羊草功能性状和地上生物量对氮素添加的响应
2016-08-02宋彦涛王平周道玮乌云娜
宋彦涛,李 强,王平,周道玮,乌云娜
(1.大连民族大学环境与资源学院,辽宁 大连 116600; 2.中国科学院东北地理与农业生态研究所,吉林 长春 130102;3.东北师范大学环境学院,吉林 长春 130117)
羊草功能性状和地上生物量对氮素添加的响应
宋彦涛1,李 强2,王平3,周道玮2,乌云娜1
(1.大连民族大学环境与资源学院,辽宁 大连 116600; 2.中国科学院东北地理与农业生态研究所,吉林 长春 130102;3.东北师范大学环境学院,吉林 长春 130117)
摘要:植物功能性状对资源变化的适应策略是生态学研究的基本问题之一。本研究在不同氮添加梯度处理下,测定了羊草(Leymus chinensis)的11个功能性状和地上生物量的变化。结果表明,氮添加显著影响羊草的叶片叶绿素a和b含量、叶片类胡萝卜素含量、叶片氮含量、叶片干物质含量和比根长(P<0.05),而对叶片磷含量、比叶面积、叶片厚度、比茎重和植株高度没有显著作用(P>0.05);叶绿素a和b含量、类胡萝卜素含量、叶片氮含量、叶片干物质含量、比茎重和比根长之间有显著的相关关系(P<0.05),比叶面积、叶片厚度和植株高度与其它功能性状的相关性不显著(P>0.05);20 g·m-2的氮添加处理下,羊草地上生物量最高;羊草地上生物量随叶绿素a和b含量、类胡萝卜素含量、叶片氮磷含量、株高的增大而显著升高(P<0.05),随叶片干物质含量、比茎重的增大而显著降低(P<0.05)。
关键词:氮添加;羊草;叶片性状;松嫩草地
氮素是植物生长的必需元素[1],适量的氮添加能促进植物对养分的吸收,从而提高植物生产力[2-3]。而对于氮饱和的地区,氮添加则抑制植物生长[4]。氮添加改变植物生长的环境,而植物能通过功能性状变化调整对环境变化(包括氮沉降)的适应策略。氮添加能够影响植物的结构和生理性状[5]、生长与构件分配[4],改变植物在群落中的竞争力,影响群落结构和组成,并使生态系统功能发生改变[6]。在内蒙古羊草(Leymus chinensis)草原的研究表明,氮添加能够增加羊草比叶面积、叶绿素含量和叶片氮含量,从而增加羊草对光竞争的能力[5],而且向根系和叶片的分配比例增加,对根茎的分配投入减少[2],也能改变稳定草地群落和退化草地群落的物种丰富度和功能群组成[6]。氮添加能够增加高寒草甸植物群落高度、比叶面积,影响物候期和物种多样性,最终改变群落组成[7]。施氮后,高寒矮嵩草(Kobresia humilis)草甸植物群落的丰富度和功能多样性显著降低,并且这些变化对群落初级生产力有直接的影响[8]。
松嫩草地以营养价值高、适口性好的羊草蜚声海内外,而且羊草在松嫩草地往往形成单优势植被[9]。近30多年,学者们在松嫩草地研究了氮添加对草地产量和质量的作用及所影响的经济效益[10]。氮添加能够增加羊草株高、密度、产量和蛋白质含量,并能提高土壤肥力[11],氮添加1个月后其对羊草草地促进效果最佳,而后降低[12]。氮添加能显著影响羊草的叶面积和籽实重量[13],延后羊草的抽穗期,提高羊草在群落中的重要值,降低羊草群落的多样性和均匀性[14]。这些研究更多关注羊草在松嫩草地的生产性能,对羊草功能性状研究的较少。氮添加能增加土壤资源的有效性,改变植物生长环境条件,因此,研究不同梯度氮添加对松嫩草地羊草功能性状及羊草生产力的影响,有助于从性状角度解释羊草草地结构、功能表现及其变化的原因,并为草地生产管理提供参考。
1材料和方法
1.1研究区域概况
研究地点位于中国科学院长岭草地农牧生态研究站(44°33′N,123°31′E,海拔145m)。该区为温带大陆性季风气候,年均温4.9 ℃,≥10 ℃年有效积温2 920 ℃·d,无霜期140~160d,年日照时数2 800h;年降水量300~500mm,70%集中在6-9月;年蒸发量1 600mm,约是降水量的3.5倍。该区优势植被为羊草,土壤类型主要为黑钙土,pH为7.5~9。
1.2试验设计
试验在天然草地上进行,采用完全随机设计,小区面积1.5m×1.5m,5次重复。2008年7月中旬的雨季施尿素(吉林松原长山化肥集团长达公司,含氮量46.4%),施氮梯度0、10、20、30、40g·m-2,0为对照。准确称量每个小区的尿素量,均匀撒于地表,8月下旬地上生物量最大时取样。
1.3取样和测定方法
选取常见的功能性状指标进行测定,包括叶片叶绿素a含量(leafchlorophyllacontent,Cha,mg·g-1),叶片叶绿素b含量(leafchlorophyllbcontent,Chb,mg·g-1),叶片类胡萝卜素含量(leafcarotenoidcontent,Cad,mg·g-1),叶片氮含量(leafnitrogencontent,LNC,mg·g-1),叶片磷含量(leafphosphoruscontent,LPC,mg·g-1),叶片干物质含量(leafdrymattercontent,LDMC,g·g-1),比叶面积(specificleafarea,SLA,cm2·mg-1),叶片厚度(leafthickness,LT,mm),比茎重(stemspecificdensity,SSD,mg·mm-3),比根长(specificrootlength,SRL,m·g-1),植株高度(plantheight,PH,cm)和羊草地上生物量(abovegroundbiomass,AB,g·m-2)。
每个小区采集新长出完全展开的成熟羊草叶片约30g迅速带回实验室105 ℃杀青18min,然后75 ℃烘干至恒重,粉碎后放入封口袋保存待测。按照上述相同标准在每个小区采集5片羊草叶片,装入有少量去离子水的封口袋中,然后放入保温箱中并尽快带回实验室,在无光条件下于4 ℃恒温冰箱中保存12h。取出后用定性滤纸擦干叶片表面水分,称重(万分之一天平)即为叶片的饱和水鲜重;LT用厚度仪(YH-1型叶片厚度测定仪)测量;照相后用SigmaScanPro4 (SystatEoftwareInc.,California,USA)计算每个羊草叶片的叶面积;最后把叶片放入75 ℃烘箱烘干至恒重。SLA为叶片面积除以叶片干重,LDMC为叶片干重除以叶片饱和水鲜重。每个小区挖10段10cm的羊草根茎,带回实验室洗净后75 ℃恒温下烘干至恒重,SRL=根茎长度/根茎干重。每个小区随机选10株羊草测量株高。每个小区选中心齐地面收割1m×1m的羊草,75 ℃恒温下烘干至恒重后称量,计算羊草地上生物量。
LNC和土壤全氮含量用凯式定氮法测定(Kjeltec2003AnalyzerUnit,FOSSTECATOR,Höganäs,Sweden),LPC和土壤全磷含量用NaOH熔融-钼锑钪比色法测定[15]。Cha、Chb、Cad用分光光度法测定[16]。
取植物样品的同时,对试验设计的中未施氮的5个小区进行1m×1m的群落学调查,分物种测定植株高度、密度、地上生物量,地上生物量采用收获法测定,75 ℃恒温下烘干至恒重;取0-10cm层土壤,带回实验室风干、过筛,测定土壤pH、电导率、有机碳、全氮、全磷等指标作为样地本地信息。采用三因素综合优势比确定不同放牧梯度上物种成分的优势度(SDR3):
SDR3=(密度比+盖度比+干重比)/3×100%。
式中:密度比为某一物种的密度占群落密度最高的物种密度的百分比;高度比为某一物种的高度占高度最大物种的高度的百分比;干重比为某一物种的地上生物量干重占地上生物量干重最大物种干重的百分比。
1.4数据处理
先检验正态分布和方差齐性,不满足条件首先进行数据转化,然后进行单因素方差分析和多重比较,Turkey检验均值间的差异性,显著性水平a=0.05。性状间的相关性用Pearson相关分析统计(双尾检验,a=0.05),一般线性模型用于建立羊草地上生物量和性状间的回归关系。数据处理使用SPSS软件 (2004,ver. 13.0;SPSSInc.,USA)完成。
2结果与分析
从研究地点的本底信息可以看出,群落结构简单,物种丰富度低,以单优势的羊草为主,其地上生物量占群落地上生物量的97.3%,密度占群落密度的95.2%,羊草具有绝对优势(表1)。土壤的pH、电导率、有机碳含量、全氮量、全磷含量分别为8.96、208μs·cm-1、19.83mg·g-1、2.32mg·g-1和0.35mg·g-1。
方差分析结果表明,氮添加处理对羊草叶片磷含量、比叶面积、叶片厚度、比茎重和植株高度无显著影响(P>0.05),对其它功能性状有显著影响(P<0.05)(图1)。对照和10g·m-2处理的羊草叶绿素a含量差异不显著(P>0.05),20、30、40g·m-2氮添加处理间叶绿素含量差异不显著,但均显著高于对照(P<0.05)。氮添加显著增加了叶绿素b的含量(P<0.05),但各添加处理之间差异不显著(P>0.05)。氮添加处理对类胡萝卜素含量的作用与叶绿素a结果一致。氮添加显著增加了叶片氮含量,但在10 和20g·m-2处理间差异不显著(P>0.05),30和40g·m-2处理与20g·m-2处理间氮含量差异不显著,但显著高于10g·m-2处理和对照(P<0.05)。氮磷比随着氮添加量的增加而增加,对照区氮磷比为20,氮添加处理氮磷比均高于24。对照叶片干物质含量显著高于氮添加处理(P<0.05),氮添加各处理间差异不显著(P>0.05)。比根长也随氮添加量的增加而增加。10和20g·m-2处理下的比根长和对照差异不显著(P>0.05),30和40g·m-2处理下比根长显著高于对照。
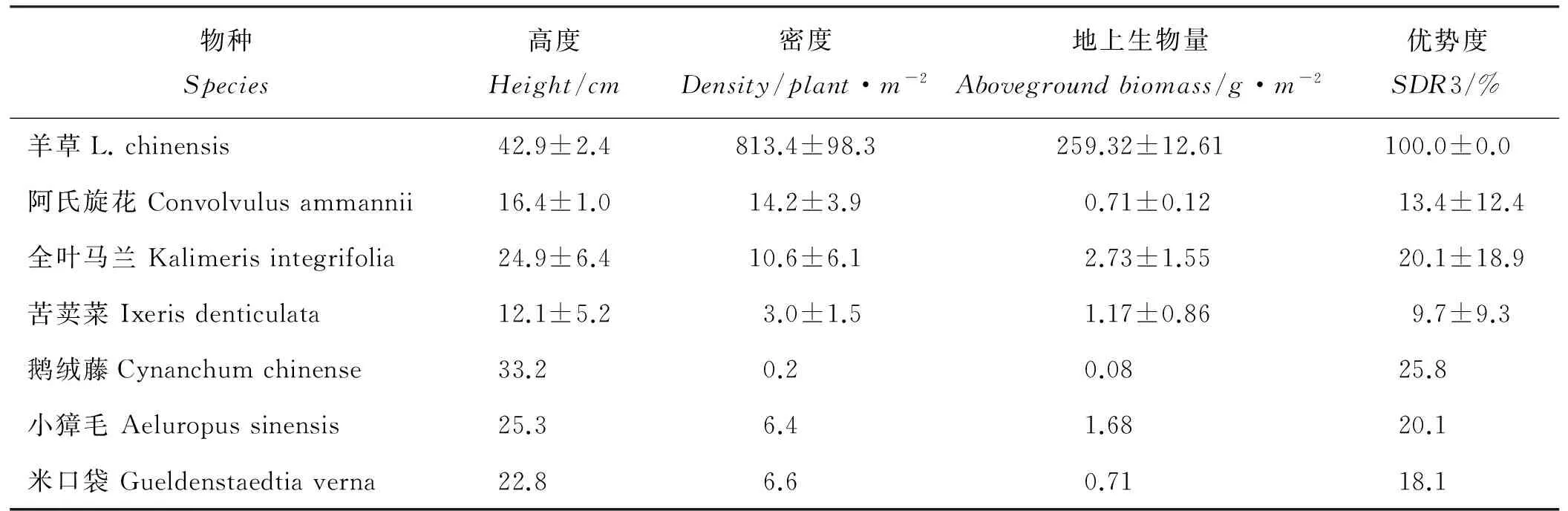
表1 研究地点植物群落特征(n=5)Table 1 The plant community characters at the study site(n=5)
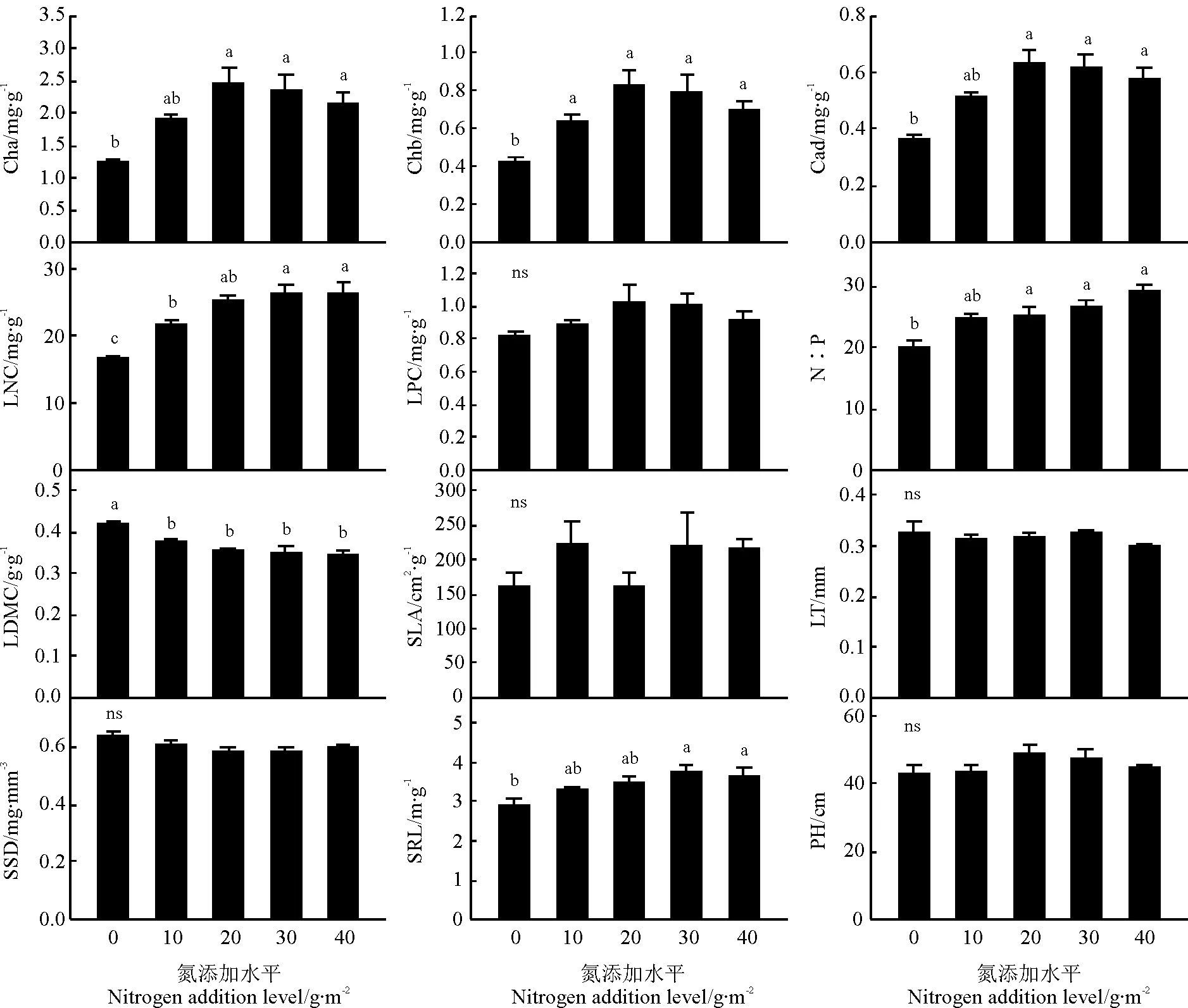
图1 羊草功能性状在不同氮添加水平间的多重比较Fig.1 Multiple comparisons of functional traits of L. chinensis among different nitrogen addition levels
注:Cha,叶片叶绿素a含量;Chb,叶片叶绿素b含量;Cad,叶片类胡萝卜素含量;LNC,叶片氮含量;LPC,叶片磷含量;LDMC,叶片干物质含量;SLA,比叶面积;LT,叶片厚度;SSD,比茎重;SRL,比根长;PH,植株高度。不同小写字母表示不同氮添加水平间差异显著(P<0.05),ns表示差异不显著(P>0.05)。下同。
Note:Cha,leafchlorophyllacontent;Chb,leafchlorophyllbcontent;Cad,leafcarotenoidcontent;LNC,leafnitrogencontent;LPC,leafphosphoruscontent;LDMC,leafdrymattercontent;SLA,specificleafarea;LT,leafthickness;SSD,stemspecificdensity;SRL,specificrootlength;PH,plantheight.Differentlowercaselettersmeansignificantdifferencesamongdifferentnitrogenadditionlevelsat0.05level,andnsmeansnosignificant.Thesamebelow.
叶绿素a与叶绿素b及类胡萝卜素呈极显著正相关(P<0.01),相关系数均大于0.95(表2)。叶片氮、磷含量呈显著正相关(P<0.05),叶片氮含量与氮磷比例也呈显著正相关,但氮磷比与叶片磷含量无显著相关(P>0.05)。叶片干物质含量与比叶面积、叶片厚度无显著关系(P>0.05),与比茎重呈极显著正相关(P<0.01),与其它性状呈显著负相关(P<0.05)。比叶面积与植株高度呈显著负相关(P<0.05),与其它性状无显著关系(P>0.05)。叶片厚度与其它性状均不相关(P>0.05)。比茎重与叶绿素a、叶绿素b、类胡萝卜素、叶片氮含量及叶片磷含量呈显著负相关(P<0.05),比根长与这些性状呈极显著正相关(P<0.01),植株高度与这些性状不相关(P>0.05) (表2)。
氮添加对羊草地上生物量有显著影响(P<0.05),随着氮添加量的增加,羊草地上生物量呈现先增加后减少的趋势,在20g·m-2处理达到最高,为381.67g·m-2,显著高于对照(P<0.05),其它氮添加处理与对照无显著差异(P>0.05)(图2)。
氮磷比、比叶面积、叶片厚度和比根长与羊草地上生物量之间无显著相关关系(P>0.05);羊草地上生物量随叶片干物质含量增加而显著减少(P<0.05),随比茎重增加而极显著减少(P<0.01),随叶片叶绿素a、叶绿素b、类胡萝卜素、叶片氮、磷含量和植株高度增加而极显著增加(P<0.01)(图3)。
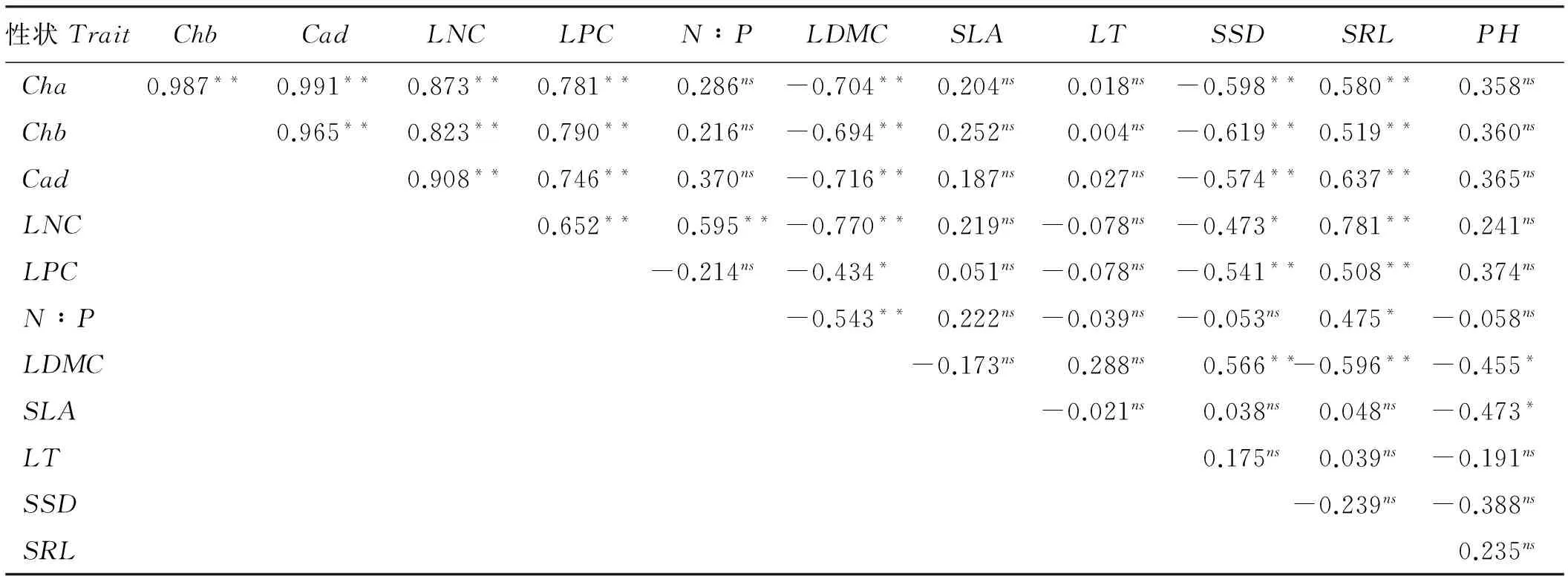
表2 羊草功能性状间的相关性Table 2 Correlation coefficient among functional traits of L. chinensis
注: **表示极显著相关(P<0.01),*表示显著相关(P<0.05),ns表示不相关(P>0.05)。
Note: **and*meanssignificantcorrelationat0.01and0.05level,respectively,andnsmeansnosignificantcorrelation.
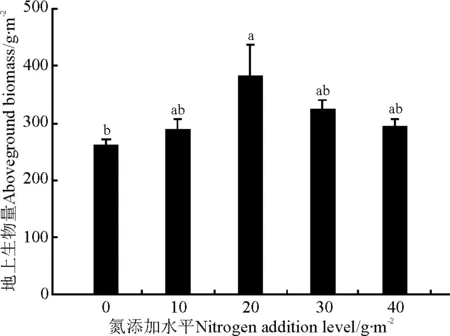
图2 氮添加对羊草地上生物量的影响Fig.2 Effect of nitrogen addition on abovegroundbiomass of L. chinensis
3讨论
植物功能性状的变化反映了植物在变化环境下的生存策略,包括对资源的获取、利用、储存及适应能力等[17]。研究结果显示,短期的氮添加主要影响羊草的生理性状,如叶绿素a、叶绿素b、类胡萝卜素和叶片氮含量随着氮添加量的增加而增加,这些功能性状主要与植物的光合作用相关。这说明氮添加缓解了植物对养分的竞争,使物种对资源的竞争从地下部分转向地上部分,即增加植物对光资源的竞争[1]。有意思的是,氮添加对羊草的结构性状无显著影响,如比叶面积、叶片厚度等在不同氮添加处理间没有表现出显著差异,这与其它研究并不一致[5]。这可能因为本研究时间较短,而植物性状随着资源水平的改变,首先是调整生理性状使其快速适应,而结构性状相对滞后,需要较长的时间才能发生显著变化。
叶片的氮、磷含量反映了植物对土壤养分的吸收特性。本研究中氮添加显著增加了叶片氮含量,而对叶片磷含量没有显著作用,因而氮磷比随着氮添加量的增加而增加,表明氮添加显著提高了土壤氮的有效性而对磷的影响较小[18]。Güsewell[19]综述了大量研究结果后认为,氮磷比>20和<10作为植物氮磷限制的评价指标,即氮磷比>20说明植物生长受磷素限制,<10说明植物生长受氮素限制,氮磷比在10~20说明植物生长与氮磷关系不明确。本研究中,在不施氮条件下,氮磷比为20,而随着氮添加的增多,氮磷比高于20,表明羊草的生长随着土壤中氮素的增多,磷素逐渐成为限制因子。因此,在施氮肥的同时,需要适当增加土壤中磷素的含量,以保持植物在土壤中可利用资源的平衡。
随着氮添加的增多,羊草的功能性状值并非一成不变地增加或减少,有的性状会出现单峰模式,如叶绿素a、叶绿素b的含量,这与万宏伟等[5]的研究结果一致,说明过量的氮也会抑制植物的生长,不利于植物对资源的利用[3]。因为过多的氮添加使草原土壤酸化和铵离子浓度增加,对植物造成毒害,不利于植物生长[20]。

图3 羊草地上生物量和功能性状的关系Fig.3 The relationships between aboveground biomass and functional traits of L. chinensis
量化植物功能性状之间的关系,特别是多个物种性状间的关系在过去20年中逐渐受到生态学家和生理学家的关注,因为它提供了不同物种间主要变异程度的信息,显示其对环境异质性的适应或者对生态系统功能的作用能力[21-23]。而同一物种在不同环境条件性状之间的相关性则能表现性状之间的权衡,反映物种对资源的利用情况[5]。本研究显示,叶绿素a、叶绿素b、类胡萝卜素、叶片氮、磷含量等性状之间存在极显著正相关关系,这反映了植物叶片的光合特征及对光的捕获能力。叶片色素含量越高,叶片氮、磷含量也越高,光合能力越强。比根长和叶片光合性状之间也呈正相关,说明氮添加条件下,根系从土壤中吸收更多的营养,根系生长速度加快,进而促进植物地上部分更快生长,植物光合能力更强。比茎重与叶片光合性状之间呈负相关,说明比茎重越小,茎生长越快,支持叶片获取更多的光资源,提高光合能力。
性状作为研究的解释变量是功能生态学研究的主要内容之一,植物性状对生态系统功能的变化有显著的指示作用[24-25]。“质量比假说(massratiohypothesis)”认为物种对生态系统功能的贡献与物种的多度有关,即功能性状对生态系统功能的影响与物种丰富度有关[26]。因此,优势种的功能性状决定了生态系统功能。在松嫩草地,羊草由于其根茎密织、耐践踏、竞争能力强,常形成大面积的纯群落[9]。本研究区域,羊草的密度和地上生物量均占到群落的95%以上,因此,羊草功能性状的变化与生态系统功能的关联有助于理解植物功能性状在影响生态过程中的作用。本研究表明,除了比叶面积、叶片氮磷比、比根长和叶片厚度之外,其它性状都能很好地预测羊草地上生物量的变化。因此,对羊草占绝对优势的松嫩草地建立易测的性状与地上生物量的关系,对草地管理有重要意义,如叶绿素含量,可以通过SPAD-502(日本,大阪)容易、快速地测量[27],且与羊草的地上生物量呈极显著正相关。
4结论
短期氮添加显著影响羊草的叶片光合生理性状,如叶绿素a和b含量、叶片氮含量等,而对羊草的结构性状如植株高度没有显著作用;而且生理性状之间表现出显著的相关性,而比叶面积、叶片厚度和植株高度等结构性状与其它功能性状相关性不显著;氮添加促进羊草地上生物量的提高,并在20g·m-2的氮添加处理下达到最高;氮添加条件下,羊草的功能性状可用于预测羊草地上生物量的变化。
参考文献References:
[1]BobbinkR,HicksK,GallowayJ,SprangerT,AlkemadeR,AshmoreM,BustamanteM,CinderbyS,DavidsonE,DentenerF,EmmettB,ErismanJW,FennM,GilliamF,NordinA,PardoL,deVriesW.Globalassessmentofnitrogendepositioneffectsonterrestrialplantdiversity:Asynthesis.EcologicalApplications,2010,20(1):30-59.
[2]潘庆民,白永飞,韩兴国,杨景成.氮素对内蒙古典型草原羊草种群的影响.植物生态学报,2005,29(2):311-317.
PanQM,BaiYF,HanXG,YangJC.EffectofnitrogenadditionsonaLeymus chinensispopulationintypicalsteppeofInnerMongolia.ChineseJournalofPlantEcology,2005,29(2):311-317.(inChinese)
[3]董晓兵,郝明德,郭胜安,石学军,马甜,刘盼盼,刘公社.施肥对羊草产量和品质的影响.草业科学,2014,31(10):1935-1942.
DongXB,HaoMD,GuoSA,ShiXJ,MaT,LiuPP,LiuGS.FertilizationeffectsonhayyieldandqualityofLeymus chinensis.PrataculturalScience,2014,31(10):1935-1942.(inChinese)
[4]李德军,莫江明,方运霆,蔡锡安,薛璟花,徐国良.模拟氮沉降对三种南亚热带树苗生长和光合作用的影响.生态学报,2004,24(5):876-882.
LiDJ,MoJM,FangYT,CaiXA,XueJH,XuGL.EffectsofsimulatednitrogendepositionongrowthandphotosynthesisofSchima superba,Castanopsis chinensisandCryptocarya concinnaseedlings.ActaEcologicaSinica,2004,24(5):876-882.(inChinese)
[5]万宏伟,杨阳,白世勤,徐云虎,白永飞.羊草草原群落 6 种植物叶片功能特性对氮素添加的响应.植物生态学报,2008,32(3):611-621.
WanHW,YangY,BaiSQ,XuYH,BaiYF.VariationsinleaffunctionaltraitsofsixspeciesalonganitrogenadditiongradientinLeymus chinensissteppeinInnerMongolia.ChineseJournalofPlantEcology,2008,32(3):611-621.(inChinese)
[6]BaiYF,WuJG,ClarkCM,NaeemS,PanQM,HuangJH,ZhangLX,HanXG.Tradeoffsandthresholdsintheeffectsofnitrogenadditiononbiodiversityandecosystemfunctioning:EvidencefromInnerMongoliagrasslands.GlobalChangeBiology,2010,16(1):358-372.
[7]章志龙,刘旭东,贾鹏,杜国祯.氮添加对高寒草甸植物花期物候和群落结构的影响.草业科学,2013,30(5):728-735.
ZhangZL,LiuXD,JiaP,DuGZ.Effectsofnitrogenadditiononfloweringphenology,functionaltraitsandcommunitystructureofalpinemeadowintheEasternQinghai-TibetPlateau.PrataculturalScience,2013,30(5):728-735.(inChinese)
[8]李晓刚,朱志红,周晓松,袁芙蓉,樊瑞俭,许曼丽.刈割,施肥和浇水对高寒草甸物种多样性,功能多样性与初级生产力关系的影响.植物生态学报,2011,35(11):1136-1147.
LiXG,ZhuZH,ZhouXS,YuanFR,FanFJ,XuML.Effectsofclipping,fertilizingandwateringontherelationshipbetweenspeciesdiversity,functionaldiversityandprimaryproductivityinalpinemeadowofChina.ChineseJournalofPlantEcology,2011,35(11):1136-1147.(inChinese)
[9]郑慧莹,李建东.松嫩平原的草地植被及其利用保护.北京:科学出版社,1993.
ZhengHY,LiJD.TheGrasslandVegetationandItsUtilizationandConservationontheSongnenPlain.Beijing:SciencePress,1993.(inChinese)
[10]杨金,刘扬,李楠,丁桂云.施肥对羊草矿质养分和产量的影响研究.土壤通报,1992,23(2):64-66.
YangJ,LiuY,LiN,DingGY.TheinfluenceoffertilizationonmineralnutrientsandyieldofLeymus chinensis.ChineseJournalofSoilScience,1992,23(2):64-66.(inChinese)
[11]李建东,刘建新.羊草草原施用氮肥的研究初报.中国草地学报,1981(3):58-59,35.
LiJD,LiuJX.PrimaryreportonresearchofnitrogenfertilizeronLeymus chinensisgrassland.ChineseJournalofGrassland,1981(3):58-59,35.(inChinese)
[12]尤英豪.羊草草地施肥效果探究.吉林林业科技,2005,34(1):38-40.
YouYH.StudyontheeffectsoffertilizerapplicationingrasslandofLeymus chinensis.JilinForestryScienceandtheTechnology,2005,34(1):38-40.(inChinese)
[13]WangJ,XieJ,ZhangY,GaoS,ZhangJ,MuC.Methodstoimproveseedyieldofbasedonnitrogenapplicationandprecipitationanalysis.AgronomyJournal,2010,102(1):277-281.
[14]刘思雅.模拟增温与施氮对羊草草原植物群落结构及生物量的影响.长春:东北师范大学硕士论文,2009.
LiuSY.EffectsofsimulatedwarmingandnitrogenapplicationonthestructuresandbiomassofplantcopnrnunityinLeymus chinensisgrassland.MasterThesis.Changchun:NortheastNormalUniversity,2009.(inChinese)
[15]鲍士旦.土壤农化分析.第三版.北京:中国农业出版社,2005.
BaoSD.AnalysisofSoilCharacteristics.3rd.Beijing:ChineseAgriculturalPress,2005.(inChinese)
[16]王英典,刘宁.植物学实验指导.北京:高等教育出版社,2001:56-57.
WangYD,LiuN.ExperimentalGuidanceforPlantBiology.Beijing:HigherEducationPress,2001:56-57.(inChinese)
[17]PywellRF,BullockJM,RoyDB,WarmanL,WalkerKJ,RotheryP.Planttraitsaspredictorsofperformanceinecologicalrestoration.JournalofAppliedEcology,2003,40(1):65-77.
[18]黄菊莹,袁志友,李凌浩.羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化.植物生态学报,2009,33(3):442-448.
HuangJY,YuanZY,LiLH.ChangesinN,PandspecificleafofgreenleavesofLeymus chinensisalongnitrogen,phosphorusandwatergradients.ChineseJournalofPlantEcology,2009,33(3):442-448.(inChinese)
[19]GüsewellS.N∶Pratiosinterrestrialplants:Variationandfunctionalsignificance.NewPhytologist,2004,164(2):243-266.
[20]ZhangY,LyuX,IsbellF,StevensC,HanX,HeN,ZhangG,YuQ,HuangJ,HanX.RapidplantspecieslossathighratesandatlowfrequencyofNadditionintemperatesteppe.GlobalChangeBiology,2014,20(11):3520-3529.
[21]CornelissenJHC,LavorelS,GarnierE,DíazS,BuchmannN,GurvichDE,ReichPB,terSteegeH,MorganHD,vanderHeijdenMGA,PausasJG,PoorterH.Ahandbookofprotocolsforstandardisedandeasymeasurementofplantfunctionaltraitsworldwide.AustralianJournalofBotany,2003,51(4):335-380.
[22]ReichPB,OleksynJ.GlobalpatternsofplantleafNandPinrelationtotemperatureandlatitude.ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica,2004,101(30):11001-11006.
[23]WrightIJ,GroomPK,LamontBB,PootP,PriorLD,ReichPB,SchulzeED,VeneklaasEJ,WestobyM.LeaftraitrelationshipsinAustralianplantspecies.FunctionalPlantBiology,2004,31(5):551-558.
[24]MokanyK,AshJ,RoxburghS.Functionalidentityismoreimportantthandiversityininfluencingecosystemprocessesinatemperatenativegrassland.JournalofEcology,2008,96(5):884-893.
[25]GarnierE,CortezJ,BillèsG,NavasML,RoumetC,DebusscheM,LaurentG,BlanchardA,AubryD,BellmannA.Plantfunctionalmarkerscaptureecosystempropertiesduringsecondarysuccession.Ecology,2004,85(9):2630-2637.
[26]GrimeJ.Benefitsofplantdiversitytoecosystems:Immediate,filterandfoundereffects.JournalofEcology,1998,86(6):902-910.
[27]BakkerMA,Carreo-RocabadoG,PoorterL.Leafeconomicstraitspredictlitterdecompositionoftropicalplantsanddifferamonglandusetypes.FunctionalEcology,2011,25(3):473-483.
(责任编辑武艳培)
DOI:10.11829/j.issn.1001-0629.2016-0017
*收稿日期:2016-01-11接受日期:2016-04-05
基金项目:国家自然科学基金项目(31500366、31470504);中央高校基本科研业务费专项资金资助项目(DC201501070402);人力资源和社会保障部2015年度留学人员科技活动择优资助项目
通信作者:周道玮(1963-),男,山东青岛人,研究员,博士,主要从事草地生态学研究。E-mail:zhoudaowei@iga.ac.cn
中图分类号:S543+.9;Q945.79
文献标志码:A
文章编号:1001-0629(2016)7-1383-08*
Corresponding author:Zhou Dao-weiE-mail:zhoudaowei@iga.ac.cn
ResponseofLeymus chinensisfunctionaltraitsandabovegroundbiomasstonitrogenadditioninSongnengrasslandinnortheastChina
SongYan-tao1,LiQiang2,WangPing3,ZhouDao-wei2,Wuyunna1
(1.CollegeofEnvironmentandResources,DalianMinzuUniversity,Dalian116600,China;2.NortheastInstituteofGeographyandAgroecology,ChineseAcademyofSciences,Changchun130102,China;3.SchoolofEnvironment,NortheastNormalUniversity,Changchun130117,China)
Abstract:The adaptive strategy of plant functional traits to resources change is one of the basic problems in ecology. In the present study, 11 plant functional traits and aboveground biomass of Leymus chinensis with different amount of nitrogen addition were determined. The results showed that nitrogen addition had significant effects on leaf chlorophyll a (Cha), chlorophyll b (Chb) and carotenoid (Cad) content, leaf nitrogen concentration (LNC), leaf dry matter content (LDMC), and specific root length (SRL), however, had no effects on leaf phosphorus concentration (LPC), specific leaf area (SLA), leaf thickness (LT), stem specific density (SSD), and plant height (PH). There were significant correlation between Cha, Chb, Cd, LNC, LDMC, SSD and SRL. SLA, LT and PH had no significant correlation with other functional traits. The aboveground biomass of L. chinensis was the highest at 20 g·m-2nitrogen addition. The aboveground biomass significantly increased with Cha, Chb, Cad, LNC, LPC, and PH increasing, but significantly decreased with LDMC and SSD increasing.
Key words:nitrogen addition; Leymus chinensis; leaf traits; Songnen grassland
宋彦涛,李强,王平,周道玮,乌云娜.羊草功能性状和地上生物量对氮素添加的响应.草业科学,2016,33(7):1383-1390.
SongYT,LiQ,WangP,ZhouDW,Wuyunna.ResponseofLeymus chinensisfunctionaltraitsandabovegroundbiomasstonitrogenadditioninSongnengrasslandinnortheastChina.PrataculturalScience,2016,33(7):1383-1390.
第一作者:宋彦涛(1982-),男,河南许昌人,讲师,博士,主要从事草地植物群落生态学研究。E-mail:yantaosong@dlnu.edu.cn
