不同生境下柠条种子出苗及幼苗生长特征
2016-08-02孙毅闫兴富
孙毅,闫兴富
(国家民委生态系统模型及应用重点实验室, 北方民族大学生物科学与工程学院, 宁夏 银川 750021)
不同生境下柠条种子出苗及幼苗生长特征
孙毅,闫兴富*
(国家民委生态系统模型及应用重点实验室, 北方民族大学生物科学与工程学院, 宁夏 银川 750021)
摘要:为了揭示柠条种群实生更新的机制及其影响因素,在宁夏白芨滩自然保护区的人工柠条灌丛群落,设置不同的灌丛(柠条灌丛、沙蒿灌丛和裸露沙地)基部生境、不同坡位(底部、中部和顶部)的沙丘生境和柠条灌丛植株基部不同距离(0.5,1.5 m和丛间沙地)生境,研究了柠条种子在不同生境下的出苗和幼苗生长特征。结果表明,1) 不同生境(柠条灌丛、沙蒿灌丛和裸露沙地)和沙埋处理对柠条种子出苗率和留存率均具有显著影响(P<0.05);在沙蒿灌丛,沙埋种子的早期出苗率显著高于地表种子(P<0.05),且显著高于裸露沙地和柠条灌丛的沙埋种子(P<0.05);种子的出苗率在沙蒿灌丛的沙埋处理最高,但不同生境间、沙埋和非沙埋种子间的差异均未达到显著水平。2) 沙丘底部的种子出苗率显著大于沙丘顶部(P<0.01),试验结束时约为沙丘顶部的20倍;各时期出苗率均在距柠条灌丛基部1.5 m处最大,灌丛间沙地最小,沙丘不同部位生境的出苗率均在7月18日最高,整个试验期间的出苗率和出苗速率在不同生境间均无显著差异。3) 幼苗株高、基径、主根长、根茎比和总干质量均在沙丘底部最大,而根冠比在沙丘顶部最大;距灌丛基部0.5 m处幼苗的株高显著大于1.5 m处幼苗(P<0.05),幼苗主根长、根茎比和根冠比均随距灌丛距离的增大而增大,而总干质量随灌丛距离的增大而减小。
关键词:柠条;种子萌发;出苗;生长;实生更新
木本植物的实生苗更新不仅对植物种群的数量动态和分布格局具有重要影响,而且在植物群落的演替、退化植被恢复及生物多样性维持等方面都具有重要作用。种子是植物生活史的重要阶段,也是植物在时空上逃避不利的环境并确保成功定居和更新的保障,种子萌发和幼苗建立是种群更新过程的重要环节。植物种子的萌发特性与其自然分布区的气候和生境条件密切相关,是植物对分布区自然生境长期适应的结果。在恶劣的环境条件下,种子长时间保持休眠状态或“坐待”适宜萌发的条件是一种有效分散萌发风险的适应性策略[1-2],而且可降低种子萌发和幼苗建立过程中的死亡风险[3-4];相反,种子响应单一环境因子的迅速萌发可能导致幼苗的大量死亡[1],因为种子一旦萌发,个体即从一生中风险最小的阶段跨越到风险最大、最为脆弱的阶段[5]。在干旱的荒漠地区,水分是影响幼苗生长与存活的重要因素之一,幼苗建立期间的水分胁迫是限制荒漠地区植物种群实生更新的重要瓶颈因素[6]。因此,种子成熟散落最初形成的种子雨“模板”可能在种子的二次扩散过程中因动物捕食、种子萌发和幼苗生长与存活等而改变[7]。植物幼苗的生长和存活受微环境改变的强烈影响[5],例如,灌丛可因其对水分、养分截留及遮阴等作用而改变周围的局部土壤和植被环境,从而导致树冠下的养分和水分与周围空地相比更为肥沃,即灌丛肥岛效应[8];灌丛“肥岛”不仅具有减弱灌丛下土壤干旱胁迫的保育效应[9],而且可有效促进植物幼苗的建立[10-12]。此外,不同坡位生境的土壤含水量差异也是影响植物生长的重要因素[13-14]。
柠条(Caraganakorshinskii)是豆科(Leguminosae)锦鸡儿属旱生灌木,主要分布于沙地草场、荒漠或半荒漠地区的流动沙地和半固定沙地,是我国西北荒漠地区植树造林和沙漠化防治的主要灌木树种,也是北方农牧交错带极具推广价值和较好固沙效果的重要树种之一。柠条种子于6月中下旬成熟散落,干旱荒漠地区在这一季节的地表温度不会限制种子的萌发和出苗,而且种子萌发对干旱胁迫的较强耐受性[15]决定了这一季节的柠条种子遇有少量降雨能够萌发,但伴随少量降雨后随之而来的干旱胁迫无疑会增大幼苗死亡的风险,而且夏季高温[16]、啮齿动物对种子的取食[17]和过度沙埋[18]等都可能成为限制其实生更新的因素。有关柠条种群实生更新方面的研究主要集中于昆虫对种子的取食危害[19-20]、种子萌发与幼苗建立[15-16,18,21-22]、幼苗根系生长特征[23-24]及种群动态[25]等方面,有关种子萌发和幼苗建立环节的研究主要局限于一些人为控制下的光照、水分和沙埋等环境因素的影响方面,在高度异质性的原生荒漠环境中,种子萌发、出苗和幼苗生长对柠条种群的自然更新可能具有瓶颈性的制约作用,并直接影响干旱荒漠区沙漠化治理和造林的成败,但与此相关的野外观测研究并未引起学术界的足够关注。因此,本研究在宁夏白芨滩自然保护区的人工柠条群落,研究了不同微生境条件下种子的出苗和幼苗生长特征,研究结果可为深入揭示该区人工柠条种群的实生更新机制、促进干旱荒漠地区退化草地的恢复和可持续利用提供科学依据。
1材料与方法
1.1研究区概况
宁夏灵武白芨滩国家级自然保护区地处鄂尔多斯台地西南隅(37°54′-38°22′ N;106°23′-106°48′ E),海拔1150~1650 m,保护区地貌分为低山丘陵、缓坡丘陵和沙漠低山丘陵等类型;土壤以灰钙土和风沙土为主。该区属典型大陆性季风气候,四季分明,气候干燥,风大沙多,雨量少而集中,年降水量267.3 mm,年蒸发量2634.5 mm,年均温6.7~8.8 ℃,沙面夏季极端温度可高达70 ℃以上;植被以低矮旱生、超旱生灌木和半灌木为主,除以猫头刺(Oxytropisaciphylla)-沙冬青(Ammopiptanthusmongolicus)为主的荒漠灌丛和草原群落以及以柠条为主的荒漠灌丛[26]外,还有大量的人工固沙柠条林、柠条-沙拐枣(Calligonummongolicum)混交林、柠条-细枝岩黄芪(Hedysarumscoparium)混交林和由刺槐(Robiniapseudoacacia)、樟子松(Pinussylvestrisvar.mongolica)、侧柏(Platycladusorientalis)、紫穗槐(Amorphafruticosa)等组成的人工防风林带。
野外样地位于保护区南部的龙坑林区(37°52.5′ N;106°23′-106°30.7′ E,海拔约1350 m)人工固沙柠条群落,灌丛树种除柠条外,还零星分布有少量的沙冬青、沙拐枣;草本植物以沙蒿(Artemisiadesertorum)、黑沙蒿(Artemisiaordosica)、沙蓬(Agriophyllumsquarrosum)、蒺藜(Tribulusterrestris)、达乌里胡枝子(Lespedezadavurica)和角蒿(Incarvilleasinensis)为主,还有少量甘草(Glycyrrhizauralensis)、白香草木樨(Melilotusalbus)、砂蓝刺头(Echinopsgmelini)、鹅绒藤(Cynanchumchinense)、沙葱(Alliummongolicum)、沙芦草(Agropyronmongolicum)、沙生针茅(Stipaglareosa)等。据灵武市气象局(距试验地区约30 km)的观测记录,2013年该区在6月份最大降水量达46.4 mm,其次为5月份(32.2 mm),试验开始的7月份也有较高的降水量;6-8月份的均温都在22 ℃以上(图1)。
1.2不同生境下柠条种子的出苗率

图1 研究地区2013年4-10月份月降水量和月均温Fig.1 Monthly rainfall and average temperature of study area during the period from April to October, 2013
2013年7月15日,在上述人工柠条群落样地,选取柠条灌丛(冠幅直径≥2 m)和沙蒿灌丛(冠幅直径≥0.8 m)植株各4株(丛),另在灌丛间选取面积4 m2的裸露沙地4处,所选植株和裸露沙地间相互间隔20 m以上。在所选的灌丛植株下和裸露沙地将种子分别作以下处理:1)置于沙地表面;2)以厚约1 cm湿沙埋藏。试验用种子于2012年6月中旬采自试验地区的10年以上树龄的成年柠条灌丛植株,种子采集后,置于室温条件下自然风干后贮藏于阴凉室温下的透气布袋内;取大小均匀、无虫蛀、发育良好的种子,在上述每一处理分别投放种子100粒,总共用种子:4重复×3生境(柠条灌丛、沙蒿灌丛和裸露沙地)×2处理×100粒=2400粒。分别于7月31日和8月16日统计完好种子(种子保持未吸水状态)、霉烂种子(明显被微生物侵染)和萌发出苗种子(种子萌发且至少有1片真叶)的数量,分别计算不同类型种子的百分比;其中8月16日只统计出苗数和留存种子总数(包括部分已萌发但未出苗的种子),计算出苗率和种子留存率。
1.3不同坡位对种子出苗和幼苗生长的影响
于2013年6月22日,在上述人工柠条群落内选取4个较大的半月形沙丘,分别在每一沙丘背风一侧(东南方向)的底部(bottom of sand dune, BSD)、中部(middle of sand dune, MSD)和顶部(top of sand dune, TSD) 3个不同坡位各设置3个1 m×1 m的小样方,共设置小样方4重复×3坡位×3个=36个。每一小样方内播入预先分拣过的无虫蛀柠条种子(来源和贮藏方式同上)100粒;播种时,先将样方内的上层约20 cm深的表层沙土翻开,浇水后将种子点播于湿润沙土表面,再用约1 cm厚的湿沙将种子覆盖;为了防止强光下过度的水分蒸发,播种后立即在样方的4个角以4根长约50 cm的木棍作为支撑,用2层遮阳网在距地面高约20 cm处将每一小样方遮住(遮光率约70%),除在背风一侧预留通风口外,其他部位的边缘均埋入地下以防强风引起水分过度蒸散,播种2周后撤去遮阳网。分别于2013年7月7日、18日、23日和31日,以幼苗出现第一片真叶为标准,记录种子出苗数,此后约每隔2周统计记录一次幼苗留存数;因种子出苗周期较长,且出苗过程中幼苗大量死亡,评价种子出苗质量的指标用出苗率(percentage of seedling emergence, PSE);出苗速率(emergence rate, ER)按照Rozema[27]的公式计算,出苗速率越大,表示出苗率越快。2013年10月6日,收获全部存活的幼苗,收获时小心地将幼苗连根挖出,带回实验室用清水洗净,用滤纸吸干表面水珠后测定幼苗的株高(seedling height, SH)、基径(basal stem diameter, BSD) (用数字式游标卡尺测定)、主根长(taproot length, TRL);测定后分根、茎(含叶轴)和叶片在85 ℃烘箱中烘干48 h后用1/10000电子天平分别称重;根据以上数据计算幼苗根茎比(root stem ratio, RSR)、总干质量(total dry mass, TDM)和根冠比(root shoot ratio, RSHR)。计算公式如下:
PSE=∑(Gi×100%)/n
ER=∑(100×Gi/n×ti)
式中,n为每一试验处理使用的种子总数;Gi为ti(ti=1, 2, 3, ……)天的出苗数。
RSR=主根长/茎长(株高)
RSHR=根干质量/茎叶干质量
1.4灌丛沙堆和灌丛间沙地对种子出苗和幼苗生长的影响
于2013年6月22日,在上述人工柠条群落选取冠幅直径约2.5 m的柠条灌丛植株3株(丛),各植株间隔20 m以上;在每一植株周围相互垂直的4个方向距离植株基部0.5和1.5 m处各设置1个1 m×1 m的小样方,另在距每一植株约10 m处的丛间沙地分别设置同样大小的3个小样方(间隔2 m)作为对照,共设置小样方33个。播种方法、数据记录频次和幼苗收获时间、测定方法及相关参数的计算与“1.3”相同。
1.5数据处理
在进行统计分析前将全部试验数据进行平方根转换,用二因素方差分析的方法分析“1.2”中生境和处理对种子出苗率和其他类型种子百分比的影响;采用单因素方差分析的最小显著差法(least significant difference, LSD)分析其他研究数据在不同处理或水平间的差异;所有数据的统计分析均在SPSS 13.0软件中进行。用Excel 2003和SigmaPlot 10.0制作图表,全部数据以平均值±标准差表示。
2结果与分析
2.1柠条种子在不同灌丛生境的出苗率
从表1可以看出,柠条灌丛、沙蒿灌丛和裸露沙地3种生境对7月31日的柠条出苗率和霉烂率具有显著影响,但对种子完好率的影响不显著,对出苗率和留存率均影响显著;沙埋与否对种子出苗率和完好率影响显著,但对种子霉烂率影响不显著,不同处理对8月16日的种子最终出苗率无显著影响,但显著影响种子的最终留存率;生境和处理间的交互作用对不同类型种子命运的影响均未达显著水平。

表1 生境和处理对宁夏白芨滩自然保护区人工林下柠条种子出苗率影响的二因素方差分析Table 1 Two way ANOVA analysis of effects of habitat and treatment of sand burial on emergence percentage ofC. korshinskii seedlings in forest plantation of C. korshinskii in Baijitan Nature Reserve of Ningxia, China
PSE: percentage of seedling emergence; PIS: percentage of intact seed; PRO: percentage of rotten seed; PRE: percentage of remained seed; 下同 The same below.
从表2可以看出,地表种子的留存率在沙蒿灌丛最高(18.0%),裸露沙地次之(3.5%),前者显著高于后者和柠条灌丛生境(P<0.05),其他类型的地表种子的百分比在生境间差异不显著。在7月31日,沙蒿灌丛下沙埋种子的出苗率(21.25%)显著高于柠条灌丛和裸露沙地生境(P<0.05),沙埋种子的霉烂率在沙蒿灌丛显著高于柠条灌丛(P<0.05),裸露沙地上沙埋种子的霉烂率最低(0.50%),但与柠条和沙蒿灌丛间均差异不显著;沙埋种子的留存率在沙蒿灌丛显著高于柠条灌丛(P<0.05),尽管沙埋种子留存率在裸露沙地最高,但其与柠条灌丛、沙蒿灌丛间无显著差异。在沙蒿灌丛生境,7月31日的沙埋种子的出苗率显著高于地表种子(P<0.05),但其完好率显著低于地表种子(P<0.05),试验结束时地表种子的留存率显著高于沙埋藏种子(P<0.05);在柠条灌丛和裸露沙地生境,各类型种子的百分比在地表种子和沙埋种子间均差异不显著。

表2 宁夏白芨滩自然保护区人工柠条群落不同生境下柠条种子的出苗率Table 2 The seedling emergence percentage of C. korshinskii seeds in different habitats of forest plantation ofC. korshinskii in Baijitan Nature Reserve in Ningxia, China %
SS和SB分别为地表和沙埋。 SS and SB is sand surface and sand burial respectively; 同一参数在相同处理的不同生境间的不同大写字母和同一生境内不同处理间的不同小写字母表示差异显著(P<0.05)。 Different capital letters with the same parameters in different habitat within the same treatment and small letters in different treatment within the same habitat mean significant differences at 0.05 level.
2.2不同坡位和灌丛生境柠条种子的出苗率
2.2.1不同坡位的沙丘生境从图2可以看出,在沙丘底部,各观测时期的出苗率均高于沙丘中部和顶部,沙丘中部的出苗率除7月23日外的其他时期均高于沙丘顶部,其中沙丘底部和沙丘顶部间的差异在多数观察时期达显著水平(P<0.05),试验结束时沙丘底部的出苗率约为顶部的20倍;在出苗早期阶段,沙丘不同坡位生境的出苗率均略有提高,但从7月23日开始,出苗率持续降低,表明死亡幼苗的逐渐积累(图2A)。沙丘不同坡位生境的出苗速率与出苗率一致,沙丘底部、沙丘中部和沙丘顶部的出苗速率分别为9.8,7.7和6.3,其中在沙丘底部与沙丘顶部间差异显著(P<0.05) (图2B)。
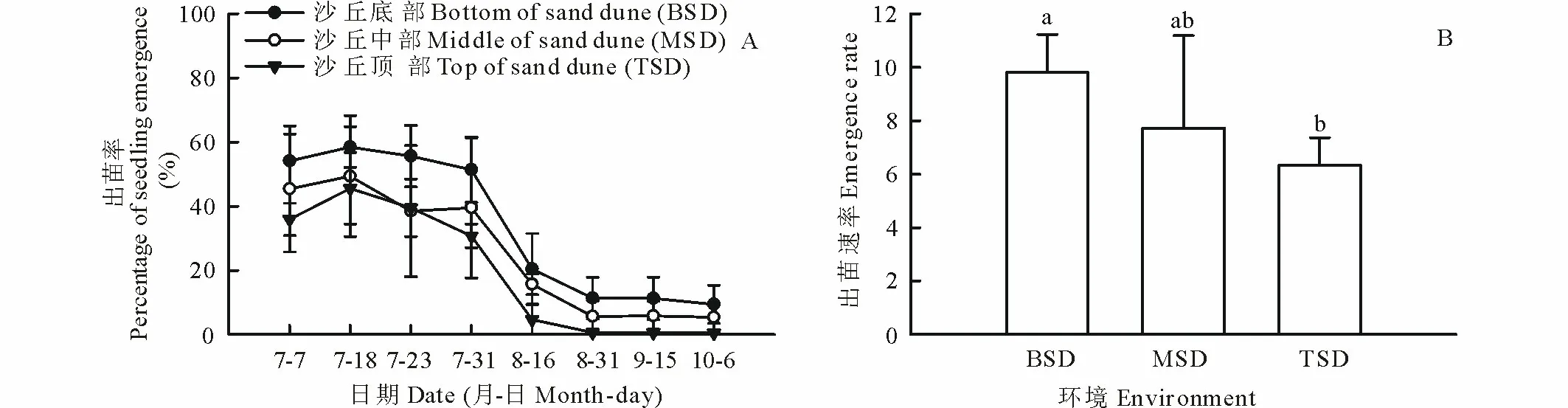
图2 宁夏白芨滩自然保护区人工柠条群落不同坡位的沙丘生境中柠条种子的出苗率动态和出苗速率Fig.2 Dynamics of seedling emergence percentage and emergence rate of C. korshinskii seeds at different slope position of sand dune in forest plantation of C. korshinskii in Baijitan Nature Reserve of Ningxia, China 不同小写字母表示不同坡位生境间差异显著(P<0.05)。Different small letters means significant differences among different slope habitats of sand dune at 0.05 level; 下同 The same below.
2.2.2柠条灌丛基部和灌丛间沙地距柠条灌丛基部约1.5 m处的出苗率在各个时期均最大,灌丛间沙地最小,但各个观察时期的出苗率,在灌丛基部不同距离间均无显著差异。3种不同生境的出苗率均在7月18日达到最大值,距灌丛基部约0.5,1.5 m处和灌丛间沙地分别为39.4%,49.2%和38.3%,此后均随时间的延长而持续下降(图3A)。出苗速率在距灌丛基部0.5,1.5 m和灌丛间沙地生境分别为6.0,7.0和5.3,各生境间差异不显著(图3B)。
2.3不同坡位和灌丛生境柠条幼苗的生长特征
2.3.1不同坡位的沙丘生境柠条幼苗的株高、基径、主根长、根茎比均在沙丘底部最大,分别为4.8 cm,1.0 mm,23.2 cm和6.1,除株高外的其他3个生长参数在沙丘底部均显著大于沙丘顶部(P<0.05),主根长和根茎比在沙丘中部与沙丘顶部间也差异显著(P<0.05);幼苗总干质量也在沙丘底部最大,不同坡位生境间差异不显著;根冠比在沙丘底部最小,沙丘顶部最大,生境间无显著差异(图4)。
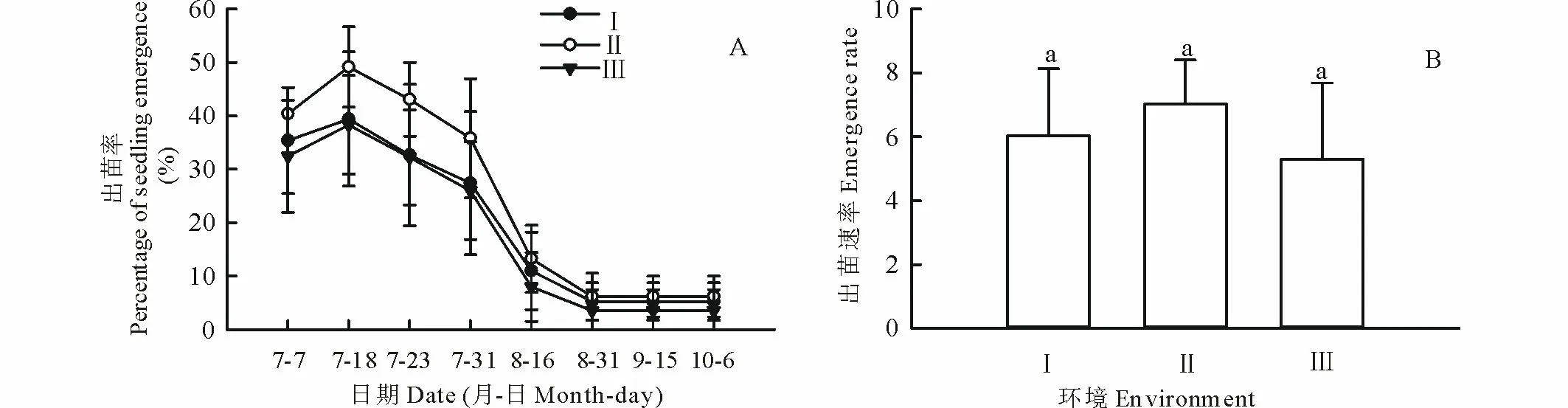
图3 宁夏白芨滩自然保护区人工柠条群落灌丛基部和灌丛间沙地生境下柠条种子的出苗率动态和出苗速率Fig.3 Dynamics of seedling emergence percentage and emergence rate of C. korshinskii seeds at shrub base and sand land of shrub-interspaces in forest plantation of C. korshinskii in Baijitan Nature Reserve of Ningxia, China Ⅰ、Ⅱ和Ⅲ分别为距灌丛基部0.5 m、距灌丛基部1.5 m和灌丛间沙地 Ⅰ, Ⅱ and Ⅲ is 0.5, 1.5 m from shrub base and sand of shrub-interspace respectively; 不同小写字母表示距离灌丛不同距离生境间差异显著(P<0.05)。Different small letters mean significant differences among habitats of different distance from shrubs base at 0.05 level; 下同 The same below.
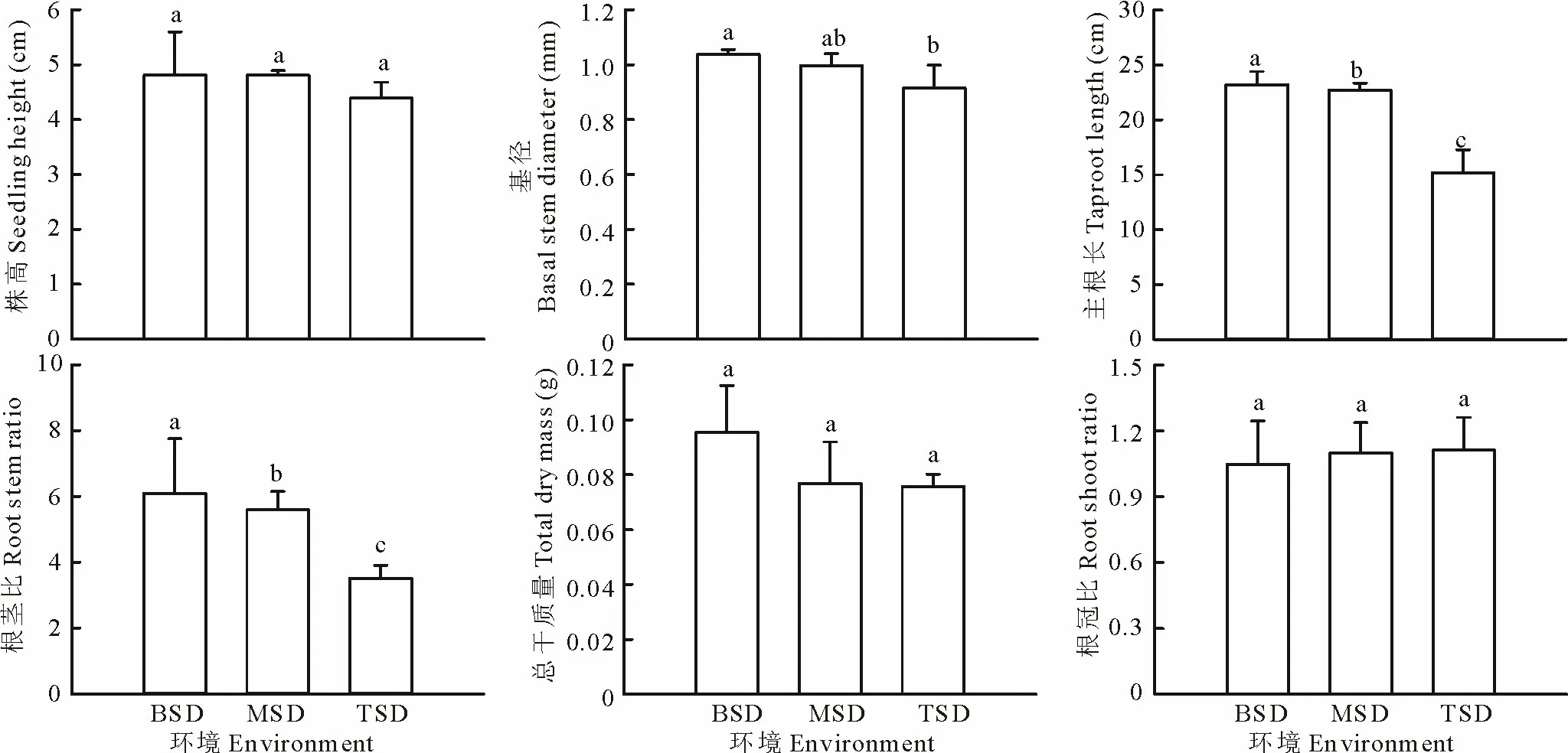
图4 宁夏白芨滩自然保护区人工柠条群落不同坡位沙丘生境对柠条幼苗生长的影响Fig.4 Effects of different slop position of sand dun habitats on the growth of C. korshinskii seedlings in forest plantation of C. korshinskii in Baijitan Nature Reserve of Ningxia, China
2.3.2灌丛基部和灌丛间沙地从图5可以看出,幼苗株高在距灌丛基部0.5 m处最大为6.6 cm,显著大于距灌丛基部1.5 m处幼苗(P<0.05);基茎在距灌丛基部不同生境间无显著差异;主根长和根茎比均随着距灌丛基部距离的增大而增大,在灌丛间沙地达最大值,分别为23.1 cm和5.6,且均显著大于距灌丛基部0.5 m幼苗的最小值(P<0.05);幼苗总干质量在距灌丛基部0.5 m最大(0.1 g),随着距灌丛距离的增大而减小,生境间差异不显著;幼苗根冠比在距灌丛基部0.5和1.5 m间差异显著(P<0.05),但两者与灌丛间沙地间无显著差异。

图5 宁夏白芨滩自然保护区人工柠条群落灌丛基部和丛间沙地生境对柠条幼苗生长的影响Fig.5 Effects of shrub base and sand land of shrub-interspaces on the growth of C. korshinskii seedlings in forest plantation of C. korshinskii in Baijitan Nature Reserve of Ningxia, China
3讨论
柠条种子于6月中下旬成熟,这一时期散落于地表的种子如遇降雨即可快速萌发,但在降雨量和降雨时间高度不确定的荒漠环境中,种子萌发形成的幼苗可能因少量降雨后随之而来的干旱胁迫而大量死亡,少量未能萌发的种子进入土壤种子库可降低干旱造成的死亡风险。本研究发现,柠条灌丛、沙蒿灌丛和裸露沙地等生境条件均显著影响柠条种子的出苗率和留存率,置于地表的种子留存率在沙蒿灌丛最高,柠条灌丛最低,表明荒漠环境中偶然出现的少量降水可通过植株冠层下的沙地孔隙迅速渗入深层土壤而促进种子萌发和出苗,即灌丛根际土壤具有富集降水的“水分岛”效应[6]。研究结果还发现,干旱地区植物的种子可对少量降水做出快速反应而萌发并建立幼苗,尤其在沙埋条件下对促进柠条幼苗建立的效果更好,但根据赵晓英和任继周[17]对3种锦鸡儿属植物的研究结果,沙埋对出苗的促进作用也非常有限,埋藏的种子很快被啮齿动物挖掘取食或出苗后被取食。此外,尽管种子的出苗率在不同生境间差异不显著,但沙蒿灌丛仍有微弱优势,可能与其浓密枝叶的遮阴降低了土壤水分的散失有关。本研究中除出苗、留存(完好)和霉烂种子外,部分未能统计到的“丢失”种子可能被啮齿动物取食或贮藏于洞穴和其他生境中;根据调查过程中在投放点遗留的种皮残渣和动物粪便判断,啮齿动物可能是这些“丢失”种子的捕食者,但调查中并未发现任何种子被贮藏的证据。已有报道显示,荒漠草原中的沙鼠(Merionessp.)和跳鼠(Dipussp.)类啮齿动物均有捕食植物种子的记录[28],而且有研究结果表明锦鸡儿属植物的种子在成熟散落后大量被啮齿动物取食[17]。灌丛生境可能至少在种子投放的早期具有使种子免遭啮齿动物取食的保护作用,尽管啮齿动物在裸露沙地被捕食的风险更高,但这一生境为靠视觉觅食的啮齿动物提供了便利,因而在早期调查结果中种子完好率较高。另一方面,与沙蒿灌丛相比,柠条灌丛基部分布有更多的鼠类洞穴,是啮齿动物更为偏好的活动场所,因而种子留存率在柠条灌丛下最低;相反,低矮而浓密的沙蒿灌丛对啮齿动物的活动具有机械障碍作用,而且蒿属植物特有的气味也可能对动物有驱避作用,尤其在长期缺乏降雨的干燥条件下气味更为浓烈,这些因素都可能在一定程度上减弱啮齿动物对柠条种子的捕食强度,但并未见相关的研究报道,有待于更加深入的研究。
植物种子的萌发特性与其自然分布区的气候和生境条件密切相关,在长期的适应过程中形成了两种对立的萌发策略:一是遇有适宜萌发的条件即迅速萌发建立幼苗以避开不利于生长与存活的恶劣环境,二是在适宜萌发的条件下缓慢萌发或保持休眠并进入土壤种子库的“坐待”策略;显然,前者属于“冒险”的萌发策略,因为种子一旦萌发,个体便从一生中风险最小的阶段跨越到风险最大和最为脆弱的阶段[5],而后者则可分散种子萌发和幼苗建立过程中的死亡风险[1-2]。本研究发现,沙丘底部种子的出苗率均高于沙丘中部和顶部,甚至在试验结束时的出苗率在沙丘底部约为沙丘顶部的20倍,且出苗速率表现为沙丘底部>沙丘中部>沙丘顶部。这些结果表明,种子成熟散落后萌发期间的土壤水分状况是影响柠条出苗的重要环境因素,尤其是出苗从7月份以后,不论是不同坡位的沙丘生境,还是柠条灌丛不同距离的生境,出苗率都持续降低,表明8月份前后降水量的急剧减少(8月份降水量仅有6.1 mm)导致了死亡幼苗的逐渐积累。在沙丘底部,相对充足的水分可在一定程度上满足种子萌发的水分需求,因而种子出苗的机会相对较高;相反,在容易遭受干旱胁迫的沙丘顶部,那些经历反复吸水和脱水过程而未能萌发的种子则更易于保持休眠[29],因而有更多的机会进入土壤种子库而得以幸存。干旱荒漠地区灌丛植株的“肥岛”效应作为幼苗建立的重要机制[10-12],对柠条种子的萌发和出苗具有促进作用,而导致柠条灌丛沙堆内不同部位的种子出苗率和出苗速率差异的原因可能与沙堆内部光照和水分资源的异质性有关;尽管光照不是其种子萌发的限制因素[30],但在距灌丛基部更近的0.5 m处的种子也可能会由于灌丛遮阴和播种早期人为遮阴造成的过度荫蔽而影响出苗,而且还可能影响水分蒸散导致土壤水分过多而降低种子萌发和出苗[21]。因此,本研究中柠条种子的出苗率和出苗速率均在距灌丛基部较远的1.5 m处更大。从本研究的沙丘顶部和灌丛间沙地的种子出苗率和出苗速率判断,水分是柠条种子萌发和出苗的限制因素。尽管大量研究强调柠条种子萌发对干旱胁迫具有较强的耐性[15,18],但在干旱的荒漠环境中,种子长时间保持不萌发状态可能是其防止在适宜萌发但不利于幼苗建立的条件下萌发的适应性策略[2],这一策略可降低大量种子同时萌发后因缺乏连续降水而导致的幼苗死亡[1,3],并可在一定程度上分散萌发风险[1,4]。
幼苗建立和生长期间的水资源亏缺是限制荒漠地区植物种群实生苗更新的重要瓶颈因素[6]。有研究结果显示,在内蒙古典型草原,0~60 cm土层中的土壤含水量从坡顶到坡底呈逐渐增加的趋势[14]。柠条具有较强的耐受干旱胁迫的能力[12],其幼苗可充分利用沙丘底部相对充足的土壤水分快速生长,因而本研究中柠条幼苗的株高、基径、主根长、根茎比和幼苗总干质量等生长参数均在沙丘底部最大,尤其是主根的快速伸长和大量侧根的形成可为幼苗吸收利用深层土壤的水分和养分提供保障[31];根冠比从坡底到坡顶的逐渐增大反映了幼苗应对干旱胁迫的适应[32-33],根冠比的增大不仅意味着有相对更多的根系用于水分和养分的吸收[34],而且地上生物量分配的减少可降低生长于极端干旱的坡顶生境中幼苗的水分消耗,高慧等[13]以草本植物大针茅(Stipagrandis)为材料也发现了类似的研究结果。研究发现,柠条幼苗的株高、基径和总干质量均在距灌丛基部0.5 m处最大,这一结果可能是幼苗生长对灌丛基部不同距离微生境土壤水分和养分分布异质性反应的结果,因为灌丛对其自身树冠下水分和养分条件的改善与植物的生长密切相关[8];大量研究结果表明,灌丛对水分和养分富集的“肥岛”效应对植物地上部分生长具有重要促进作用[11-13]。幼苗主根长、根茎比和根冠比都在灌丛间沙地最大,表明幼苗可通过形态学适应[32,35]以应对干旱胁迫,从而使分配至根系的生物量随着灌丛基部距离的增大而增加,即幼苗以牺牲地上部分的生长为代价增强吸收深层土壤水分的能力,以确保幼苗在低土壤含水量条件下维持生长[33]。
References:
[1]Bu H Y, Ren Q J, Xu X L,etal. Seed germination characteristics of 54 gramineous species in the alpine meadow on the Eastern Qinghai-Tibetan Plateau. Chinese Journal of Plant Ecology, 2006, 30(4): 624-632.
[2]Wang J H, Ma R J, Chen W. Effects of cold stratification and dry storage at room temperature on seed germination of eight desert species from the Hexi Corridor of China. Chinese Journal of Plant Ecology, 2012, 36(8): 791-801.
[3]Liu Y J, Liu S Z, Ji Y F,etal. Seed germinative strategy and ecological adaptability ofCorispermumpatelliforme. Acta Ecologica Sinica, 2010, 30(24): 6910-6918.
[4]Zhang L, Zhang C H, Lv J P,etal. Seed germination characteristics and their correlations with seed sizes of 31 common weeds in eastern Qinghai-Tibet Plateau. Chinese Journal of Ecology, 2011, 30(10): 2115-2121.
[5]Su Y, Jiao J Y, Wang Z J. Characteristics of seedling survival in habitats of hill and gully slopes in hill-gully Loess Plateau region of northern Shaanxi. Chinese Journal of Plant Ecology, 2014, 38(7): 694-709.
[6]Liu H L, Shi X, Wang J C,etal. Effects of sand burial, soil water content and distribution pattern of seeds in sand on seed germination and seedling survival ofEremospartonsongoricum(Fabaceae), a rare species inhabiting the moving sand dunes of the Gurbantunggut Desert of China. Plant and Soil, 2011, 345(1): 69-87.
[7]Du Y J, Ma K P. Temporal and spatial variation of seed fall in a broad-leaved evergreen forest in Gutianshan Nature Reserve of Zhejiang Province, China. Chinese Journal of Plant Ecology, 2012, 36(8): 717-728.
[8]Qu W L, Yang X P, Zhang C T,etal. Shrub mediated “fertile island” effects in arid and semi-arid grassland. Acta Prataculturae Sinica, 2015, 24(4): 201-207.
[9]Rango A, Tartowshi S L, Laliberte A,etal. Islands of hydrologically enhanced biotic productivity in natural and managed arid ecosystems. Journal of Arid Environments, 2006, 65(2): 235-252.
[10]Zhao H L, Zhou R L, Su Y Z. Shrub facilitation of desert land restoration in the Horqin Sand Land of Inner Mongolia. Ecological Engineering, 2007, 31(1): 1-8.
[11]Zhang P J, Yang J, Song B Y,etal. Spatial heterogeneity of soil resources ofCaraganatibeticacommunity. Chinese Journal of Plant Ecology, 2009, 33(2): 338-346.
[12]Zhang P J, Yang J, Zhao L Q. Ecological adaptability ofCaraganatibeticaindividuals in the Ordos plateau. Arid Zone Research, 2011, 28(5): 839-848.
[13]Gao H, Gao Y B, Liu H Y,etal. Characteristics ofStipagrandisgrowth and reproductive biomass allocation at different hill position of Xilingol grassland, Inner Mongolia. Chinese Journal of Applied Ecology, 2009, 20(9): 2123-2128.
[14]Peng H Y, Li X Y, Tong S Y. Effects of shrub (CaraganamicrophallaLam.) encroachment on water redistribution and utilization in the typical steppe of Inner Mongolia. Acta Ecologica Sinica, 2014, 34(9): 2256-2265.
[15]Zheng M Q, Zheng Y R, Jiang L H. Effects of one-time water supply and sand burial on seed germination and seedling emergence of four popular psammophyte in Mu Us sandy land. Acta Ecologica Sinica, 2006, 26(8): 2474-2484.
[16]Yan X F, Zhou L B, Si B B,etal. Stress effects of simulated drought by polyethylene glycol on the germination ofCaraganakorshinskiiKom. seeds under different temperature conditions. Acta Ecologica Sinica, 2016, 36(7): 1989-1996.
[17]Zhao X Y, Ren J Z. Influence of seed predation on regeneration of threeCaranagaspecies. Biodiversity Science, 2005, 13(6): 514-519.
[18]Yang H L, Liang Z L, Zhu X W,etal. Effects of sand burial and seed size on seed germination, seedling emergence and growth ofCaraganakorshinskiiKom. (Fabaceae). Acta Ecologica Sinica, 2012, 32(24): 7757-7763.
[19]Luo Y Y, Li Q F, Jin H. Survey of seed pest harm onCaraganaand its influence on seed quality in Inner Mongolia. Research of Soil and Water Conservation, 2006, 13(4): 25-29.
[20]Zhang D Z, He D H. Responses ofCaraganaseed pests to host plant patch quality and patch pattern in desert regions of Ningxia, Northwest China. Chinese Journal of Applied Ecology, 2011, 22(7): 1871-1877.
[21]Nie C L, Zheng Y R. Effects of water supply and sand burial on seed germination and seedling emergence of four dominant psammophytes in the Ordos plateau. Chinese Journal of Plant Ecology, 2005, 29(1): 32-41.
[22]Yan X F, Liu J L, Bei Z L,etal. Characteristics of seed germination and seedling growth ofCaraganakorshinskiiunder different light intensities. Chinese Journal of Ecology, 2015, 34(4): 912-918.
[23]Wang M B, Xun J J, Chen J W,etal. The net growth rate of fine roots ofCaraganakorshinskiiseedlings in the Loess Plateau region, Northwest Shanxi. Acta Ecologica Sinica, 2010, 30(5): 1117-1124.
[24]Zhang F, Chen J W, Wang M B. The spatial distribution and seasonal dynamics of fine roots in a youngCaraganakorshinskiiplantation. Acta Ecologica Sinica, 2012, 32(17): 5484-5493.
[25]Bao C H, Zhang W H, He J F. Dynamics of artificialCaraganakorshinskiipopulation planted for thirty years in hilly region of Loess plateau. Acta Botanica Boreali-Occidentalia Sinica, 2010, 30(8): 1681-1688.
[26]Song C S, Wang Y D. Scientific Survey of Ningxia Baijitan Naturae[M]. Beijing: Chinese Forestry Press, 1999.
[27]Rozema J. The influence of salinity, inundation and temperature on germination of some halophytes and non-halophytes. Oecologia Plantarum, 1975, 10: 341-353.
[28]Liu C Z. Protection of Grassland[M]. Beijing: China Agricultural University Press, 2009: 368-397.
[29]Zhu Y J, Yang X J, Baskin C C,etal. Effects of amount and frequency of precipitation and sand on seed germination, seedling emergence and survival of the grassLeymussecalinusin semiarid China. Plant and Soil, 2014, 374(1): 399-409.
[30]Zheng Y R, Xie Z X, Gao Y,etal. Germination responses ofCaraganakorshinskiiKom. to light, temperature and water stress. Ecological Research, 2004, 19(5): 553-558.
[31]Zhu Y L, Wang S, Lin Y G,etal. Development ofCaraganamicrophyllaseedling root system in hilly regions of Loess plateau. Bulletin of Soil and Water Conservation, 2011, 31(2): 232-237.
[32]Zhang M, Zhu J J, Li M C,etal. Different light acclimation strategies of two coexisting tree species seedlings in a temperature secondary forest along five natural light levels. Forest Ecology and Management, 2013, 306(15): 234-242.
[33]Tian Y, Ta Xi P L T, Li Y,etal. The survival and above/below ground growth ofHaloxylonammodendronseedling. Acta Ecologica Sinica, 2014, 34(8): 2012-2019.
[34]Jin J Y, Zhang W H, Yuan L. Physiological responses of three forages to drought stress and evaluation of their drought resistance. Acta Prataculturae Sinica, 2015, 24(10): 157-165.
[35]Hamerlynck E P, Huxman T E. Ecophysiology of two Sonoran Desert evergreen shrubs during extreme drought. Journal of Arid Environments, 2009, 73(4-5): 582-585.
参考文献:
[1]卜海燕, 任青吉, 徐秀丽, 等. 青藏高原东部高寒草甸54种禾本科植物种子的萌发特性. 植物生态学报, 2006, 30(4): 624-632.
[2]王桔红, 马瑞君, 陈文. 湿冷层积和室温干燥贮藏对河西走廊8种荒漠植物种子休眠水平的影响. 植物生态学报, 2012, 36(8): 791-801.
[3]刘有军, 刘世增, 纪永福, 等. 碟果虫实种子萌发对策及生态适应性. 生态学报, 2010, 30(24): 6910-6918.
[4]张蕾, 张春辉, 吕俊平, 等. 青藏高原东缘 31 种常见杂草种子萌发特性及其与种子大小的关系. 生态学杂志, 2011, 30(10): 2115-2121.
[5]苏嫄, 焦菊英, 王志杰. 陕北黄土丘陵沟壑区坡沟立地环境下幼苗的存活特征. 植物生态学报, 2014, 38(7): 694-709.
[7]杜彦君, 马克平. 浙江古田山自然保护区常绿阔叶林种子雨的时空变异. 植物生态学报, 2012, 36(8): 717-728.
[8]瞿王龙, 杨小鹏, 张存涛, 等. 干旱、半干旱地区天然草原灌木及其肥岛效应研究进展. 草业学报, 2015, 24(4): 201-207.
[11]张璞进, 杨劼, 宋炳煜, 等. 藏锦鸡儿群落土壤资源空间异质性. 植物生态学报, 2009, 33(2): 338-346.
[12]张璞进, 杨劼, 赵利清. 鄂尔多斯高原藏锦鸡儿(Caraganatibetica)个体层次对干旱的生态适应性. 干旱区研究, 2011, 28(5): 839-848.
[13]高慧, 高玉葆, 刘海英, 等. 不同坡位大针茅生长与生殖分配特征. 应用生态学报, 2009, 20(9): 2123-2128.
[14]彭海英, 李小雁, 童绍玉. 内蒙古典型草原小叶锦鸡儿灌丛化对水分再分配和利用的影响. 生态学报, 2014, 34(9): 2256-2265.
[15]郑明清, 郑元润, 姜联合. 毛乌素沙地4种沙生植物种子萌发及出苗对沙埋及单次供水的响应. 生态学报, 2006, 26(8): 2474-2484.
[16]闫兴富, 周立彪, 思彬彬, 等. 不同温度下PEG-6000模拟干旱对柠条锦鸡儿种子萌发的胁迫效应. 生态学报, 2016, 36(7): 1989-1996.
[17]赵晓英, 任继周. 食种子动物对三种锦鸡儿属植物繁殖更新的影响. 生物多样性, 2005, 13(6): 514-519.
[18]杨慧玲, 梁振雷, 朱选伟, 等. 沙埋和种子大小对柠条锦鸡儿种子萌发、出苗和幼苗生长的影响. 生态学报, 2012, 32(24): 7757-7763.
[19]罗于洋, 李青丰, 金花. 内蒙古自治区柠条种子害虫为害情况及其对柠条种子质量的影响. 水土保持研究, 2006, 13(4): 25-29.
[20]张大治, 贺达汉. 荒漠地区柠条种实害虫对寄主斑块质量与格局的反应. 应用生态学报, 2011, 22(7): 1871-1877.
[21]聂春雷, 郑元润. 鄂尔多斯高原4种主要沙生植物种子萌发与出苗对水分和沙埋的响应. 植物生态学报, 2005, 29(1): 32-41.[22]闫兴富, 刘建利, 贝盏临, 等. 不同光强条件下柠条锦鸡儿的种子萌发和幼苗生长特征. 生态学杂志, 2015, 34(4): 912-918.
[23]王孟本, 荀俊杰, 陈建文, 等. 晋西北黄土区幼龄柠条细根的净生长速率. 生态学报, 2010, 30(5): 1117-1124.
[24]张帆, 陈建文, 王孟本. 幼龄柠条细根的空间分布和季节动态. 生态学报, 2012, 32(17): 5484-5493.
[25]保长虎, 张文辉, 何景峰. 黄土高原丘陵沟壑区30年柠条人工种群动态研究. 西北植物学报, 2010, 30(8): 1681-1688.
[26]宋朝枢, 王有德. 宁夏白芨滩自然保护区科学考察集[M]. 北京: 中国林业出版社, 1999.
[28]刘长仲. 草地保护学[M]. 北京: 中国农业大学出版社, 2009: 368-397.
[31]朱元龙, 王桑, 林永刚, 等. 黄土高原丘陵区柠条根系生长发育特性研究. 水土保持通报, 2011, 31(2): 232-237.
[33]田媛, 塔西甫拉提·特依拜, 李彦, 等. 梭梭幼苗的存活与地上地下生长的关系. 生态学报, 2014, 34(8): 2012-2019.
[34]靳军英, 张卫华, 袁玲. 三种牧草对干旱胁迫的生理响应及抗旱性评价. 草业学报, 2015, 24(10): 157-165.
DOI:10.11686/cyxb2015449
*收稿日期:2015-09-23;改回日期:2016-01-26
基金项目:北方民族大学研究生创新研究项目(YCX1529)和国家自然科学基金项目(31260151)资助。
作者简介:孙毅(1991-),男,云南曲靖人,在读硕士。E-mail: 1271169501@qq.com *通信作者Corresponding author. E-mail: xxffyan@126.com
* 1Seed emergence and growth ofCaraganakorshinskiiin different habitats
SUN Yi, YAN Xing-Fu*
KeyLaboratoryofEcosystemModelandApplicationsofStateNationalitiesAffairsCommission,CollegeofBiologicalScienceandEngineering,BeifangUniversityofNationalities,Yinchuan750021,China
Abstract:The aim of this study was to explore the mechanism of seed emergence and the factors affecting seedling regeneration of Caragana korshinskii. Field surveys and experiments were conducted in the C. korshinskii forest plantation in Baijitan Nature Reserve, Ningxia, China. We conducted field surveys and seed burial experiments in three different base habitats: C. korshinskii shrubs (CKS), Artemisia desertorum shrubs (ADS), and bare sand (BS); and in three different sand dune habitats: bottom (BSD), middle (MSD), and top (TSD). We also evaluated seed emergence and seedling growth at different distances from the base of C. korshinskii shrubs (0.5 m, 1.5 m, and shrub-interspace). The results showed that the habitat (CKS, ADS, and BS) and treatment (sand surface and sand burial) had significant effects on the percentage of seedling emergence (PSE) and survival rate of C. korshinskii seeds (P<0.05). In the ADS habitat, for example, the PSE of buried seeds was significantly higher than that of seeds on the sand surface during the early stage of the survey (P<0.05) and significantly higher than those of buried seeds in both the CKS and BS habitats (P<0.05). The highest PSE was in buried seeds in the ADS habitat, but there was no significant difference in PSE between the other two habitats and between buried seeds and those on the sand surface. The results also showed that the PSE of seeds planted at the BSD was significantly higher than that of seeds planted at the TSD (P<0.01; 20 times higher at the end of the experiment). The maximum and minimum PSE was at 1.5 m from the C. korshinskii shrub base and in the shrub-interspace, respectively, on all survey dates. All three habitats showed the highest PSEs on July 18, but there were no significant differences in PSE and emergence rate (ER) among the three habitats during the whole experiment period. The maximum seedling height, basal stem diameter, taproot length, root∶stem ratio, and total dry mass of seedlings were in BSD, while the maximum root∶shoot ratio was in TSD. Seedlings at 0.5 m from the C. korshinskii shrub base had the greatest seedling height (P<0.05). With increasing distance of seedlings from the C. korshinskii shrub base, the tap root length, root∶stem ratio, and maximum root∶shoot ratio increased, but total dry mass decreased.
Key words:Caragana korshinskii; seed germination; seedling emergence; growth; seedling regeneration
http://cyxb.lzu.edu.cn
孙毅, 闫兴富. 不同生境下柠条种子出苗及幼苗生长特征. 草业学报, 2016, 25(7): 186-195.
SUN Yi, YAN Xing-Fu. Seed emergence and growth ofCaraganakorshinskiiin different habitats. Acta Prataculturae Sinica, 2016, 25(7): 186-195.
