洞头列岛及邻近海域大型底栖动物群落结构的研究
2016-07-26贾胜华曾江宁廖一波高爱根汤雁滨
贾胜华,曾江宁*,廖一波,寿 鹿,黄 伟,高爱根,汤雁滨
(1.国家海洋局 海洋生态系统与生物地球化学重点实验室,浙江 杭州 310012;2. 国家海洋局 第二海洋研究所,浙江 杭州 310012)
洞头列岛及邻近海域大型底栖动物群落结构的研究
贾胜华1,2,曾江宁*1,2,廖一波1,2,寿鹿1,2,黄伟1,2,高爱根1,2,汤雁滨1,2
(1.国家海洋局 海洋生态系统与生物地球化学重点实验室,浙江 杭州 310012;2. 国家海洋局 第二海洋研究所,浙江 杭州 310012)
摘要:为了解洞头列岛及邻近海域大型底栖动物的群落结构特征,于2015年4月在该海域布设35个站位进行大型底栖动物调查采样,并获取相关环境资料,分析了该海域大型底栖动物的群落结构及其与环境因子的关系。共鉴定出大型底栖动物122种,其中多毛类57种,甲壳动物34种,软体动物19种,棘皮动物5种,其它类动物7种。各调查站位大型底栖动物物种数目在1~22种之间,瓯江口附近站位物种数较少。大型底栖动物平均生物量为3.35 g/m2,平均栖息密度为321个/m2。多毛类具有种数上的优势,软体动物具有生物量和密度上的优势。Shannon-Wiener多样性指数、Margalef丰富度指数和Pielou均匀度指数的平均值分别为1.29、2.45和0.66。通过聚类分析和多维尺度排序分析将该海域大型底栖动物划分为3个群落:异蚓虫-薄云母蛤-绒毛细足蟹-不倒翁虫群落(群落Ⅰ),薄云母蛤-异蚓虫-双鳃内卷齿蚕-不倒翁虫群落(群落Ⅱ)和其他群落(群落Ⅲ)。典范对应分析表明,影响大型底栖动物群落的主要环境因子有底层海水的温度、盐度、活性磷酸盐、活性硅酸盐、溶解性无机氮、化学需氧量以及表层沉积物中的总有机碳等。ABC曲线分析结果表明,大型底栖动物群落受到了一定程度的扰动,但基本稳定。与历史资料相比,大型底栖动物的栖息密度和生物量均无明显变化,种类组成由以多毛类和软体动物为主转变为以多毛类和甲壳动物为主,且个体较大的棒锥螺大量死亡。
关键词:大型底栖动物;群落结构;生物多样性;环境因子;洞头列岛及邻近海域
0引言
海洋大型底栖动物在评价海洋底栖生态环境质量和反映人类活动干扰中具有明显的优势,一是大型底栖动物与沉积物环境的距离最近——生活在海洋底层,直接与沉积环境接触;二是大型底栖动物暴露在沉积环境的时间最长——生活范围有限,多数种类的成体终生栖息在固定场所或只能在底质表面有限范围内活动[1],某些最终会沉降到海洋底层的污染物如重金属[2-3]、多环芳烃(PAHs)[4]等会对其长期作用;三是大型底栖动物本身对物理化学扰动非常敏感[5],其种类和丰度减少或群落结构发生变化是对环境扰动或污染的响应[6]。这些特点使大型底栖动物在深入研究底栖生态环境质量状况中具有重要的作用,并且越来越受到广大学者以及环境管理者的关注。
洞头列岛(121°10′E,27°50′N)位于瓯江口和乐清湾口交汇的浙江东南海域,由168个岛屿组成。海洋生物资源丰富,洞头渔场是仅次于舟山渔场的浙江省第二大渔场[7]。该海域有浙江省第二大河流——瓯江注入,浙南沿岸流、东海外海水与台湾暖流在此进行水交换,瓯江入海泥沙扩散加上岛屿本身复杂的地形特征,使得该海域大型底栖动物群落具有独特的特征。然而近年来随着社会经济的发展,尤其是温州半岛工程实施以来,滩涂围垦工程、跨海大桥工程以及养殖活动、污水排放和港口航运等使该海域的生态环境发生了一定的变化,给该海域的大型底栖动物带来了威胁[8]。目前,有关洞头大型底栖动物的报道多集中于潮间带[8-10],只限于局部较小范围[11-14],鲜有对该海域大范围大型底栖动物的报道。本文研究了洞头列岛及邻近海域35个调查站位大型底栖动物的群落结构特征,并探讨其与环境因子的关系,以期为探明该海域大型底栖动物的资源现状和评价底栖生态环境质量状况提供科学依据。
1材料与方法
1.1样品采集与室内分析
于2015年4月在洞头列岛及邻近海域进行大型底栖动物采样和环境因子调查,共布设35个采样站位(图1)。采用0.1 m2的Van Veen 型抓斗式采泥器采集样品,每站重复取样2次,合并为1个样品。采用孔径为0.5 mm的网筛分选样品,获得的生物样品用5%的中性福尔马林溶液现场固定,带回实验室进行种类鉴定、计数和称重(湿重)。样品的采集、分析、称重、计算和资料整理均按《海洋调查规范》(GB/T 12763.6—2007)[15]的要求进行。
1.2环境因子的测定
本次调查的海洋底层水体环境因子有:温度、盐度、pH、化学需氧量、溶解氧、活性磷酸盐、活性硅酸盐、溶解性无机氮和悬浮物;表层沉积物环境因子有:总有机碳和总氮,同时记录每个站位的水深。环境因子调查过程中样品的采集、贮存、运输、预处理及分析测定过程均按《海洋调查规范》(GB/T 12763.6—2007)[15]和《海洋监测规范》(GB 17378.4—2007)[16]的要求进行。
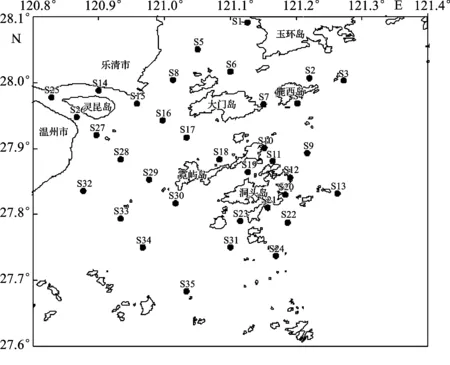
图1 洞头列岛及邻近海域大型底栖动物采样站位Fig.1 Sampling stations of macrozoobenthos in Dongtou Islands and adjacent sea area
1.3数据处理
采用Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(d)和Pielou均匀度指数(J′)[17]来分析大型底栖动物群落的生态特征,其计算公式分别为:
(1)
d=(S-1)/lnN
(2)
J′=H′/lnS
(3)
式中:Pi为第i种的个数与该样品总个数的比值,S为样品中的总种类数,N为样品中所有种类的总个体数。
采用物种优势度指数(Y)[17]确定大型底栖动物的优势种:
Y=Pi×fi
(4)
式中:fi为该种在各站位出现的频率。当Y>0.02时,确定该物种为优势种。
以Bray-Curtis相似性系数为基础对大型底栖动物群落进行等级聚类分析和非度量多维尺度排序分析[18-19]:首先对密度数据进行平方根转换,以便对稀有种(小个体种)给予一定程度的加权;然后应用PRIMER 6.0软件计算每个站位物种之间的Bray-Curtis相似性系数,并构建相似性矩阵,采用类平均法进行等级聚类分析并得到聚类树枝图,使相似矩阵变得直观;再通过非度量多维尺度排序分析将站位之间的相似关系转化为距离反映在二维平面上,使群落间的关系更加直观,但其具有随意性。相似矩阵、聚类分析与MDS排序结合使用,反映群落间的关系具有准确和直观的优点[20]。采用典范对应分析(Canonical Correspondence Analysis,CCA)对大型底栖动物群落特征与环境因子的关系进行分析,该分析在Canoco for Windows 4.5中完成,并利用Monte Carlo检验CCA排序中所有轴的显著性。采用丰度/生物量比较曲线(Abundance/Biomass Curves, ABC曲线)分析洞头海域大型底栖动物群落结构的稳定性[19],根据ABC曲线中丰度/生物量的部分优势度曲线和累积优势度曲线可分析研究海域大型底栖动物受扰动的状况。
2结果与分析
2.1种类组成与分布
本次调查共获得大型底栖动物122种,其中多毛类57种,甲壳动物34种,软体动物19种,棘皮动物5种,其它类动物7种。多毛类和甲壳动物是该海域大型底栖动物群落的主要类群,两者占总物种数的74.6%。大型底栖动物优势种有4种,分别是薄云母蛤Yoldiasimilis、异蚓虫Heteromastinsfiliformis、不倒翁虫Sternaspisscutata和双鳃内卷齿蚕Aglaophamusdibranchis,优势度分别为0.34,0.05,0.03和0.02,栖息密度分别为165,24,15和12个/m2。习见种为圆筒原盒螺Eocylichnabraunsi、寡节甘吻沙蚕Glycindegurjanvae、绒毛细足蟹Raphidopusciliatus、长吻吻沙蚕Glycerachirori、多鳃齿吻沙蚕Nephtyspolybranchia、小头虫Capitellacapitata和钩虾Gammarideasp.。
各调查站位大型底栖动物物种数在1~22种之间,并且以多毛类、软体动物和甲壳动物为主(图2)。平均每个站位有大型底栖动物3.49种,S11站位的物种数最多,达22种,S24站位有21种,这2个站位均位于洞头海域东部。而处于瓯江口海域的站位S14、S15、S25和S28物种数较少,分别只有1种。从总体分布上看,大型底栖动物的物种数在瓯江口与乐清湾口附近的站位较少,而在东部和南部海域的站位较多;且棘皮动物只出现在洞头东部和南部海域,包括S4、S19、S21、S23、S31、S32、S33、S34和S35站位,以S35站位为中心向北呈“V”形分布(图3),瓯江口与乐清湾口海域附近站位没有出现。

图2 大型底栖动物的种类数目分布Fig.2 Distribution of species number of macrozoobenthos
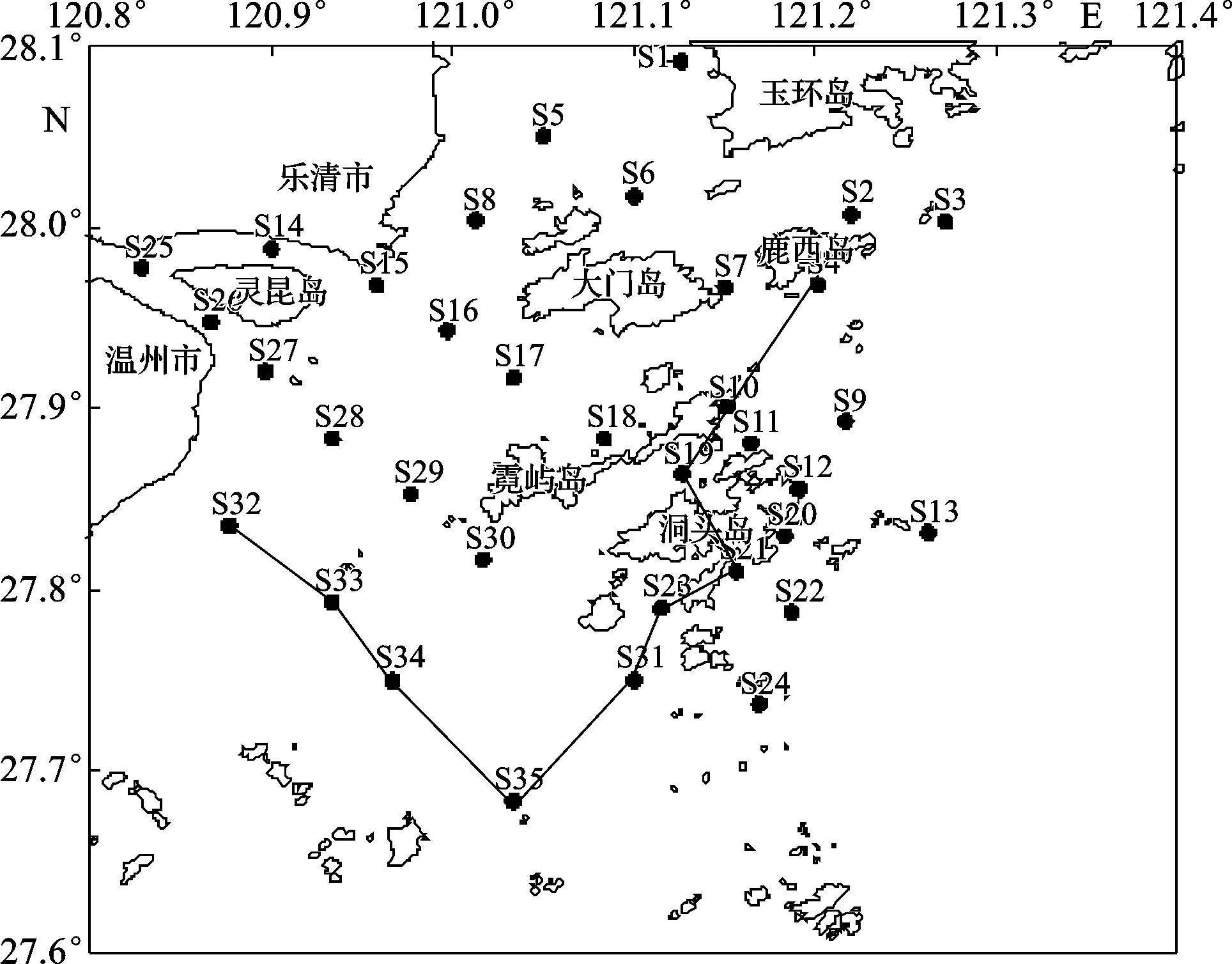
图3 棘皮动物的空间分布Fig.3 Spatial distribution of Echinodermata
2.2数量组成与分布
调查海域春季大型底栖动物平均生物量为3.35 g/m2,平均栖息密度为321个/m2(表1)。软体动物的生物量和栖息密度最大,分别占该海域总生物量和总栖息密度的63.4%和64.0%;多毛类位居第二位,分别占总生物量和总栖息密度的21.7%和29.1%;与甲壳动物相比,棘皮动物栖息密度低,但生物量高,原因是某些站位的蛇尾个体很大,如S4站位的滩栖阳遂足Amphiuravadicola可达到1.66 g/个,而甲壳动物中主要物种为钩虾和涟虫,占82.4%,它们一般个体很小,湿重均不超过0.005 g/个。

表1 大型底栖动物各类群的生物量和栖息密度
大型底栖动物生物量和栖息密度分布见图4。大型底栖动物生物量分布范围为0.01~27.70 g/m2,高值出现在S4和S27站位,分别为13.59 g/m2和27.70 g/m2。S4站位生物量的主要构成者为短脊鼓虾Alpheusbrevicristatus、绒毛细足蟹和滩栖阳遂足,三者占该站位生物量的94.4%;S27站位生物量的主要构成者为焦河篮蛤Potamocorbulaustulata,占该站位生物量的98.8%。生物量低值出现在S14和S28站位,均低于0.02 g/m2。大型底栖动物栖息密度分布范围为5~1 445个/m2,高值出现在S33站位和S35站位,分别为1 445个/m2和1 010个/m2。S33站位和S35站位栖息密度的主要构成者均为薄云母蛤,分别占对应站位栖息密度的85.8%和35.6%。最低值出现在S14、S15和S28站位,均为5个/m2。S27站位与其他站位明显不同,该站位出现大量焦河篮蛤,密度达630个/m2,且个体较大,可达1.3 g/个。总体上看,瓯江口附近海域以及瓯江口与洞头列岛之间海域大型底栖动物的栖息密度和生物量较低,洞头列岛东南部海域大型底栖动物的栖息密度和生物量高于西北部海域。
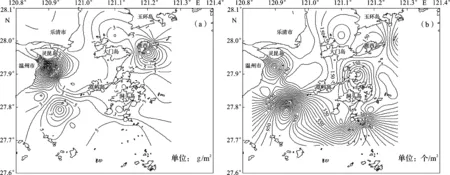
图4 大型底栖动物生物量(a)和栖息密度(b)的空间分布Fig.4 Spatial distribution of macrozoobenthic biomass(a) and abundance(b)
2.3群落结构特征
采用Primer 6.0软件对调查海域35个采样站位的大型底栖动物数据进行聚类分析,并构建非度量多维尺度排序图(图5和图6)。由图5可知,站位S1和站位S5的相似性最高,为60.7%。根据Bray-Curtis相似性系数聚类分析和非度量多维尺度排序结果,并综合考虑调查站位所处海域的地理位置以及海流、水深、盐度和沉积物等特征,可将该海域大型底栖动物划分为3个群落:
异蚓虫-薄云母蛤-绒毛细足蟹-不倒翁虫群落(群落Ⅰ):该群落位于洞头海域西北部,乐清湾口正对面海域,包括站位有S7、S8、S12、S18和S22,平均水深为13.84 m。大型底栖动物共有42种,平均生物量和栖息密度分别为4.66 g/m2和249个/m2。其中多毛类的生物量和栖息密度最大,分别为1.25 g/m2和127个/m2。优势种为异蚓虫、薄云母蛤、绒毛细足蟹和不倒翁虫。
薄云母蛤-异蚓虫-双鳃内卷齿蚕-不倒翁虫群落(群落Ⅱ):该群落相似性较高,优势种明显,其中薄云母蛤的优势度可达到0.43。主要位于洞头东部和南部海域,受东海外海水和台湾暖流影响,包含范围比较大,包括站位有S1、S2、S3、S4、S5、S6、S9、S10、S11、S13、S17、S19、S20、S21、S23、S24、S30、S31、S33、S34和S35,平均水深为13.28 m。大型底栖动物共有92种,平均生物量和栖息密度分别为2.82 g/m2和395个/m2。该群落中多毛类种类数目最多,达42种,但软体动物的生物量和栖息密度最大,分别为1.90 g/m2和266个/m2。优势种为薄云母蛤、异蚓虫、双鳃内卷齿蚕和不倒翁虫。
其他群落(群落Ⅲ):该群落相似性很低(低于20%),差异很大,因此归为其他群落。位于瓯江口和洞头海域西南部,包括S14、S15、S16、S25、S26、S27、S28、S29和S32站位,平均水深为7.01 m。大型底栖动物共有44种,平均生物量和栖息密度分别为3.96 g/m2和187个/m2。其中软体动物的生物量和栖息密度最大,分别为3.57 g/m2和134 个/m2。该群落的生物量主要受焦河篮蛤影响,本次调查在瓯江口南侧的S27站位出现了大量的焦河篮蛤,且个体较大,若除去该种,则群落Ⅲ的生物量只有0.92 g/m2。优势种为薄云母蛤和异蚓虫。

图5 大型底栖动物群落等级聚类图Fig.5 The dendrogram for cluster of macrozoobenthic community
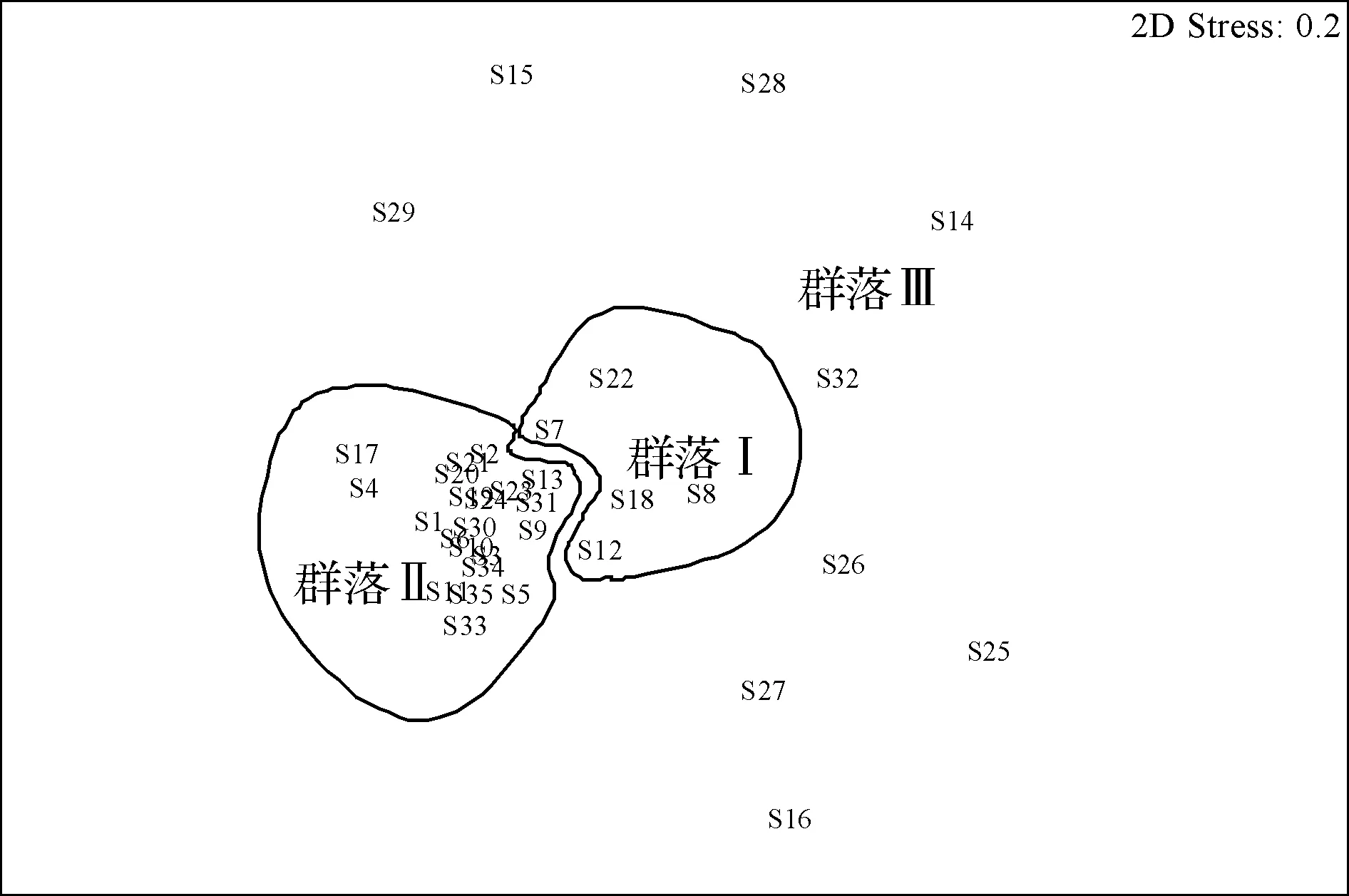
图6 大型底栖动物群落多维尺度排序分析Fig.6 The MDS analysis of macrozoobenthic community
2.4群落生态特征
对调查海域大型底栖动物群落物种多样性指数(H′)、物种丰富度指数(d)和均匀度指数(J′)进行计算,结果见表2。由表2可以看出,不同站位大型底栖动物物种多样性指数、物种丰富度指数和均匀度指数波动范围较大。物种多样性指数H′范围在0~2.34,平均值为1.29。丰富度指数d范围在0~4.52,平均值为2.45。均匀度指数J′范围在0.15~1.00,平均值为0.66。群落Ⅱ的H′和d平均值均最高;群落Ⅲ的H′平均值最低,但J′平均值最高;群落Ⅰ的d平均值最低。
2.5大型底栖动物群落与环境因子的关系
大型底栖动物群落与环境因子的CCA分析结果如图7所示。Monte Carlo显著性检验结果表明,CCA所有排序轴均呈显著性差异(P<0.05)。排序轴1~4可解释物种-环境关系的74.5%。大型底栖动物群落与第1轴呈较强正相关性的环境因子为活性磷酸盐(r=0.84)、溶解性无机氮(r=0.82)、活性硅酸盐(r=0.70)、化学需氧量(r=0.62)和温度(r=0.59),呈较强负相关性的环境因子为盐度(r=-0.91);与第2轴呈较强负相关性的环境因子为总有机碳(r=-0.42)。
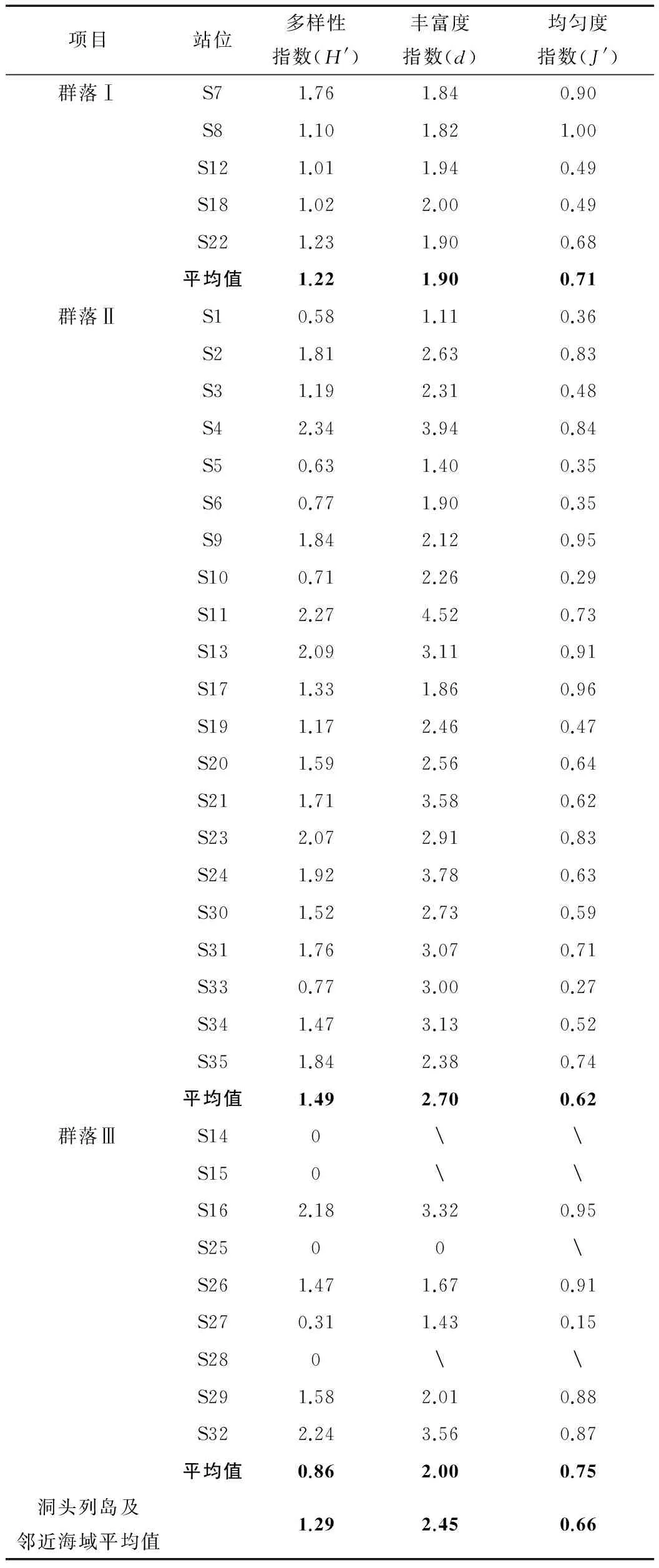
表2 大型底栖动物群落的生态特征
注:“”表示该站位为单一物种,且个数为1,对应值无法计算。

图例:T—温度;D—水深;S—盐度;pH—酸碱度;COD—化学需氧量; DO—溶解氧;P—活性磷酸盐;Si— 活性硅酸盐;DIN—溶解性无机氮;SS—悬浮物;TOC—总有机碳;TN—总氮
1—长吻吻沙蚕Glycerachirori; 2—寡节甘吻沙蚕Glycindegurjanvae; 3—双鳃内卷齿蚕Aglaophamusdibranchis; 4—多鳃齿吻沙蚕Nephtyspolybranchia; 5—寡鳃齿吻沙蚕Nephtysoligobranchia; 6—双形拟单指虫Cossurelladimorpha; 7—不倒翁虫Sternaspisscutata; 8—异蚓虫Heteromastinsfiliformis; 9—内肋蛤Endopleuralubrica; 10—薄云母蛤Yoldiasimilis; 11—日本镜蛤Dosiniajaponica; 12—圆筒原盒螺Eocylichnabraunsi; 13—钩虾Gammarideasp.; 14—绒毛细足蟹Raphidopusciliates; 15—四齿蛇尾Paramphicondriustetradontus; 16—背蚓虫Notomastuslatericeus
图7大型底栖动物主要种类与环境因子的CCA排序图
Fig.7CCA ordination diagram of macrozoobenthic community
between major species and environmental factors
2.6大型底栖动物的群落结构稳定性
根据调查海域大型底栖动物丰度和生物量数据作出丰度/生物量部分优势度曲线和累积优势度曲线(图8)。结果表明,部分优势度曲线出现交叉、翻转,两条曲线多次相互穿过;累积丰度优势度曲线和生物量优势度曲线在末端交叉,两条曲线紧靠在一起,说明群落受到一定程度的扰动。但生物量曲线起点均高于丰度曲线,说明大型底栖动物群落基本稳定。该分析结果与本次调查的大型底栖动物种类组成特征相吻合:群落物种多是个体较小的多毛类、软体动物和甲壳动物,在部分站位出现个体较大的焦河篮蛤、虾、滩栖阳遂足,而在瓯江口海域多数站位底栖动物数量少且生物量很低。
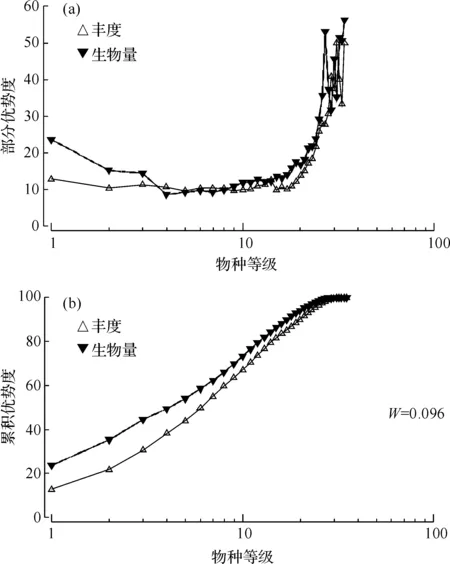
图8 大型底栖动物丰度/生物量的部分优势度曲线(a)和累积优势度曲线(b)Fig.8 Partial dominance curves (a) and cumulative dominance curves (b) of macrozoobenthic abundance/biomass
3讨论
3.1大型底栖动物的历史演变
研究海域大型底栖动物的种类组成变化见表3。由表3可以看出,近25 a来,该海域的大型底栖动物种类组成发生了一定的改变。1990年到2007年,多毛类所占比例几乎不变,软体动物所占比例由28.4%上升至55.0%,而甲壳动物所占比例由21.1%下降至5.0%。近25 a来,大型底栖动物种类组成变化比较显著:1990年和2007年调查均为多毛类和软体动物占有最大比例,而本次调查是多毛类和甲壳动物占有最大比例。这是由于本次调查出现大量个体较小的钩虾,导致甲壳动物物种数较多。钩虾的大量出现可能有两方面的原因:一是春季是一些种类钩虾的繁殖季节,如一种蜾蠃蜚Corophiumsp.抱卵雌体出现的高峰期为3 月和4月期间[21];二是可能与近年来洞头列岛的养殖活动发展有关,鱼排区或大型海藻养殖区数量的增多,为钩虾提供了丰富的饵料和适宜的生存环境。
大型底栖动物平均生物量和栖息密度的变化见表4。本次调查大型底栖动物平均生物量和栖息密度和1990年调查结果接近。2006年调查中出现了一定量的棒锥螺,其外壳占有相当重量,导致大型底栖动物的平均生物量较高。与2007年调查结果比较,该海域大型底栖动物的平均生物量明显下降,而平均栖息密度增加了近6倍。

表3 大型底栖动物种类组成的年际变化

表4 大型底栖动物平均生物量和栖息密度的变化
3.2棒锥螺的死亡现象
1981—1982年[11]在温州沿岸15~20 m等深线的海域调查发现大型底栖动物优势种是棒锥螺,且该种数量很大,生物量高达每平方米几千克或数百克;1990年春季[12]在洞头本岛附近海域调查发现大型底栖动物中软体动物占有生物量的优势,其动态变化主要受棒锥螺和黄短口螺的影响;2006年5月和2007年1月[13]调查发现瓯江口及其邻近海域大型底栖动物在站位S4和S10(分别对应于本次调查的站位S14和S25)物种分布单一,但数量集中,生物量主要受棒锥螺影响。本次调查发现站位S14和S25物种均为多毛类,且栖息密度和生物量均很低,瓯江口海域没有棒锥螺出现,其它站位出现的棒椎螺均为死壳,分别为洞头海域南部的S34和S35站位(棒锥螺的栖息密度均为10个/m2),与瓯江口北部沿岸海域S8站位(棒锥螺的栖息密度为70个/m2)。这说明调查海域曾经适宜棒锥螺生存,近年来变化的栖息环境不再适宜其生存。另外,由于棒锥螺是腐食性的腹足类动物,这一现象也暗示该海域曾经生活有大量的像棒锥螺一样的腐食性腹足类动物因栖息环境的改变而大量死亡。
浙江省台州市养殖泥螺也出现过相似的情况,由最初的零星死亡现象到死亡面积扩大,最后到个别滩涂颗粒无收[22]。根据养殖泥螺暴发性死亡原因可推测本次调查海域棒锥螺死亡的原因:一是环境污染,水体富营养化与有害赤潮频发导致棒锥螺等腹足动物出现死亡,仅2003—2005年间,调查海域就发生了6次赤潮[23]。二是海域中某些污染物含量增加,水环境中的有机污染物如多环芳烃、多氯有机物、有机磷农药等可使腹足类慢性中毒[24],推测可能由于近年来工农业发展导致调查海域某些污染物含量增加而引起棒锥螺死亡。三是底质变化,棒锥螺适宜生活在泥沙底质,围垦促淤可使底质发生巨大改变[25],近年来调查海域跨海大桥、围垦等工程活动使底质发生变化导致其不再适宜棒锥螺生存。
3.3棘皮动物的分布与环境因子的关系
棘皮动物具有呈斑块状分布的特点,在少数海域其生物量很高,而在广阔海域中其生物量往往很低,甚至成片海域常采不到棘皮动物[11]。本次调查仅在洞头东部和南部海域出现棘皮动物再次证实了其斑块状分布的特点。
影响棘皮动物分布的环境因子有很多,如溶解氧、温度和盐度等。将本次调查测定的10个环境因子分别与各站位棘皮动物的生物量进行相关性分析可知,溶解氧含量与其生物量的相关系数为0.482,差异极显著(P<0.01)。本次调查溶解氧含量在8.43 mg/L以上的站位才有可能出现棘皮动物,其中S4站位的溶解氧含量最高,为9.38 mg/L,该站位棘皮动物的生物量最大,为8.48 g/m2。由于受到台湾暖流影响,洞头东部和南部海域水温较高,暖水种数量较多[26]。洞头西部海域由于受到瓯江及浙江南部沿岸流系的影响,海水盐度很低,而棘皮动物对低盐海水几乎普遍难以忍受[24],导致该区域多数棘皮动物无法生存,但有少数种如滩栖阳遂足能生活在盐度很低的海域[27],如在S4站位其单个个体可达到1.66 g。另外,与洞头岛群间的海域相比,西部海域底质扰动也比较剧烈,环境条件不稳定。
4结论
2015年春季在洞头列岛及邻近海域共获得大型底栖动物122种,多毛类和甲壳动物是该海域大型底栖动物群落的主要类群。棘皮动物仅分布在洞头东部和南部海域,瓯江口附近海域没有采到。大型底栖动物平均生物量为3.35 g/m2,平均栖息密度为321个/m2。Shannon-Wiener多样性指数、Margalef丰富度指数和Pielou均匀度指数的平均值分别为1.29、2.45和0.66。瓯江口附近海域以及瓯江口与洞头列岛之间海域大型底栖动物的栖息密度和生物量较低,洞头列岛东南部海域大型底栖动物的栖息密度和生物量高于西北部海域。
洞头列岛及邻近海域大型底栖动物可划分为3个群落:位于乐清湾口正对面海域的异蚓虫-薄云母蛤-绒毛细足蟹-不倒翁虫群落(群落Ⅰ),位于洞头东部和南部海域的薄云母蛤-异蚓虫-双鳃内卷齿蚕-不倒翁虫群落(群落Ⅱ)以及位于瓯江口附近海域的其他群落(群落Ⅲ)。典范对应分析结果表明影响大型底栖动物群落的主要环境因子有底层海水的温度、盐度、活性磷酸盐、活性硅酸盐、溶解性无机氮、化学需氧量以及表层沉积物中的总有机碳等。ABC曲线分析结果表明,大型底栖动物群落受到了一定程度的扰动,但基本稳定。与历史资料相比,大型底栖动物的密度和生物量均无明显变化,种类组成由以多毛类和软体动物为主转变为以多毛类和甲壳动物为主,且个体较大的棒锥螺大量死亡。
参考文献(References):
[1] CAI Wen-qian, MENG Wei, LIU Lu-san, et al. Macrozoobenthos community structure of the Bohai Bay in spring time[J]. Acta Scientiae Circumstantiae,2013,33(5):1 458-1 466.
蔡文倩,孟伟,刘录三,等.春季渤海湾大型底栖动物群落结构特征研究[J].环境科学学报,2013,33(5):1 458-1 466.
[2] WU Bin, SONG Jing-ming, LI Xue-gang. Evaluation of potential relationships between benthic community structure and toxic metals in Laizhou Bay[J]. Marine Pollution Bulletin,2014(87):247-256.
[3] KALANTZI I, PAPAGEORGIOU N, SEVASTOU K. Metals in benthic macrofauna and biogeochemical factors affecting their trophic transfer to wild fish around fish farm cages[J]. Science of the Total Environment,2014(470-471):742-753.
[4] QIN Yan-wen, ZHENG Bing-hui, LEI Kun, et al. Distribution and mass inventory of polycyclic aronmatic hydrocarbons in the sediments of the south Bohai Sea, China[J]. Marine Pollution Bulletin,2011(62):371-376.
[5] MARIANO J A, PAULO DA C L, CLAUDIA B, et al. Macrobenthos and multi-molecular markers as indicators of environmental contamination in a South American port[J]. Marine Pollution Bulletin,2013(73):102-114.
[6] SHIN P K S, HUANG Z G, WU R S S. An updated baseline of subtropical macrobenthic communities in Hong Kong[J]. Marine Pollution Bulletin,2004(49):128-135.
[7] WANG Tie-gan, HU Ren-yong, SUN Qing-hai, et al. Marine macroalgae of Dongtou Islands in Zhejiang Province[M]. Beijing: China Ocean Press,2012.
王铁杆,胡仁勇,孙庆海,等.浙江洞头大型海藻[M].北京:海洋出版社,2012.
[8] QIU Jian-biao, PENG Xin, XIE Qi-lang, et al. The spatial-temporal distribution and its biodiversity of intertidal macrobenthos in Dongtou Archipelago[J]. Journal of Fisheries of China,2012,36(4):608-614.
仇建标,彭欣,谢起浪,等.洞头列岛潮间带大型底栖生物的时空分布及其多样性[J].水产学报,2012,36(4):608-614.
[9] WANG Yi-nong, ZHANG Yong-pu, YU Hong, et al. Study on mollusca and ecological characteristic in the sea shore of Dongtou Island, Zhejiang Province[J]. Journal of Ningbo University:NSEE,1999,12(3):39-48.
王一农,张永普,於宏,等.浙江洞头列岛的软体动物及生态特点[J].宁波大学学报:理工版,1999,12(3):39-48.
[10] ZHANG Yong-pu, ZHENG Jie, WANG Yi-nong. Ecological characteristics of the intertidal mussels of the islands south of Zhejiang[J]. Transactions of Oceanology and Limnology,2000(3):24-28.
张永普,郑洁,王一农.浙南岛屿岩相潮间带贻贝类的生态特点[J].海洋湖沼通报,2000(3):24-28.
[11] LIU Rui-yu. Biological resources of coastal zone in China[M]. Beijing: China Ocean Press,1996.
刘瑞玉.中国海岸带生物[M].北京:海洋出版社,1996.
[12] Research Group of Comprehensive Survey for Biological Resources in The Island Waters of Zhejiang. A report of comprehensive survey for biological resources in the island waters of Zhejiang[M]. Hangzhou: Second Institute of Oceanography,SOA,1993.
浙江海岛海域生物资源综合调查课题组.浙江省海岛海域生物资源综合调查报告[M].杭州:国家海洋局第二海洋研究所,1993.
[13] SHOU Lu, ZENG Jiang-ning, LIAO Yi-bo, et al. Seasonal distribution of macrozoobenthos and its relations to environmental factors in Oujiang River estuary sea area[J]. Chinese Journal of Applied Ecology,2009,20(8):1 958-1 964.
寿鹿,曾江宁,廖一波,等.瓯江口海域大型底栖动物分布及其与环境的关系[J].应用生态学报,2009,20(8):1 958-1 964.
[14] SHOU Lu, ZENG Jiang-ning, XUE Bin, et al. Coastal environment and marine environmental capacity of Zhejiang Province[M]. Beijing: China Ocean Press,2015.
寿鹿,曾江宁,薛斌,等.浙江沿岸生态环境及海湾环境容量[M].北京:海洋出版社,2015.
[15] GBT12763.6—2007 National oceanography census regulation methods: Marine biology survey[S].2007.
GBT12763.6—2007海洋调查规范:海洋生物调查[S].2007.
[16] GB 17378.4—2007 The specification for marine monitoring: Seawater analysis[S].2007.
GB 17378.4—2007 海洋监测规范: 海水分析[S].2007.
[17] SHEN Guo-ying, SHI Bing-zhang. Marine ecology[M]. Beijing: China Science Press,2002.
沈国英,施并章.海洋生态学[M].北京:科学出版社,2002.
[18] ZHOU Hong, ZHANG Zhi-nan. Rationale of the multivariate statistical software PRIMER and its application in benthic community ecology[J]. Journal of Ocean University of Qingdao,2003,33(1):58-64
周红,张志南.大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用[J].青岛海洋大学学报,2003,33(1):58-64.
[19] CLARKE K, WARWICK R. Change in marine communities : An approach to statistical analysis and interpretation[R]. 2nd ed. Plymouth: PRIMER-E,2001.
[20] SUN Ya-wei, CAO Lian, QIN Yu-tao, et al. Analysis of macrobenthos community structure in the adjacent sea area of Changjiang River Eastuary[J]. Marine Science Bulletin,2007,26(2):66-70.
孙亚伟,曹恋,秦玉涛,等.长江口邻近海域大型底栖生物群落结构分析[J].海洋通报,2007,26(2):66-70.
[21] REN Xian-qiu. Fauna Sinica: Crustacea[M].Beijing: China Science Press,2006.
任先秋.中国动物志:甲壳动物亚门[M].北京:科学出版社,2006.
[22] WANG Guo-liang, WANG Yi-nong, YU Hong, et al. Research on the cause and prevention of sudden death ofBullactaexarata[J]. China Fisheries,2003(1):51-52.
王国良,王一农,於宏,等.养殖泥螺暴发性死亡的原因分析及防治[J].中国水产,2003(1):51-52.
[23] YAO Wei-min, ZHENG Ai-rong, QIU Jin-kun. Eutrophication in Dongtou Islands Zhejiang and the relationships between eutrophication and red tide[J]. Marine Environmental Science,2007,26(5):466-469.
姚炜民,郑爱榕,邱进坤.浙江洞头列岛海域水体富营养化及其与赤潮的关系[J].海洋环境科学,2007,26(5):466-469.
[24] ZHANG Qing-shun, HOU Jian-jun, XIONG Bang-xi. Ecotoxicological effects of POPs and heavy metals on Gastropods[J]. Journal of Hydroecology,2008,28(4):1-4.
张清顺,侯建军,熊邦喜.持久性有机污染物和重金属对腹足类的生态毒理效应[J].水利渔业,2008,28(4):1-4.
[25] MA Chang-an, XU Lin-lin, TIAN Wei, et al. The influence of a reclamation project on the macrobenthos of an east Nanhui Tidal Flat[J]. Acta Ecologica Sinica,2012,32(4):1 007-1 015.
马长安,徐霖林,田伟,等.围垦对南汇东滩湿地大型底栖动物的影响[J].生态报,2012,32(4):1 007-1 015.
[26] LIAO Yu-lin, XIAO Ning. Species composition and faunal characteristics of echinoderms in China seas[J]. Biodiversity Science,2011,19(6):729-736.
廖玉麟,肖宁.中国海棘皮动物的种类组成及区系特点[J].生物多样性,2011,19(6):729-736.
[27] LIAO Yu-lin. Fauna Sinica: Echinodermata[M].Beijing: China Science Press,2004.
廖玉麟.中国动物志:棘皮动物门[M].北京:科学出版社,2004.
收稿日期:2015-12-23修回日期:2016-03-08
基金项目:国家海洋公益性行业科研专项项目资助(201305009,201305043,201405007,201505004-3);国家海洋局第二海洋研究所基本科研业务费专项项目资助(JG1311,JG1312)
作者简介:贾胜华(1990-),男,河南平顶山市人,主要从事海洋生态学及大型底栖动物方面的研究。E-mail:jiashenghuahh@163.com *通讯作者:曾江宁(1975-),男,研究员,主要从事海洋生态学方面的研究。E-mail:jiangningz@126.com;zjn@sio.org.cn
中图分类号:Q958.8
文献标识码:A
文章编号:1001-909X(2016)02-0083-10
Doi:10.3969/j.issn.1001-909X.2016.02.011
Research on macrozoobenthic community structure in Dongtou Islands and adjacent sea area
JIA Sheng-hua1,2, ZENG Jiang-ning*1,2, LIAO Yi-bo1,2, SHOU Lu1,2, HUANG Wei1,2,GAO Ai-gen1,2, TANG Yan-bin1,2
(1.KeylaboratoryofMarineEcosystemandBiogeochemistry,SOA,Hangzhou310012,China; 2.SecondInstituteofOceanography,SOA,Hangzhou310012,China)
Abstract:Based on the macrozoobenthos and environmental materials from 35 stations in Dongtou Islands and adjacent sea area investigated in April 2015, the characteristics of macrozoobenthic community structure and its relationship with environmental factors were analyzed. Results show that a total of 122 species are identified, including 57 species of Polytraeta, 34 species of Crustacea, 19 species of Mollusca, 5 species of Echinodermata and 7 others species. The number of species is about 1~22 at each station, with lower values of species number concentrated in Oujiang River Estuary. The average biomass of macrozoobenthos is 3.35 g/m2, and the average inhabited density is 321 ind/m2. Polytraeta has an advantage on the species compositions, while Mollusca has the more advantages on the biomass and abundance. The average of Shannon-Wiener index(H′) is 1.29, the average of Margalef’s species richness index (d) is 2.45, and the average of Pielou’s evenness index (J′) of species is 0.66. By species similarity cluster analysis and multidimensional scaling ordination analysis, three communities of macrozoobenthos can be divided in Dongtou Islands and adjacent sea area, those are: Heteromastus filiforms-Yoldia similis-Raphidopus ciliatus-Sternaspis scutata (Community Ⅰ), Yoldia similis-Heteromastus filiforms-Aglaophamus dibranchis-Sternaspis scutata (Community Ⅱ), and others (Community Ⅲ ). Canonical correspondence analysis show that the sea water temperature, water salinity, activated phosphate, activated silicate, dissolved inorganic nitrogen, chemical oxygen demand and the total organic carbon in surface sediment are the main environmental factors affecting the macrozoobenthic community. The ABC curves indicate that the macrozoobenthic communities in Dongtou Islands and adjacent sea area have suffered contamination or have disturbed to a certain degree. Compared with historical data, the average biomass and abundance of macrozoobenthos have not obviously changed. The dominant organism of the species compositions are Polytraeta and Crustacea, instead of Polytraeta and Mollusca, and the number of Turritella bacillum in the area has obviously declined.
Key words:macrozoobenthos; community structure; species diversity; environmental factor; Dongtou Islands and adjacent sea area
贾胜华,曾江宁,廖一波,等.洞头列岛及邻近海域大型底栖动物群落结构的研究[J].海洋学研究,2016,34(2):83-92,doi:10.3969/j.issn.1001-909X.2016.02.011.
JIA Sheng-hua, ZENG Jiang-ning, LIAO Yi-bo, et al. Research on macrozoobenthic community structure in Dongtou Islands and adjacent sea area[J]. Journal of Marine Sciences, 2016,34(2):83-92, doi:10.3969/j.issn.1001-909X.2016.02.011.
