄主体型大小与其是否已被寄生之间折衷。一方面, 与小体型寄主相比,大体型寄主含有更多的营养资源,但体型较大的寄主通常具有更强的防卫能力,需要付出更大的代价才能获得,而合作制服寄主可降低各自的代价。王卫东和小仓信夫[17]观察发现,虽然1头寄甲1龄幼虫可麻醉1头体型较大的松褐天牛幼虫,但通常需较长的时间(3—12 d),而且1次寄生成功率只有75%;而5头寄甲幼虫联合只需1 d即可麻醉老熟幼虫。另一方面,虽然合作可有效克服寄主防卫,但需分享、竞争寄主资源,从而付出适合度降低(低于单独拥有该寄主的适合度)的代价。有研究表明,花绒寄甲幼虫可多头共享松褐天牛幼虫,5头以下共同寄生对发育没有明显影响,但更多寄甲共享1头天牛幼虫时,老熟幼虫体型变小、结白色茧或不能结茧、甚至不能完成发育[17]。
然而,似乎与上述折衷不一致的证据来自于对体型较大的松褐天牛寄主的研究。该研究采用多选试验设计观察寄甲初龄幼虫对小、中和大型(体长)松褐天牛幼虫的寄生选择后发现,寄甲偏好选择中等大小(体长)的寄主[18]。但由于该研究未提供备选不同体长寄主的比例信息,也未做统计分析,故有待进一步严格的实验验证。
本研究虽未发现幼虫期发育历期、蛹历期以及羽化率等生活史特征受寄主体型大小的显著影响,但刚羽化成虫的体重随寄主体重增大而显著增大,从体型较大寄主中羽化的寄甲成虫体重显著大于从体型较小寄主中羽化的成虫 (寄主体型大与小的平均体重相差1倍)。发育时间虽然对于内寄生性昆虫至关重要,尤其是寄生营暴露取食寄主的寄生蜂,因为寄主的存活直接关系到寄生蜂幼虫的生存[26]。但对于外寄生性昆虫而言,由于寄生物发育不受制于寄主血腔的限制,故发育时间的可塑性可以更大[26]。一般认为,成虫体型大小(通常用体重代表)是其繁殖力和适合度的线性函数,从而成为衡量昆虫繁殖力[27]和适合度[28]的一项重要的适合度相关特征。
4结论
本研究结果说明,花绒寄甲虽然可以在小体型寄主上完成幼期发育,但成功羽化及其适合度在体型较大的寄主上得到明显的提高。该发育表现与1龄幼虫偏好较大体型寄主的寄生选择性存在正相关性,从而支持“选择-表现”假说。由此说明,该假说可用于解释依赖幼虫选择资源的其他昆虫,从而为支持该假说提供了新的证据。
参考文献(References):
[1]Thompson J N. Evolutionary ecology of the relationship between oviposition preference and performance of offspring in phytophagous insects. Entomologia Experimentalis et Applicata, 1988, 47(1): 3- 14.
[2]Thompson J N, Olle Pellmyr. Evolution of oviposition behavior and host preferences in Lepidoptera. Annual Review of Entomology, 1991, 36(1): 65- 89.
[3]Bonebrake T C, Boggs C L, McNally J M, Ranganathan J, Ehrlich O R. Oviposition behavior and offspring performance in herbivorous insects: consequences of climatic and habitat heterogeneity. Oikos, 2010, 119(6): 927- 934.
[4]Sadeghi H, Gilbert F. Individual variation in oviposition preference, and its interaction with larval performance in an insect predator. Oecologia, 1991, 118(4): 405- 411.
[5]Jandricic S E, Wraight S P, Gillespie D R, Sanderson J P. Oviposition behavior of the biological control agentAphidoletesaphidimyza(Diptera: Cecidomyiidae) in environments with multiple pest aphid species (Hemiptera: Aphididae). Biological Control, 2013, 65(2): 235- 245.
[6]Godfray H C. Parasitoids: Behavioral and Evolutionary Ecology. Princeton, New Jersey: Princeton University Press, 1994.
[7]Imms A D, Richards O W, Davies R G. Imms′ General Textbook of Entomology. Vol. 2: Classification and Biology. New York: John wiley & Sons, Inc., 1977: 889- 891.
[8]周嘉熹, 鲁新政, 逯玉中. 引进花绒坚甲防治黄斑星天牛试验报告. 昆虫知识, 1985, 22(2): 84- 86.
[9]秦锡祥, 高瑞桐. 花绒坚甲生物学特性及应用研究. 昆虫知识, 1988, 25(2): 109- 112.
[10]王希蒙, 任国栋, 马峰. 花绒穴甲的分类地位及应用前景. 西北农业学报, 1996, 5(2): 75- 78.
[11]杨忠岐. 利用天敌昆虫控制我国重大林木害虫研究进展. 中国生物防治, 2004, 20(4): 221- 227.
[12]Urano Tadahisa. Experimental release of adultDastarcushelophoroides(Coleoptera: Bothrideridae) in a pine stand damaged by pine wilt disease: Effects onMonochamusalternatus(Coleoptera: Cerambycidae). Bulletin of FFPRI, 2006, 5(4): 257- 263.
[13]崔永三, 刘云鹏, 宋玉双, 徐福元, 徐克勤. 应用天敌昆虫防治松墨天牛试点试验. 中国森林病虫, 2011, 30(4): 31- 33.
[14]刘跃进, 喻锦秀, 周刚, 戴立霞, 杨忠岐, 张翌楠. 释放天敌昆虫对松褐天牛的防治效果研究. 湖南林业科技, 2012, 39(5): 20- 23.
[15]杨远亮, 杨忠岐, 王小艺, 喻锦秀, 颜学武. 应用花绒寄甲防治松褐天牛. 林业科学, 2013, 49(3): 103- 109.
[16]雷琼, 李孟楼, 杨忠歧. 花绒坚甲的生物学特性研究. 西北农林科技大学学报: 自然科学版, 2003, 31(2): 62- 66.
[17]王卫东, 小仓信夫. 花绒穴甲室内发育研究. 北京林业大学学报, 1999, 21(4): 43- 47.
[18]王小东, 黄焕华, 许再福, 杨忠岐, 范军祥, 特拉康, 陈驹坚, 钱明惠. 花绒坚甲的生物学和生态学特性研究初报. 昆虫天敌, 2004, 26(2): 60- 65.
[19]孙萍, 郭树平, 李海霞. 杨树单宁含量与青杨天牛危害的关系. 东北林业大学学报, 2008, 36(5): 51- 52.
[20]段治卿. 不同混交林青杨天牛种群动态研究. 山西农业科学, 2010, 38(1): 88- 90.
[21]Gotelli N J, Ellison A M. A Primer of Ecological Statistics. 2nd ed. Sunderland, MA: Sinauer Associates Inc, 2013.
[22]Cottomgham K L, Lennon J T, Brown B L. Knowing when to draw the line: designing more informative ecological experiments. Frontiers in Ecology and Environment, 2005, 3(3): 145- 152.
[23]R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria, 2014. http://www.R-project.org/.
[24]Stearns S C. The Evolution of Life History. London: Oxford Univresity Press, 1992: 14- 14.
[25]Begon M, Townsend C, Harper J L. Ecology: From Individuals to Ecosystems. 4th ed. London: Wiley-Blackwell, 2006: 110- 116.
[26]Strand M R. Development traits and life-history evolution in parasitoids. In: Hochberg M E, Ives A R eds. Parasitoid Population Biology. Princeton, New Jersey: Princeton University Press, 2000: 139- 162.
[27]Honek A. Intraspecific variation in body size and fecundity in insects: a general relationship. Oikos, 1993, 66(3): 483- 492.
[28]Roitberg B D, Boivin G, Vet L E M. Fitness, parasitoids, and biological control: an opinion. The Canadian Entomologist, 2001, 133(3): 429- 438.
基金项目:江苏省林业三新工程(lysx[2014]17); 国家公益性行业(农业)专项(201103002)
收稿日期:2014- 10- 20; 网络出版日期:2015- 10- 10
*通讯作者
Corresponding author.E-mail: ml@njau.edu.cn
DOI:10.5846/stxb201410202057
Parasitization preference and developmental performances in the ectoparasitoidDastarcushelophoroides(Coleoptera:Bothrideridae)
LI Guanghua1, LI Baoping1, XU Fuyuan2, ZHENG Huaying2, XIE Chunxia2, MENG Ling1,*
1CollegeofPlantProtection,KeyLaboratoryofMonitoringandManagementforPlantDiseasesandInsectPests,MinistryofAgriculture,NanjingAgriculturalUniversity,Nanjing210095,China2InstituteofForestProtection,JiangsuAcademyofForestry,Nanjing211153,China
Abstract:The preference-performance hypothesis predicts that adults should select high quality hosts which can provide offspring with good resources for development. Other factors, however, can also influence the parasitoids in making decisions on the choice. Previus such studies often use adult parasitoids as model systems, but give scant attention to insects in which host search and selection are made by neonate larvae. Here we used as the model system the ectoparasitic beetle Dastarcus helophoroides, which depends its neonate larvae to search and select hosts. We first, using the long-horned beetle Saperda populnea larvae and the flour beetle Tenebrio molitor pupae as hosts, investigated whether D. helophoroides first instar larvae showed parasitization preferences between hosts varying in body size or in parasitism by conspecifics. We then, using the experimental regression design, examined developmental performances in relation to host body weight at parasitism. D. helophoroides first instar larva showed a significant preference for larger sized S. populnea larva, which were 4.55 times as likely to be parasitized as the smaller sized larva. D. helophoroides first instar larva preferred the parasitized T. molitor pupa over the healthy for parasitization, 12.57 times as likely for the parasitized as for the healthy host pupae. D. helophoroides larval development time (averaged to 11.49 d), pupal duration (26.67 d), and larva-to-adult survival rate (> 50%) were not related with host body weight at parasitism. But adult body weight at emergence significantly increased linearly with host body weight at parasitism, by an increase of almost 0.08% with each 0.01 g increment in host body weight. ANOVA and multiple comparison tests showed that D. helophoroides adult at emergence was significantly larger in body weight from attacking larger than smaller sized hosts. The results of our study resuggest that D. helophoroides first instar larva may take a trade-off between host body size and parasitism in making decisions on parasitization. In addition, our results suggest that the prefernece for larger hosts in body size by D. helophoroides first instar larva is aligned with developmental performances of offpring, lending suport for the preference-performance hypothesis.
Key Words:life history evolution; first instar larvae; developmental performances; trade-off; host body size
李广花,李保平,徐福元,郑华英,解春霞,孟玲.外寄生性花绒寄甲的寄生选择及其发育表现.生态学报,2016,36(12):3796- 3801.
Li G H, Li B P, Xu F Y, Zheng H Y, Xie C X, Meng L.Parasitization preference and developmental performances in the ectoparasitoidDastarcushelophoroides(Coleoptera:Bothrideridae).Acta Ecologica Sinica,2016,36(12):3796- 3801.
