北黄花菜的花部特征与繁育系统特性
2016-07-28孙颖杨雪王阿香何淼
孙颖,杨雪,王阿香,何淼
(东北林业大学园林学院,黑龙江 哈尔滨150040)
北黄花菜的花部特征与繁育系统特性
孙颖,杨雪,王阿香,何淼*
(东北林业大学园林学院,黑龙江 哈尔滨150040)
摘 要:以北黄花菜(Hemerocallis lilio-asphodelus)为试材,通过引种观察,运用定量测量、花粉萌发试验、人工授粉试验等对其花部形态特征、开花动态、花粉活力、柱头可授性、繁育系统进行了研究,对其柱头表面及花粉形态进行了扫描电镜观察。结果表明:北黄花菜花型大,芳香,单花花期持续2 d左右,花部具有吸引传粉者的特征;花粉活力在刚开花时较低,12~18 h达到最高,柱头在开花当天即有可授性,花粉活力和柱头可授性均呈先升高后降低的趋势,在持续时间上存在较长相遇期;开花时柱头高于花药,两者存在空间间隔;北黄花菜杂交指数为4,花粉胚珠比为1 838,自然条件下北黄花菜的坐果率达到82.93%,经过人工自花授粉、人工异花授粉后坐果率分别为96.68%和94.32%,人工授同株异花花粉后坐果率为83.45%。综合上述结果,认为北黄花菜的繁育系统为兼性异交,自交可孕,北黄花菜不存在自动的自花授粉及无融合生殖。
关 键 词:北黄花菜;花部特征;繁育系统;杂交指数;花粉胚珠比;花粉活力;柱头可授性
投稿网址:http://xb.ijournal.cn
萱草属(Hemerocallis)是百合科多年生草本植物,约有14种,在东北地区主要分布有5种:萱草(Hemerocallis fulva)、小萱草(Hemerocallis dumortieri)、矮萱草(Hemerocallis nana)、小黄花菜(Hemerocallis minor)
繁育系统是指有性生殖过程中直接影响后代遗传组成的特征和行为系统[3-4],其中包括花部综合特征、花的各器官寿命、花的开放式样、传粉者种类和频率、自交亲和程度和交配系统等,其中交配系统是核心[5-8]。在植物的生活史特征中,繁殖特征决定交配格局,决定基因在种群间的传递,因此,花的形态特征、有性器官的开放时间等因素都会对植物交配系统的发生、发展产生重要影响[9-10]。萱草属植物作为重要的园林植物种类,目前的研究主要集中在植物分类、倍性育种、抗病机理、化学成分、孢粉学等方面[11-17],而有关植物花部综合特征和交配系统的研究鲜见报道。本研究以北黄花菜为对象,对北黄花菜开花过程及其繁育系统类型进行研究,旨在为探明北黄花菜的个体发育特征,为北黄花菜的种质栽培、新品种选育等提供依据。
1 材料与方法
1.1 材料
供试材料为 2011年从黑龙江省大庆市绿色草原牧场引种至东北林业大学园林学院苗圃(44°04′N、125°42′E)的北黄花菜。
1.2 方法
1.2.1 花部综合特征及开花动态观察
于北黄花菜花蕾期,选择生长正常的 10个植株挂牌标记。开花前每天观察植株生长及花蕾变化,直至开花。开花之后,每2 h观察1次,直至花瓣凋落。期间分别记录花朵的组成结构,各器官生长变化的特征、时间及过程,特别关注花瓣伸展、柱头伸长、柱头弯曲、花粉散开、花朵萎蔫、花蜜与气味开始和持续时间等情况。选择标记的花朵进行单花开放动态观察并拍照。于盛花期,随机选取正开放的 30朵花,测量花冠直径、花冠筒长、花被片长度以及雌、雄蕊等各部分结构的长度。
1.2.2 花粉活力及柱头可授性的测定
分别于开花1、6、12、18、24、36、48 h随机选取 15朵开花进程一致的花朵扫取花粉到硫酸纸上,均匀混合撒于最适培养基上,28 ℃恒温箱中进行花粉离体培养。每个时期选取5个视野,以花粉管的长度大于花粉粒的直径为萌发标准,分别统计每个视野中萌发花粉比例。萌发率=已萌发花分数/观测花分数。观察单花开放进程中柱头的变化情况及黏液分泌情况,同时采用联苯胺-过氧化氢测定不同时期的柱头的可授性[18]。开花前2 d每天测定1次,开花后每6 h测定1次,直至花朵凋落。每个时期检测单花15朵,记录检测结果。
1.2.3 繁育系统的检测
采用Cruden[19]方法测定单花花粉量和胚珠数,估算花粉胚珠比(P/O)。根据Dafni[20]的标准对北黄花菜的直径、开花行为进行测定,计算杂交指数(OCI)。
1.2.4 坐果率的测定
对北黄花菜进行8种不同授粉方式处理:不去雄,不套袋,自由传粉;去雄,套袋,不授粉;去雄,不套袋,自由传粉;去雄,套袋,人工授自花花粉;去雄,套袋,人工授同株异花花粉;去雄,套袋,人工授异株花粉;不去雄,套袋,自花授粉;去雄,细白纱网套袋,自由传粉。每个处理取 30朵花,统计不同处理的坐果率及结籽率。坐果率=坐果数/处理单花数;结籽率=结籽数/总胚株数。
2 结果与分析
2.1 北黄花菜的花部综合特征
北黄花菜为多年生宿根草本。叶片基部抱茎生长,线形深绿色,全缘,先端急尖,叶长 30~90 cm,叶宽3~12 mm。扫描电镜观察发现,北黄花菜的叶片表皮外壁加厚不明显,有高突部分出现,气孔分布较密(图1-1和图1-2)。北黄花菜花葶高50~80 cm,直立,高于叶丛生长。假二歧状总状或圆锥花序,每花序上着生1~7朵花不等,以4~6朵居多。花淡黄色或黄色,芳香。花瓣6枚,为内外2轮各3枚,外轮长而窄,内轮短而宽。花呈筒状,花裂片向外反卷。花朵开放时,花冠筒长16.34~25.74 mm。外轮 3枚花裂片长 45.38~70.42 mm,宽 7.36~19.82 mm。内轮 3枚花裂片长为42.72~68.98 mm,宽8.34~19.52 mm。花冠直径为65.74~118.66 mm。雄蕊有 6枚,其花丝长为30.42~48.48 mm。花药初为灰褐色,逐渐变为紫褐色,最终变为黑色,长为 3.52~5.58 mm,宽为0.52~2.86 mm。花柱高63.62~79.54 mm,黄色。柱头高 0.46~1.58 mm,白色,表面有乳状突起,成熟后分泌黏液。北黄花菜的子房呈三棱柱状,基部较大,有明显的纵棱。扫描电镜观察发现,子房外壁表面呈鳞片状,组织致密。子房外部下方可见较大孔洞(图1-3),解剖见大量的花蜜流出。
扫描电镜观察发现,北黄花菜花粉为单粒花粉,形状为长球形(图1-4),极面观为椭圆形,赤道面观为舟形。花粉粒大小为102.49 µm×35.35 µm。依据Erdtman[21]划分标准,此类花粉属于非常大孢粉。萌发器官为远极单沟,萌发沟细而深,两侧对称(图 1-5)。外壁具有清晰深刻的网状雕纹,网脊近粗糙,网眼大小不一,内部分布有颗粒状次级雕纹。近两端雕纹变浅,网纹消失(图 1-6)。北黄花菜柱头为三角形(图1-7和图1-11),相对表面积较大,表面有大量的柱状凸起(图 1-8和图1-10),成熟的柱头可以分泌大量的黏液,极容易黏附嵌入花粉(图 1-9)。相关分析结果表明,北黄花菜花粉粒的大小与花柱高度、花冠直径均呈极显著正相关,相关系数分别为0.811和0.841,说明花冠直径越大,花柱越高的单花花粉粒越大。
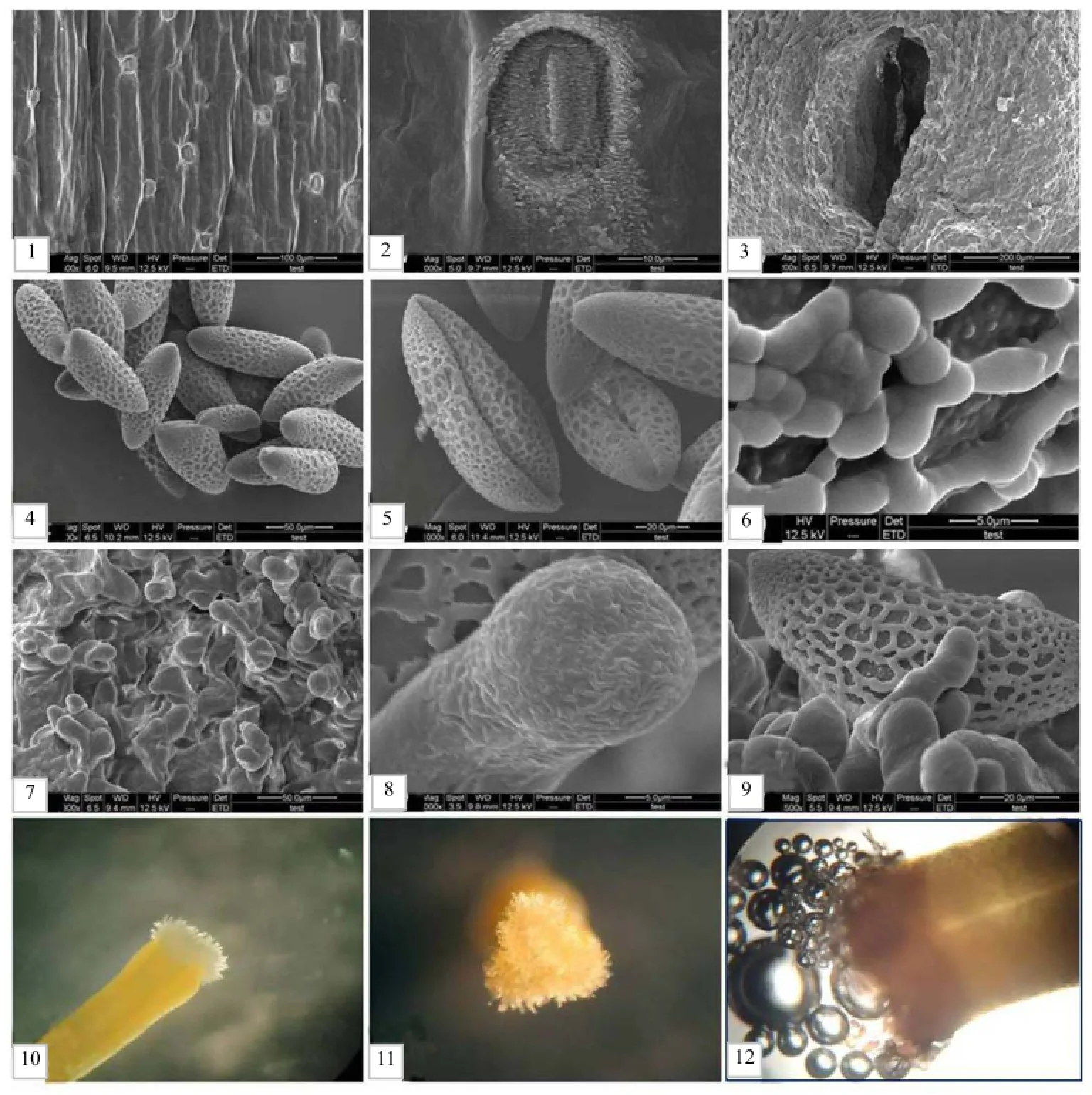
图1 北黄花菜部分器官的超微结构及显微结构Fig. 1 Microstructure and ultrastructure of part organs of H. lilio-asphodelus
2.2 北黄花菜的单花开放动态
北黄花菜现蕾时,苞片为绿色,多数在高出叶丛时苞片已被涨破,呈现出大小不等的2片,露出内部花蕾。花蕾初为绿色,开花前变为黄绿色。北黄花菜常在13:00—18:00开花。开花过程中,外轮的花裂片先松动,以减少顶端的封锁力,内轮花裂片也随之开始伸展,与外轮花瓣紧贴保持一致。6枚花裂片完全伸展大约需要6~8 h。开花时柱头白色,明显高于花药,只有少数柱头变异种柱头低于花药。开花1~6 h左右柱头有少量黏液分泌。开花8 h左右花柱弯向花药生长。花丝在开花过程中无明显变化,花药在接触空气20~30 min内即可开裂,基本无先后顺序,开裂持续时间大约 6~8 h,花药开始散粉。开花第2天早晨花裂片伸展幅度达到最大,柱头颜色变为乳白色或乳黄色,肉眼可见柱头的乳状突起,分泌大量黏液,柱头与花药之间的距离进一步缩小。开花 20 h左右花粉大量散落。开花第2天下午花瓣边缘出现萎蔫内卷,质地变透明。开花第3天早晨花冠已经闭合,基部拧卷收缩,花裂片开始失水枯萎,但仍然宿存直至果实膨大将其顶落。此时花柱变细,柱头失水收缩,体积变小。花药扭曲干枯,但仍有少量的花粉存留。北黄花菜的单花开放动态过程见图2。统计发现,北黄花菜95%以上的单花持续时间为2 d,不到5%的单花持续时间只有1 d。

图2 北黄花菜单花开放动态Fig. 2 Development of flower opening of H. lilio-asphodelus
2.3 北黄花菜的花粉活力及柱头可授性
北黄花菜开花1 h左右花药已经开裂,此时花粉已有活力,但是活力较低,在随后的18 h 内活力呈上升趋势,到12~18 h活力最大,此时是授粉的最佳时期。随后直至36 h,活力逐渐下降,少许花粉有生活力,直至花瓣闭合,花粉丧失生活力(表1)。

表1 北黄花菜的花粉活力、柱头可授性及黏液分泌情况Table 1 Pollen viability, Sigma receptivity and mucus secretion of H. lilio-asphodelus
具有可授性的柱头浸于联苯胺-过氧化氢反应液中会呈现紫黑色或蓝色,并有大量气泡(图1-12)。检测结果发现,北黄花菜开花前1~2 d部分柱头已经有可授性;开花后 1~6 h,柱头具有可授性,此时柱头呈三角状膨大;开花后6~12 h,柱头颜色变浅,柱头乳状突起明显伸长,开始有分泌物出现,可授性增强;直至开花后12~18 h,柱头颜色为乳白色,大量分泌黏液,可授性最强;开花后24~36 h柱头变为黄白色,乳状突起部分已经脱落,此时仍有少量黏液分泌,可授性较强;开花后36~48 h花瓣趋于败落,此时柱头变小,为黄色,无黏液分泌,少量柱头仍具有可授性。开花 48 h后柱头完全干枯,黄褐色,无可授性。
2.4 北黄花菜繁育系统的确定
2.4.1 花粉胚珠比(P/O)
北黄花菜的单花花粉数约为93 760粒,单花胚珠数约为51个,P/O约为1 838。根据Cruden[19]的划分标准,该物种的繁育系统属于兼性异交类型。
2.4.2 杂交指数(OCI)
北黄花菜的花朵直径在65.74~118.66 mm,大于6 mm,OCI为3。在花药散粉初期雌蕊已成熟,认为雌雄器官在成熟时间上一致,OCI为0。柱头和花药虽逐渐靠近,但柱头始终高于花药且未接触,在位置上存在空间分离,OCI记为1。综合以上结果,确定北黄花菜的OCI值为4。根据Dafni[20]提出的标准,北黄花菜的繁育系统为部分自交亲和,异交,需传粉者。
2.4.3 授粉方式对北黄花菜坐果率和单果结籽率的影响
通过人工授粉和套袋试验来检测北黄花菜的授粉方式。结果(表 2)显示,去雄、套袋、不授粉条件下坐果率为 0,说明北黄花菜不存在无融合生殖现象。在自然条件下北黄花菜的坐果率很高,达到82.93%,但是结籽率较低,仅为32.76%,经过人工自花授粉、人工异花授粉后坐果率均显著提高,人工授同株异花花粉后坐果率无显著变化,但结籽率明显提高。套袋试验结果也表明,北黄花菜自花可孕,但需要传粉者进行异花传粉。

表2 不同授粉方式下北黄花菜的坐果率和单果结籽率Table 2 The test results of different pollinating ways of H. lilio-asphodelus
3 结论与讨论
依据 Cruden的评判标准,北黄花菜的繁育系统为兼性异交;按照Dafni的判断标准,北黄花菜的繁育系统为异交,部分自交亲和,需要传粉者。本研究中不同授粉方式试验结果显示,去雄、套袋处理条件下及不去雄、套袋处理条件下北黄花菜的结实率均为0,表明北黄花菜不存在无融合生殖和自动自花授粉。雌雄异位、雌雄异熟、自交不亲和是促进异交,避免自交的重要机制[22-23]。北黄花菜花药和柱头存在空间分离,自然条件下自交受限制,人工自花授粉和人工异花授粉后北黄花菜的坐果率均达到 90%以上,表明其自交亲和,且亲和程度很高。
在自然条件下北黄花菜的坐果率即能达到80%以上,而在去雄、自由传粉情况下坐果率不到30%,可能的原因为:花粉为昆虫的主要传粉报酬,缺少花粉的单花不容易获得访花者的到访;北黄花菜不存在风媒传粉途径,这一点通过白纱网套袋处理后的坐果率也已得到证明。北黄花菜自然条件下结实率较高,但结籽率低,人工自花授粉和人工同株异花授粉后结籽率无显著变化,异花授粉后结籽率显著提高,表明自然条件下北黄花菜应该是被动自花传粉和异花传粉共同作用。这也说明传粉者在北黄花菜的生殖成功中具有极其重要的媒介作用。如何提高自然条件下异花授粉机率、提升结籽率是进行种子生产、良种选育的前提。传粉昆虫的种类、数量、访花频率、传粉效率及其与植株的相互作用等需要进一步的深入研究。
花粉活力的高低及柱头可授性的强弱是植物生殖成功与否、结实率高低的关键因素之一。柱头在整个花期都具有可授性,花粉活力和柱头可授性在开花期间均呈先增长后降低的单峰曲线,且两者的活力最高峰存在相遇期,这为被动自交提供了可能性。北黄花菜的花粉活力下降速度太快,在散粉48 h左右活力几乎完全丧失,对于传粉昆虫的远距离传粉及居群间的基因交流是不利的。
参考文献:
[1] 中国科学院中国植物志编辑委员会.中国植物志.第14卷[M].北京:科学出版社,1980.
[2] 邓放明,尹华,李精华,等.黄花菜应用研究现状与产业化开发对策[J].湖南农业大学学报(自然科学版),2003,29(6):529-532.
[3] Wyatt R.Pollination Biology [M].Orlando:Academic Press,1983.
[4] 何亚平,刘建全.植物繁育系统研究的最新进展和评述[J].植物生态学报,2003,27(2):151-163.
[5] 陈雄伟,邵玲,梁广坚,等.紫背天葵花部特征与繁育系统的研究[J].园艺学报,2013,40(2):363-372.
[6] 陈香,胡雪华,肖宜安,等.紫茉莉的花部综合特征与繁育系统[J].生态学杂志,2008,27(10):1653-1658.
[7] 胡春,刘左军,伍国强,等.钝裂银莲花花部综合特征及其繁育系统[J].草地学报,2013,21(4):783-788.
[8] 刘芬,李全健,王彩霞,等.濒危植物扇脉杓兰的花部特征与繁育系统[J].林业科学,2013,49(1):53-60.
[9] 杨琨.植物繁育系统中传粉机制的多样性及交配系统研究评析[J].西安文理学院学报(自然科学版),2012,15(3):41-48.
[10] 刘开全,邓洪平.重庆特有濒危植物缙云黄岑的繁育系统研究[J].植物研究,2011,15(3):41-48.
[11] 王汉海,程贯召,杜延飞,等.大花萱草新品种‘金娃娃’的组织培养和快速繁殖[J].植物生理学通讯,2002,38(5):458-461.
[12] 崔贵梅,杨丽莉,孙毅,等.大花萱草‘金娃娃’雌雄蕊发育进程及其与花蕾长度的相关性研究[J].西北植物学报,2008,28(3):41-48.
[13] 王晓娟,金樑,陈家宽.萱草的组织培养与快速繁殖[J].植物生理学通讯,2003,39(3):234-236.
[14] 朱云华,苏倩,武文婷,等.萱草属野生资源遗传距离和多态性分析[J].南京林业大学学报(自然科学版),2007,31(6):97-100.
[15] Hasegawa M,Yahara T,Yasumoto A,et al.Bimodal distribution of flowering time in a natural hybrid population of daylily (Hemerocallis fulva) and nightlily (Hemerocallis citrina)[J].Journal of Plant Research,2006,119(1):63-68.
[16] Hiroyuki Satio,Keiko Mizunashi,Shigefumi Tanaka,et al.Ploidy estimation in Hemerocallis species and cultivars by flow cytometry[J].Scientia Horticulturae,2003,97:185-192.
[17] 赵天荣,蔡建岗,施永泰,等.大花萱草‘金娃娃’的花器结构和繁育系统观察[J].草业科学,2013,30(1):52-57.
[18] 叶要妹,张俊卫,齐迎春,等.百日草柱头可授性和花粉生活力的研究[J].中国农业科学,2007,40(10):2376-2381.
[19] Cruden R W.Pollen-ovule ratios:a conservative indicator of breeding system in flowering plants[J]. Evolution,1977,31:32-46.
[20] Dafni A.Pollination Ecology:A Practical Approach[M]. New York:Oxford University Press,1992.
[21] Erdtman G.Handbook of Palynology,Morphology,Taxonomy Ecology[M].Copenhagen:Munksgaard,1969.
[22] 刘宗才,焦铸锦,董旭升,等.鸢尾的花部结构及繁育系统特征[J].园艺学报,2011,38(7):1333-1340.
[23] 侯勤正,段元文,司庆文,等.青藏高原晚期开花植物线叶龙胆的传粉生态学[J].植物生态学报,2009,33(6):1156-1164.
责任编辑:尹小红
英文编辑:梁 和
中图分类号:S682.1+90.1
文献标志码:A
文章编号:1007-1032(2016)01-0058-06
收稿日期:2014-11-17 修回日期:2015-11-22
基金项目:中央高校基本科研业务费专项资金项目(DL13BA08)
作者简介:孙颖(1979—),女,吉林长春人,博士,副教授,主要从事园林植物种质资源开发与利用研究,littlesuning@126.com;*通信作者,何淼,博士,副教授,主要从事园林植物培育、养护管理研究,hemiao_xu@126.com和北黄花菜(Hemerocallis lilio-asphodelus)[1]。北黄花菜花色明亮艳丽,花形美观可爱,花具芳香,观赏价值较高,同时其耐寒性强,作为高寒地区的园林景观植物有很好的发展前景,是极具开发潜力的野生草本种质资源[2]。
Floral characters and breeding system of Hemerocallis lilio-asphodelus
SunYing, Yang Xue, Wang Axiang, He Miao*
(College of Landscape Architecture, Northeast Forestry University, Harbin 150040, China)
Abstract:Hemerocallis lilio-asphodelus, a perennial herb of Liliaceae, with brilliant flower color, marked flower shape,ornamental and medicinal value, was used as experiment material. The experiments of quantitative measurement, pollen germination, emasculation, bagging and artificial pollination were conducted to study the flowering character, pollen viability, stigma receptivity and breeding system of H. lilio-asphodelus. The surface pattern of stigma and pollen were observed by an electron microscope as well. The results showed that: H. lilio-asphodelus had large fragrant flowers and the individual flower longevity was about 2 days. Floral character could attract pollinators. The pollen viability gradually increased and reached to the highest at about 12 h to 18 h after flowering, and the stigma had pollen receptivity from the beginning of bloom and lasted the whole flowering period. The stigma exceeded stamen in position. The total value of outcrossing index (OCI ) was 4 and the value of pollens and ovules was about 1 838. The rate of setting fruit reached 82.93% under natural conditions. With artificial self-pollination and cross-pollination, the rate of setting fruit was 96.68% and 94.32%, respectively. The rate of setting fruit with artificial geitonogamy was 83.45%. In conclusion, the breeding system of H. lilio-asphodelus could be determined as facultative outcrossing, self-compatible and pollinators dependent and have no syngamy.
Keywords:Hemerocallis lilio-asphodelus; floral character; breeding system; outcrossing index; pollen-ovule ratio; pollen viability; stigma receptivity
