模拟复合盐碱胁迫对芒幼苗生理特性的影响
2016-08-02徐鹏飞刘长乐周蕴薇
何 淼,王 欢,徐鹏飞,刘长乐,周蕴薇
(1.东北林业大学园林学院,黑龙江 哈尔滨 150040; 2.威海绿苑园林工程有限公司,山东 威海 264200; 3.黑龙江省林业科学院,黑龙江 哈尔滨 150040)
模拟复合盐碱胁迫对芒幼苗生理特性的影响
何 淼1,王 欢1,徐鹏飞2,刘长乐3,周蕴薇1
(1.东北林业大学园林学院,黑龙江 哈尔滨 150040; 2.威海绿苑园林工程有限公司,山东 威海 264200; 3.黑龙江省林业科学院,黑龙江 哈尔滨 150040)
摘要:土壤盐碱化在世界范围内普遍存在,日益严重的盐碱化威胁着植物的生长发育,致使植物种类减少。芒(Miscanthus sinensis)作为能源植物具有良好的经济效益和生态效益,本研究模拟我国东北大庆盐碱地的低(4.147 5、8.295 0 g·L-1)、中(12.442 5 g·L-1)、高(16.590 0、20.737 5 g·L-1)浓度土壤环境,以引种自辽宁省本溪阿家岭的芒为研究对象,对其幼苗的各项生理指标进行研究。结果发现,芒幼苗在受到复合盐碱胁迫处理时,未胁迫对照组各指标随时间的延长变化不大;低浓度处理时芒幼苗体内叶绿素含量随着胁迫时间的延长呈现出先升高后降低的趋势,中、高浓度处理时则不断降低;而相对电导率和丙二醛(MDA)含量则不断升高;超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)、谷胱甘肽还原酶(GR)活性和抗坏血酸(ASA)、谷胱甘肽(GSH)含量以及根活力一般在低浓度的复合盐碱处理时缓慢升高,高浓度处理时则呈现出先升高后降低的趋势。本研究基本界定了芒的复合盐碱耐受范围,可以为今后芒类能源植物的耐盐碱性筛选和在园林中的应用提供理论依据。
关键词:芒;幼苗;复合盐碱胁迫;生理特性
土壤盐碱化在世界范围内普遍存在,尤其是在较干旱的地区,盐碱化问题更为严重。据不完全统计,世界范围内的盐碱地面积大约为9.54亿hm2,而我国盐碱地面积大约是670万hm2,占到耕地总面积的7%左右。有些地方土地盐碱化程度过高,严重抑制了植物的生长发育,土地几乎不能使用[1]。大庆市位于松嫩平原腹地,是我国重要的石油城,而松嫩平原是我国北方土壤盐渍化最严重的区域之一[2],盐碱地面积占全国盐碱地总面积的一半以上[3],盐碱化程度日益严重。盐碱胁迫引起植物生理干旱[4-5],危害植物组织[6-7],影响植物的正常营养[8],引发氨的积累[7],并影响植物的气孔关闭[9]。随盐浓度的增加,番茄(Lycopersiconesculentum)每株叶片数、枝重、株重、根长和根表面积等明显下降[10]。如何改良盐碱化土壤、合理地将盐碱地利用起来,成为目前亟需解决的问题,而利用、筛选耐盐碱植物是最经济有效的措施之一。
芒(Miscanthussinensis)是禾本科(Gramineae)芒属的多年生草本C4能源植物[11],是极具开发潜力的能源植物之一,引起了很多国家,尤其是欧美国家的关注[12-13]。芒类植物作为能源植物开发具有干物质产量较高[14]、生物质质量较好[15]、种植成本低[16]、适应性强、生态效益好的优势。并且芒类植物的抗性较强,病虫害较少,栽培养护管理方面较为方便。据统计芒类植物的地下根茎能够在-20 ℃安全越冬,而叶片在5 ℃的环境下仍能正常生长[17]。芒属植物不存在生殖隔离,容易培育出新品种[18],在我国各地均有分布,但主要分布于我国东部地区以及南方各省。但是不同的种在其分布上也是有差异的,例如川芒(M.szechuanensis)基本上只生长在四川地区。芒属植物大都喜欢阳光充足的环境,在开阔的地段,生长势旺盛[19]。同其它植物类似,芒类植物可以通过土壤水分对矿物质的吸收进行调控[20-22]。芒类植物一般都有地下横走茎,根系较为发达,抗旱性强[23]。芒还因为其独特的美感,在城市园林中已经得到广泛应用,目前还越来越多的将其应用在专类园之中,比如岩石园、屋顶花园和禾本园等[24]。本研究以能源植物芒作为研究对象,模拟我国东北大庆盐碱地的低、中、高浓度土壤环境,对芒幼苗进行复合盐碱胁迫处理,并进行芒幼苗耐盐碱性的综合评价,以期为今后芒类能源植物的耐盐碱性筛选和在园林中的应用提供理论依据。
1材料与方法
1.1试验材料
本研究所用的中国芒种子材料采于2012年10月辽宁省本溪市阿家岭(41°22.803′ N,121°21.410′ E),种子采完后保存于东北林业大学园林学院种子储藏柜中,备用。
2013年5月将芒的种子播于穴盘中,放于东北林业大学园林学院110室光照培养箱内进行初期育苗,1个月后移栽于10 cm×10 cm的培养钵中,培养基质按照壤土∶蛭石∶河沙=3∶1∶1的质量比例配置,并将苗搬运到园林学院苗圃温室内继续培养1年。
2014年5月,在培养1年后的芒幼苗中选取生长健壮、长势一致的幼苗进行复合盐碱胁迫处理。胁迫处理之前,将培养钵的排水孔堵住,确保复合盐碱液不外流。试验共分为5个处理组(浓度1、2、3、4、5)和1个对照组(CK),处理组分别用不同浓度的复合盐碱液浇灌(表1),每次200 mL,每5 d浇灌一次,对照组(CK)用等量的蒸馏水灌溉。每组处理60盆,分别在第0、5、10、15、20、25天测定复合盐碱胁迫对芒幼苗各项生理指标的影响。低浓度为浓度1、浓度2,中浓度为浓度3,高浓度为浓度4、浓度5。
1.3指标测定及方法
叶绿素含量采用乙醇浸提法测定[25],相对电导率采用电导率仪法测定[26],丙二醛(MDA)含量采用硫代巴比妥酸(TBA)显色法测定[25],超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)法测定[25],过氧化物酶(POD)活性采用愈创木酚比色法测定[25],过氧化氢酶(CAT)酶活性参照Kato和Shimizu[27]的方法测定,抗坏血酸(ASA)含量参照Hodges等[28]的方法测定,谷胱甘肽(GSH)含量参照Griffith[29]的方法测定,抗坏血酸过氧化物酶(APX)的参照 Nakano和Asada[30]的方法测定,谷胱甘肽还原酶(GR)参照Grace和Logan[31]的方法测定,根活力采用TTC法测定[25]。

表1 复合盐碱溶液的成分Table 1 Composition of mixed saline alkali liquor
1.4数据分析
用Excel 2007软件对所有试验数据进行初步整理并作图,采用SPSS 19.0软件进行数据处理和统计分析。
2结果
2.1复合盐碱胁迫对芒幼苗叶绿素(chl)含量的影响
随着复合盐碱胁迫时间的延长,在浓度1、2处理时,芒幼苗中叶绿素含量呈先升高后下降的变化趋势,与CK相比,差异显著(P<0.05);其它各浓度处理,叶绿素含量均随着复合盐碱浓度的升高而降低,与对照组差异显著(P<0.05)(图1)。
2.2复合盐碱胁迫对芒幼苗相对电导率的影响
行政执法机关在依法查处违法行为过程中,发现违法事实涉及的金额、情节、后果等涉嫌犯罪,需要将案件移送有关司法机关处理,在移送过程中就产生了行政执法机关与刑事司法机关的衔接问题。水行政执法属于行政执法的一种,研究水行政执法与刑事司法“两法衔接”机制,必须以理清行政执法与刑事司法 “两法衔接”机制概念为前提。在理论层面,我国许多学者从不同角度对 “两法衔接”机制内涵作了深刻阐释。
随着复合盐碱胁迫时间的延长,芒幼苗相对电导率均呈现出不断升高的趋势(图2)。在浓度1处理下,芒幼苗相对电导率一直保持平稳升高,且与CK相差不大,说明浓度1下的复合盐碱胁迫处理,几乎不会对芒幼苗的膜系统产生危害。但是随着浓度不断增大,胁迫时间的不断延长,芒相对电导率的升高趋势越来越明显。浓度5处理的后期,芒幼苗膜系统已受到严重伤害,芒幼苗膜系统会被彻底破坏,会导致其植株死亡。

图1 复合盐碱胁迫对芒幼苗叶绿素(chl)含量的影响Fig.1 Effects of salt and alkali mixture stress on chlorophyll content in seedling of Miscanthus sinensis
注:不同小写母表示相同处理时间下不同浓度处理之间差异显著(P<0.05)。下同。
Note:Different lower case letters show significant difference among different concentration treatment groups in the same processing time at 0.05 level.The same below.

图2 复合盐碱胁迫对芒幼苗相对电导率的影响Fig.2 Effects of salt and alkali mixture stress on relative electric conductivity in seedling of Miscanthus sinensis
2.3复合盐碱胁迫对芒幼苗MDA含量的影响
各复合盐碱浓度处理下,芒幼苗中MDA含量均随着时间的延长有所升高,但是升高的幅度不同(图3)。在浓度1、2处理下,芒幼苗中MDA含量均与CK相差不大,说明此时复合盐碱胁迫对芒幼苗的膜脂过氧化影响很小。浓度3处理时,芒幼苗中MDA含量同样与CK相差不大,说明此时芒幼苗的膜脂过氧化已经开始,但是影响也不大。浓度4、5处理下,芒体内MDA含量均迅速升高,与CK差异性显著(P<0.05),说明此时芒幼苗的膜脂质过氧化程度已经很严重,超过了它的承受极限,会导致植株死亡。
2.4复合盐碱胁迫对芒幼苗保护酶系统的影响
2.4.1复合盐碱胁迫对芒幼苗超氧化物岐化酶(SOD)活性的影响随着复合盐碱胁迫时间的延长,各个浓度处理下的芒的SOD活性呈现出不同的变化趋势(图4)。芒幼苗在浓度1、2处理下,SOD活性随着胁迫时间的延长,呈现出不断增强的趋势,而在浓度3、4、5处理下则呈现出先升高后降低的趋势。但同样在浓度5处理下的第25天,SOD活性显著低于对照组(P<0.05),说明此时芒幼苗体内负氧离子的积累,膜脂过氧化程度加剧,严重损伤了芒幼苗的膜系统,会使幼苗死亡。

图3 复合盐碱胁迫对芒幼苗MDA含量的影响Fig.3 Effects of salt and alkali mixture stress on MDA content in seedling of Miscanthus sinensis
2.4.2复合盐碱胁迫对芒幼苗过氧化物酶(POD)活性的影响随着时间的延长,浓度1、2、3处理下,芒幼苗中POD活性均呈现出不断升高的趋势,而浓度4、5处理下的芒幼苗中POD活性则呈现出先上升后下降的趋势(图5)。在浓度4、5处理时,芒幼苗的POD活性分别在第20天和第15天时达到最大值,且芒幼苗均在其处理的后期,体内POD活性明显低于CK,说明此时过氧化物的积累已经达到了芒幼苗的忍受极限,进一步则导致其植株死亡。
2.4.3复合盐碱胁迫对芒幼苗过氧化氢酶(CAT)活性的影响随着复合盐碱胁迫时间的延长,浓度1处理下,芒幼苗体内CAT的变化趋势和CK相似,浓度2处理下,呈现不断升高的变化趋势,而浓度3、4、5处理下,则会呈现先升高后降低的变化趋势(图6)。芒幼苗的CAT活性在浓度5处理的第25天明显著低于CK(P<0.05),说明此时高浓度的复合盐碱胁迫处理下,芒幼苗体内的CAT已经失去活性,丧失了对幼苗的保护能力,会造成植株死亡。
2.5复合盐碱胁迫对芒幼苗ASA-GSH循环相关非酶抗氧化物质含量的影响
2.5.1复合盐碱胁迫对芒幼苗抗坏血酸(ASA)含量的影响随着复合盐碱胁迫时间的延长,各浓度处理下的芒幼苗体内的ASA含量均呈现先上升后下降的趋势(图7)。浓度1处理下,芒幼苗体内的ASA含量在与CK相差不大。从浓度3处理开始,芒幼苗体内的ASA含量的升高降低幅度明显增大,但在第25天时,浓度4处理下的ASA含量基本与CK持平,而浓度5处理下的ASA含量略低于CK。说明浓度3、4、5处理下,芒幼苗已经受到很严重的损害。

图4 复合盐碱胁迫对芒幼苗超氧化物岐化酶(SOD)活性的影响Fig. 4 Effects of salt and alkali mixture stress on SOD activity in seedling of Miscanthus sinensis

图5 复合盐碱胁迫对芒幼苗过氧化物酶(POD)活性的影响Fig.5 Effects of salt and alkali mixture stress on POD activity in seedling of Miscanthus sinensis

图6 复合盐碱胁迫对芒幼苗过氧化氢酶(CAT)活性的影响Fig. 6 Effects of salt and alkali mixture stress on CAT activity in seedling of Miscanthus sinensis
2.5.2复合盐碱胁迫对芒幼苗谷胱甘肽(GSH)含量的影响随着复合盐碱胁迫时间的延长,芒幼苗体内的GSH含量均呈现先上升后下降的趋势,但是上升和下降的幅度不一样(图8)。浓度1处理下,芒幼苗体内的GSH含量总体来说与CK相差不大,说明此时芒幼苗植株没有受到伤害。而浓度5胁迫处理下,芒幼苗体内的GSH含量在第5天就开始迅速升高,并显著高于CK(P<0.05),在第10天下降趋势明显,在第15天,仍高于CK,而在第20、25天稍微低于CK。说明浓度5处理下的芒幼苗受到复合盐碱胁迫的损害程度较大,后期可能导致植株死亡。

图7 复合盐碱胁迫对芒幼苗抗坏血酸(ASA)含量的影响Fig.7 Effects of salt and alkali mixture stress on ASA content in seedling of Miscanthus sinensis

图8 复合盐碱胁迫对芒幼苗谷胱甘肽(GSH)含量的影响Fig.8 Effects of salt and alkali mixture stress on GSH content in seedling of Miscanthus sinensis
2.6复合盐碱胁迫对芒幼苗ASA-GSH循环相关抗氧化酶活性的影响
2.6.1复合盐碱胁迫对芒幼苗抗坏血酸过氧化物酶(APX)活性的影响随着复合盐碱胁迫时间的延长,在浓度1、2处理下,APX活性呈现出不断上升趋势,但在浓度3、4、5处理下则呈现出先上升后下降的趋势(图9)。芒幼苗在浓度1、2处理时,其体内APX活性变化不是很大,但是在浓度4、5处理后期,APX活性均明显低于CK,说明浓度4、5处理的后期,复合盐碱胁迫对芒幼苗植株造成了严重的伤害,其体内的APX活性已经严重失活。
2.6.2复合盐碱胁迫对芒幼苗谷胱甘肽还原酶(GR)活性的影响随着复合盐碱胁迫时间的不断延长,浓度1胁迫处理下,GR活性呈现出上升趋势,而其它浓度处理下则均呈现先上升后下降的变化趋势(图10)。浓度1处理时,芒的GR活性变化不是很大,浓度2处理时,芒幼苗GR活性变化幅度仍然很小,但在浓度4、5处理前期,芒幼苗体内的GR活性迅速升高,后期,同样低于CK,说明浓度4、5处理的前期H2O2就迅速积累,促使芒幼苗体内的GR活性迅速升高,清除芒幼苗体内过多的H2O2,之后由于H2O2的积累量已经远远超过GR的最大清除能力,导致GR失去活性,直至芒幼苗植株死亡。
2.7复合盐碱胁迫对芒幼苗根活力的影响
随着复合盐碱胁迫时间的延长,浓度1、2处理下的芒幼苗根活力呈现出不断升高的趋势(图11)。而浓度3、4、5处理下,其根活力则呈现先升高后降低的趋势。浓度1、2处理下的芒幼苗根活力与CK相差不大,在浓度3、4、5处理时,变化幅度增大,且芒幼苗根活力在浓度5处理后期显著低于CK(P<0.05)。说明此时芒幼苗根系已经遭受严重破坏,会导致植株死亡。

图9 复合盐碱胁迫对芒幼苗抗坏血酸过氧化物酶(APX)的影响Fig.9 Effects of salt and alkali mixture stress on APX activity in seedling of Miscanthus sinensis

图10 复合盐碱胁迫对芒幼苗谷胱甘肽还原酶(GR)活性的影响Fig.10 Effects of salt and alkali mixture stress on GR activity in seedling of Miscanthus sinensis
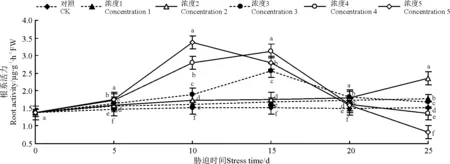
图11 复合盐碱胁迫对芒幼苗根活力的影响Fig.11 Effects of salt and alkali mixturestress on root activity in seedling of Miscanthus sinensis
3讨论与结论
在以往的研究中,胁迫指标多为NaCl[32],但盐碱地中盐害与碱害并存,单独的NaCl胁迫作用并不能代表真实情况。考虑到上述原因,本研究采用复合盐碱胁迫的方法对芒幼苗进行胁迫处理,因而在复合盐碱胁迫下,植物不仅要应对生理干旱和离子毒害,还必须应对高pH值的影响。
叶绿素是进行光合作用的物质基础,叶绿素含量的高低在一定程度上可以反映植物光合作用的强弱[33]。pH升高将导致植物生长介质中的金属离子沉淀从而失去有效性,其中受影响较为严重的金属离子是Fe、Mn、Mg、Ca等[34],而Fe和Mg是叶绿素合成时的必需元素,缺乏将导致叶绿素含量降低。盐胁迫对叶绿素含量的影响因盐类型、处理浓度、植物种类等因素的不同而有所不同,分别有下降[35]、上升[36]或先升高后降低的趋势[37]。在本研究中,复合盐碱胁迫对芒幼苗的光合作用影响很大,低浓度处理时芒幼苗体内叶绿素含量随胁迫时间的延长呈现出先升高后降低的趋势,中、高浓度处理时则不断降低,低浓度盐碱胁迫下芒幼苗体内的叶绿素含量之所以会先升后降,主要是因为复合盐碱胁迫前期,盐碱液中的Mg2+促进了叶绿素的合成,而后期胁迫程度加大,会对芒幼苗的细胞结构造成破坏,影响其光合作用。
由于离子毒害和渗透胁迫作用,盐胁迫会对植物的膜系统造成伤害[21]。盐胁迫易导致植物体内的活性氧和自由基大量积累,这时植物体内的保护酶系统就会对其进行清除,过高的盐碱胁迫会加剧对膜系统的伤害,甚至导致植物的程序性死亡[38]。在本研究中,随着胁迫时间的延长,芒体内活性氧和自由基的增加,造成膜脂过氧化伤害,各保护酶活性发生相应变化,且随着胁迫时间的延长和复合盐碱浓度的增大,这种破坏愈发严重。在低浓度复合盐碱胁迫处理时,芒幼苗体内的过氧化物和自由基积累,芒幼苗体内相对电导率,MDA含量,SOD、POD、CAT、APX、GR活性和ASA、GSH含量升高。在高浓度复合盐碱胁迫处理时,相对电导率和MDA含量依然呈现升高的趋势,而其它指标则不同。在高浓度处理初期,芒幼苗植株体内过氧化物的过量聚集,促进SOD、POD、CAT、APX、GR活性和ASA、GSH含量的急剧升高,而后期过氧化物的聚集量过大就会造成SOD、POD、CAT、APX、GR活性和ASA、GSH含量的迅速降低,导致芒幼苗植株死亡,这与对黄连(Coptischinensis)[39]、烟草(Nicotianatabacum)[40]、黄瓜(Cucumissativus)[41]的研究结果类似。
植物的根系是植物最重要的吸收合成器官,直接影响植物的生长。逆境下,植物体内过氧化物和自由基就会积累,加快植物根系衰老氧化,因此,植物的根活力大小将反映出植物体内过氧化物和自由基的含量。有研究表明,4种绣线菊(Spiraea)根系活力的变化随盐碱胁迫的加强先升高后下降,表明绣线菊对盐碱胁迫有一定的耐受能力,但超过一定范围时,根系受损严重导致根系活力下降[42]。本研究也得到类似结果,芒幼苗根系活力在低浓度的盐碱处理时呈现出缓慢升高的趋势,说明较低浓度的盐碱处理促进了根活力的提升,此时芒幼苗植株生长良好;随后为抵御外界的高浓度盐碱环境,根活力再次升高;最后根活力又急剧降低,说明芒幼苗根系已受到严重破坏,无法适应逆境环境,植株死亡。
综上所述,芒幼苗能够抵抗低、中浓度的复合盐碱胁迫,这与其自身的渗透物质调节、膜结构以及相关保护酶系统有着密切关系。芒幼苗通过自身相关保护酶系统和调节物质的积累来抵御来自外界的盐碱胁迫,清除体内过多活性氧和自由基,保护细胞结构不被破坏,存活较长时间,而高浓度的盐碱胁迫时,芒幼苗在处理前期还能够正常生长,但随着胁迫时间的延长,芒幼苗的细胞膜结构和保护酶系统会遭到破坏,造成生长调节机制紊乱,使植株不能正常生长。
大庆地区的土地盐碱化对植物造成的伤害主要是盐碱的过度积累造成土壤颗粒较为分散,土壤不断膨大化,造成土壤的透水性、相对含水量、吸水速度以及水力传导度相应降低,从而造成土壤理化性质恶化,土壤结构黏连、透气性变差、微生物活动较少、渗透系数变低,使植物处于生理干旱,导致其生长不良。大庆地区盐碱地的离子种类和pH值基本符合本研究低、中浓度的复合盐碱液浓度[2],芒幼苗能够适应低、中浓度的复合盐碱胁迫处理,说明芒幼苗基本能够适应大庆地区的土壤环境,且生长良好。
禾本科还有许多耐盐碱的植物,有研究发现,禾本科披碱草(Elymusdahuricus)是松嫩平原上耐盐碱性较强并最有代表性的优质牧草,其耐盐碱性明显强于豆科植物紫花苜蓿(Medicagosativa)[43];长穗冰草(Agropyronelongatum)在土壤pH≤10.5的苏打盐碱土上仍可以出苗和生长[44],是一种耐碱能力较强的植物;东北碱地常见牧草虎尾草(Chlorisvirgata)比羊草(Leymuschinensis)更耐盐碱[45],虎尾草在土壤pH 10.57、盐浓度高达300 mmol·L-1的高盐高碱条件下仍能存活,可以证明虎尾草具有很强的耐盐碱能力[46],一定程度上可能强于本研究的中国芒。另外,对禾本科植物种子的耐盐碱情况近年来也有一定的研究[47-48]。在本研究中,芒幼苗适应碱胁迫的临界点可能在pH为10左右,低于此点时,胁迫对芒幼苗影响相对较小;高于此点时,胁迫的作用明显增强。综合比较来看,芒幼苗具有较强的耐盐碱能力。
植物耐盐碱生理指标的评价和分析是一种较有效的提供选择耐盐碱植物的方法,要想系统地研究芒的耐盐碱性,不仅要考虑到芒的生长阶段、盐碱胁迫的时间,还要考虑不同个体间的差异。并且芒的耐盐碱性筛选主要是为了将来的园林应用,而具有园林观赏价值的主要是成苗期的花絮,本研究只是探究了复合盐碱胁迫对芒幼苗生长的影响,并没有深入研究复合盐碱胁迫是否影响其开花等其它生理过程,因此,仅凭实验室里面测定的几个生理指标进行芒的耐盐碱性评价是不全面的,有必要对芒做更全面的比较分析,才能得到其耐盐碱本质。
参考文献References:
[1]田福平,王锁民,郭正刚,张自和.紫花苜蓿脯氨酸含量和含水量、单株干质量与抗旱性的相关性研究.草业科学,2004,21(1):3-6.
Tian F P,Wang S M,Guo Z G,Zhang Z H.Relationship between proline content and water content,single plant dry matter,and drought resistance of alfalfa.Pratacultural Science,2004,21(1):3-6.(in Chinese)
[2]刘东兴,宫伟光.大庆盐碱地立地质量评价.中国水土保持科学,2009,7(3):98-103.
Liu D X,Gong W G.Evaluation of site quality for saline-alkali soil in Daqing.Science of Soil and Water Conservation,2009,7(3):98-103.(in Chinese)
[3]宋德成,洪影,于大永.松嫩平原盐碱地开发利用状况分析.东北水利水电,2014(9):21-22.
[4]张敏,蔡瑞国,李慧芝,李建敏,戴忠民,王振林,尹燕枰.盐胁迫环境下不同抗盐性小麦品种幼苗长势和内源激素的变化.生态学报,2008,28(1):310-320.
Zhang M,Cai R G,Li H Z,Li J M,Dai Z M,Wang Z L,Yin Y P.Responses of seedling growth and endogenous hormone contents in different wheat cultivars to salt stress.Acta Ecologica Sinica,2008,28(1):310-320.(in Chinese)
[5]阮海华.一氧化氮提高小麦幼苗耐盐性及其分子机制研究.南京:南京农业大学博士学位论文,2006.
Ruan H H.Mechanisms of nitric oxide conferring salt tolerance to wheat(TriticumaestivumL.) seedlings against salt stress.PhD Thesis.Nanjing:Nanjing Agricultural University,2006.(in Chinese)
[6]杨春武,李长有,尹红娟,鞠淼,石德成.小冰麦对盐胁迫和碱胁迫的生理响应.作物学报,2007,33(8):1255-1261.
Yang C W,Li C Y,Yin H J,Ju M,Shi D C.Physiological response of Xiaobingmai(Triticumaestivum-Agropyronintermedium) to salt-stress and alkali-stress.Acta Agronomica Sinica,2007,33(8):1255-1261.(in Chinese)
[7]商洪池,洪金祥.盐碱环境的改良措施及施工方法.河北林业科技,2003(4):29-32.
[8]夏阳,梁慧敏,王太明,束怀瑞,王清华,柴传华.盐胁迫对苹果器官中钙镁铁锌含量的影响.应用生态学报,2005,11(3):431-434.
Xia Y,Liang H M,Wang T M,Shu H R,Wang Q H,Chai C H.Effects of NaCl stress on Ca,Mg,Fe and Zn contents of different apple organs.Chinese Journal of Applied Ecology,2005,11(3):431-434.(in Chinese)
[9]段九菊,郭世荣,康云艳,李璟,刘香娥.盐胁迫对黄瓜幼苗根系生长和多胺代谢的影响.应用生态学报,2008,19(1):57-64.
Duan J J,Guo S R,Kang Y Y,Li J,Liu X E.Effect of salt strees on cucumber seedlings root growth and polyamine metabolism.Chinese Journal of Applied Ecology,2008,19(1):57-64.(in Chinese)
[10]Mohammad M,Shibli R,Ajlouni M,Nimri L.Tomato root and shoot responses to salt stress under different levels of phosphorus nutrition.Plant Nutrition,1998,21:1667-1680.
[11]Chen S L,Renvoize S A.Flora of China.Beijing:Science Press,2006:581-583.
[12]Pang J,Zheng M,Wang A,Sun R,Wang H,Yu J,Zhang T.Catalytic conversion of concentratedMiscanthusin water for ethylene glycol production.Aiche Journal,2014,60(6):2254-2262.
[13]Hastings A,Clifton-Brown J,Wattenbach M,Mitchell P,Stampfl P,Smith P.Future energy potential ofMiscanthusin Europe.Global Change Biology Bioenergy,2009,1(2):180-196.
[14]Heaton E A,Dohleman F G,Long S P.Meeting US biofuel goals with less land:The potential ofMiscanthus.Global Change Biology,2008,14(9):2000-2014.
[15]Lewandowski I,Scurlock J M O,Lindvall E,Lindvall E,Christou M.The development and current status of perennial rhizomatous grasses as energy crops in the US and Europe.Biomass and Bioenergy,2003,25(4):335-361.
[16]Wang D,Portis A R,Moose S P,Long S P.Cool C4photosynthesis:Pyruvate Pi dikinase expression and activity corresponds to the exceptional cold tolerance of carbon assimilation inMiscanthus×giganteus.Plant Physiology,2008,148(1):557-567.
[17]VanloockeA,Bernacchi C J,Twine T E.The impacts ofMiscanthus×giganteusproduction on the Midwest US hydrologic cycle.Global Change Biology Bioenergy,2010,2(4):180-191.
[18]朱明东.芒与五节芒种间自然杂交研究.长沙:湖南农业大学硕士学位论文,2011.
Zhu M D.Studies on natural hybridization betweenMiscanthussinensisandM.floridulus.Master Thesis.Changsha:Hunan Agricultural University,2011.(in Chinese)
[19]周昌弘,黄生,陈淑华.台湾芒属植物生态与演化.科学发展,2007,27(10):1158-1169.
Zhou C H,Huang S,Chen S H.Ecology and evolution ofMiscanthusin Taiwan.Scientific Development,2007,27(10):1158-1169.(in Chinese)
[20]毕银丽,丁保建,全文智,李晓林.VA菌根对白三叶吸收水分和养分的影响.草地学报,2001,9(2):154-158.
Bi Y L,Ding B J,Quan W Z,Li X L.Influence of VA mycorrhize on nutrient and water absorption in white clover.Acta Agrestia Sinica,2001,9(2):154-158.(in Chinese)
[21]陈晓远,高志红,刘晓英,罗远培.水分胁迫对冬小麦根、冠生长关系及产量的影响.作物学报,2004,30(7):723-728.
Chen X Y,Gao Z H,Liu X Y,Luo Y P.Effects of water stress on root/shoot relation and grain yield in winter wheat.Acta Agronomica Sinica,2004,30(7):723-728.(in Chinese)
[22]刘金祥,王铭铭,肖生鸿,刘家琼.干旱胁迫对香根草生长及光合生理主要特征的影响.四川草原,2005(3):28-30.
Liu J X,Wang M M,Xiao S H,Liu J Q.The responds of growth and photosynthesis ofVetiveriazizanioidesin condition of drought stress.Journal of Sichuan Grassland,2005(3):28-30.(in Chinese)
[23]解新明,周峰,赵燕慧,卢小良.多年生能源禾草的产能和生态效益.生态学报,2008,28(5):2329-2342.
Xie X M,Zhou F,Zhao Y H,Lu X L.A sunnary of ecological and energy-producing effects of perennial energy grasses.Acta Ecologica Sinica,2008,28(5):2329-2342.(in Chinese)
[24]任君霞.5种(品种)芒属观赏草的抗性研究及园林应用.杭州:浙江农林大学硕士学位论文,2012.
Ren J X.Study on resistance of fiveMiscanthusornamental grasses and the application in landscape.Master Thesis.Hangzhou:Zhejiang A & F University,2012.(in Chinese)
[25]李合生.植物生理生化试验原理和技术.北京:高等教育出版社,2000.
[26]陈建勋,王晓峰.植物生理学实验指导.第2版.广州:华南理工大学出版社,2006:64-66.
[27]Kato M,Shimizu S.Chlorophyll metabolism in higher plants.Ⅶ.Chlorophyll degradation in senescing tobacco leaves;phenolic-dependent peroxidative degradation.Canadian Journal of Botany,1987,65:729-735.
[28]Hodgcs D M,Andrews C J,Johnson D A,Hamilton R I.Antioxidant compound responses to chilling stress in differentially sensitive inbred maize lines.Plant Physiology,1996,98:685-692.
[29]Griffith O W.Determination of glutathione and glutathione disulfide using glutathione reductase and 2-vinylpyridine.Annual Review of Biochemistry,1980,106:207-212.
[30]Nakano Y,Asada K.Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts.Plant,Cell & Physiology,1981,22:867-880.
[31]Grace S C,Logan B A.Acclimation of foliar antioxidant systems to growth irradiance in three broad-leaved evergreen species.Plant Physiology,1996,112:1631-1640.
[32]汤章城.现代植物生理学实验指南.北京:中国科学技术出版社,1999:308.
[33]王伟华,张希明,闫海龙,梁少民,杨小林.盐处理对多枝怪柳光合作用和渗调物质的影响.干旱区研究,2009,26(4):561-568.
Wang W H,Zhang X M,Yan H L,Liang S M,Yang X L.Effects of salt stress on photosynthesis and osmo-regulation substances ofTamarixramosissimaLedeb.Arid Zone Research,2009,26(4):561-568.(in Chinese)
[34]Yang C,Chong J,Li C,Kim C,Shi D,Wang D.Osmotic adjustment and ion balance traits of an alkali resistant halophyteKochiasieversianaduring adaptation to salt and alkali conditions.Plant and Soil,2007,294:263-276.
[35]贾娜尔·阿汗,杨春武,石德成,王德利.盐生植物碱地肤对盐碱胁迫的生理响应特点.西北植物学报,2007,27(1):70-84.
Jianaer·Ahan,Yang C W,Shi D C,Wang D L.Physiological response of an alkali resistant halophyteKochiasieversianato salt and alkali stresses.Acta Botanica Boreali-Occidentalia Sinica,2007,27(1):70-84.(in Chinese)
[36]范希峰,侯新村,朱毅,武菊英.盐胁迫对柳枝稷苗期生长和生理特性的影响.应用生态学报,2012,23(6):1476-1480.
Fan X F,Hou X C,Zhu Y,Wu J Y.Impacts of salt stress on the growth and physiological characteristics ofPanicumvirgatumseedlings.Chinese Journal of Applied Ecology,2012,23(6):1476-1480.(in Chinese)
[37]徐静,董宽虎,高文俊,谢开云.NaCl和Na2SO4胁迫下冰草幼苗的生长及生理响应.中国草地学报,2011,33(1):36-41.
Xu J,Dong K H,Gao W J,Xie K Y.Growth and physiological responses ofAgropyroncristatumseedlings under NaCl and Na2SO4stress.Chinese Journal of Grassland,2011,33(1):36-41.(in Chinese)
[38]Katsuhara M,Kawasaki T.Salt stress induced nuclear and DNA degradation in meristematic cells of barley roots.Plant,Cell & Physiology,1996,37:169-173.
[39]武敬亮,田桂香,孙敏.盐碱胁迫和SNP对黄连膜脂过氧化及保护酶活性的影响.浙江林业科技,2006,26(5):5-9.
Wu J L,Tian G X,Sun M.Effect of salt-alkaline stress and SNP on membrane-lipid peroxidation and protective enzyme ofCoptischinensis.Journal of Zhejiang Forestry Science and Technology,2006,26(5):5-9.(in Chinese)
[40]陈展宇,崔喜艳,史岩玲.混合盐碱胁迫对3种类型烟草幼苗保护酶活性的影响.吉林农业大学学报,2006,28(5):486-489.
Chen Z Y,Cui X Y,Shi Y L.Effect of complex alkali-saline stress on activity of protective enzyme in three varieties of tobacco seedlings.Journal of Jilin Agricultural University,2006,28(5):486-489.(in Chinese)
[41]张丽平.盐碱胁迫对黄瓜种子发芽和幼苗生理代谢的影响.泰安:山东农业大学硕士学位论文,2008.
Zhang L P.Effect of salt and alkali stress on physiological metalbolism in germination and seedlings of cucumber.Master Thesis.Taian:Shandong Agricultural University,2008.(in Chinese)
[42]石溪婵.四种绣线菊根系对混合盐碱胁迫的生理响应研究.哈尔滨:东北农业大学硕士学位论文,2010.
Shi X C.Study on the physiological response in roots of 4 spiraeas to saline-alkali stress.Master Thesis.Harbin:Northeast Agricultual University,2010.(in Chinese)
[43]胡宗英.不同盐碱胁迫对披碱草和紫花苜蓿种子萌发的影响.长春:吉林农业大学硕士学位论文,2014.
Hu Z Y.The effects of different saline-alkaline stress on seed germination ofElymusdahuricusandMedicagosativa.Master Thesis.Changchun:Jilin Agricultural University,2014.(in Chinese)
[44]黄立华,梁正伟,王志春,杨福,陈渊.苏打盐碱胁迫对长穗冰草幼苗生长和K+、Na+含量的影响.中国草地学报,2006,28(5):60-65.
Huang L H,Liang Z W,Wang Z C,Yang F,Chen Y.Effect of saline-sodic stress on seedling growthand content of K+and Na+ofAgropyronelongatum.Chinese Journal of Grassland,2006,28(5):60-65.(in Chinese)
[45]鞠晓峰,李国良,杜广明,韩玉静,于开源.盐碱胁迫对非常规草坪草羊草和虎尾草生理生态特性的影响.东北农业大学学报,2011,42(11):155-160.
Ju X F,Li G L,Du G M,Han Y J,Yu K Y.Effect of saline-alkali tolerance physiological and ecological character onLeymuschinensisandChlorisvirgatain Songnen Plain.Journal of Northeast Agricultural University,2011,42(11):155-160.(in Chinese)
[46]尹红娟.虎尾草对盐碱混合胁迫的生理响应特点.长春:东北师范大学硕士学位论文,2008.
Yin H J.Physiological responses ofChlorisvirgateto various salt-alkaline mixed stress.Master Thesis.Changchun:Northeast Normal University,2008.(in Chinese)
[47]高战武,蔺吉祥,邵帅,盛后财,范春燕,屈吉.复合盐碱胁迫对燕麦种子发芽的影响.草业科学,2014,31(3):451-456.
Gao Z W,Lin J X,Shao S,Sheng H C,Fan C Y,Qu J.Effects of complex salt-alkali stresses on seed germination of oat.Pratacultural Science,2014,31(3):451-456.(in Chinese)
[48]纪荣花,于磊,鲁为华,艾尼娃尔·艾合买提.盐碱胁迫对芨芨草种子萌发的影响.草业科学,2011,28(2):245-250.
Ji R H,Yu L,Lu W H,Ainiwaer·Aihemaiti.Effects of saline-alkaloid stress on the seed germination ofAchantherumsplendens.Pratacultural Science,2011,28(2):245-250.(in Chinese)
(责任编辑武艳培)
DOI:10.11829/j.issn.1001-0629.2015-0632
*收稿日期:2015-11-13接受日期:2016-02-24
基金项目:林业公益性行业科研专项课题(201404202);黑龙江省科研院所基本科研业务费专项资金
通信作者:周蕴薇(1970-),女,吉林长春人,教授,博士,主要从事园林植物资源等方向的研究工作。E-mail:nefuzhouyw@126.com
中图分类号:Q945.78
文献标志码:A
文章编号:1001-0629(2016)7-1342-11*
Corresponding author:Zhou Yun-weiE-mail:nefuzhouyw@126.com
Research on the physiological characteristics ofMiscanthussinensisseedlings under the treatments of complex saline-alkaloid stress
He Miao1,Wang Huan1, Xu Peng-fei2, Liu Chang-le3, Zhou Yun-wei1
(1.Landscape Architecture College of Northeast Forestry University, Harbin 150040, China;2.Weihai Lvyuan Gardening Engineer Co., Ltd, Weihai 264200, China;3.Heilongjiang Academy of Forestry, Harbin 150040, China)
Abstract:Soil salinization is prevalent in the world. The increasingly salinization seriously threatens the growth and development of plants, which leads to the reduction of plant species. Miscanthus sinensis, which possesses good economic efficiency and ecological benefits, was a sort of emerging energy plant. In this study, M. sinensis introduced from A Jialing of Benxi City in Liaoning Province were used as materials. Through simulating the saline-alkali soil environment of Daqing in northeast of China which include low(4.147 5, 8.295 0 g·L-1), middle(12.442 5 g·L-1) and high concentration(16.590 0, 20.737 5 g·L-1), we studied each physiological index of the seedlings when they were in the complex saline-alkaloid stress treatment. The study found:When M. sinensis seedlings were in complex saline-alkaloid stress treatment, each index of CK group had a little change over time; the chlorophyll content in low concentration treatment showed a trend of rising first then falling; while in middle and high concentration treatment, the chlorophyll content dropped; the relative electrical conductivity and MDA content increasedover time; and the SOD, POD, CAT, APX, GR activity, ASA, GSH content and root activity were increased in low concentration, but in the treatment of high concentration, these indexes showed a trend of rising first then falling. This study basically defined the range of the saline-alkaloid tolerance of M. sinensis which can provide a theoretical basis for the salt-tolerant screening and applications in landscape of energy plants like M. sinensis in the future.
Key words:Miscanthus sinensis; seedling; complex saline-alkaloid stress; physiological characteristics
何淼,王欢,徐鹏飞,刘长乐,周蕴薇.模拟复合盐碱胁迫对芒幼苗生理特性的影响.草业科学,2016,33(7):1342-1352.
He M,Wang H,Xu P F,Liu C L,Zhou Y W.Research on the physiological characteristics ofMiscanthussinensisseedlings under the treatments of complex saline-alkaloid stress.Pratacultural Science,2016,33(7):1342-1352.
第一作者:何淼(1975-),女(满族),辽宁本溪人,副教授,博士,主要从事园林植物种质资源方面的教学科研工作。E-mail:hemiao@nefu.edu.cn
