秦巴山区黑木耳菌株酯酶同工酶的分析
2016-07-25钱雪婷陈文强彭浩解修超邓百万
钱雪婷++陈文强++彭浩++解修超++邓百万
摘要:对秦巴山区41个黑木耳供试菌株进行酯酶同工酶垂直板聚丙烯酰胺凝胶电泳分析,结果表明,41个供试菌株共检测到19条不同迁移率酯酶同工酶酶带,各菌株有5~9条不等的酶带,共16个酶谱类型。应用NTSYS-pc 2.11软件分析,41个黑木耳菌株两两之间的相似系数分布在0.474~1.000之间,当遗传相似系数为80%时,41个黑木耳供试菌株被聚类分成5类,初步鉴定了41个供试菌株间的亲缘关系。
关键词:秦巴山区;黑木耳;酯酶同工酶;亲缘关系
中图分类号: S646.603文献标志码: A文章编号:1002-1302(2016)06-0269-03
收稿日期:2015-09-23
基金项目:陕西省科学技术研究发展计划(编号:2014K01-16-03);陕西省科技统筹创新工程计划(编号:2012HBGC-20)。
作者简介:钱雪婷(1990—),女,陕西略阳人,硕士研究生,主要从事微生物资源利用开发等研究。E-mail:qianxueting163@163.com.
通信作者:陈文强,教授,硕士生导师,主要从事微生物资源开发保护及利用相关研究。E-mail:wenqiangc@126.com.黑木耳(Auricularia auricular),别称木耳、细耳、云耳、黑叶、木蛾,分类上属异担子菌纲、木耳目、木耳科、木耳属,子实体一般较小,胶质,浅圆盘形、耳形或不规则形[1],是一种含有极高营养价值和药用价值的美味食用菌[2]。现代研究表明,黑木耳具有抗氧化、抗菌、抗癌,增强体液免疫等多种功效[3-5]。秦巴山区气候温和湿润,适宜菌类生长,是我国黑木耳的主要栽培基地之一。近年来,由于部分地区对黑木耳菌株的管理存在很多缺陷,黑木耳菌株乱引种、乱命名的现象较为严重,使同一菌株冠以不同的品种代号或不同菌株冠以相同的品种代号,造成黑木耳菌种、品种的混乱。目前,黑木耳种质资源遗传多样性的研究逐渐受到重视,Tang等[6]、李媛媛等[7]、张旭等[8]、唐利华等[9]分别利用ISSR和SRAP分子标记、SCAR分子标记、SRAP和ITS分子标记、酯酶同工酶分析对我国不同地区黑木耳栽培菌株以及野生菌株的亲缘关系,结果表明,我国黑木耳菌株具有较高的遗传多样性。本研究利用酯酶同工酶技术对秦巴山区41个黑木耳菌株的亲缘关系进行分析,旨在为黑木耳菌种管理和种质资源遗传多样性的研究提供理论依据。
1材料与方法
1.1材料
1.1.1供试菌株秦巴山区41个黑木耳供试菌株及来源见表1。
1.1.2培养基母种培养基(CPDA):20%马铃薯,2%葡萄糖,0.5% KH2PO4,0.3% MgSO4·7H2O,0.001% 维生素B1,1.2%琼脂,pH值自然。同工酶制备液体培养基(元葱培养基):20%元葱,1%蔗糖,0.006%阿魏酸,pH值自然。
1.1.3主要仪器高压蒸汽灭菌锅(MJ-54A型,上海施都凯仪器设备有限公司);超净工作台(SW-CJ-2F型,苏州净化设备有限公司);电子分析天平(TB-214型,北京赛得利斯仪器系统有限公司);离心机(5418R型,德国Eppendorf);电泳仪(DYCZ-24B型,北京市六一仪器厂);凝胶成像系统(GEL-RAD型,美国伯乐)。
1.2方法
1.2.1样品制备将黑木耳菌株接种于CPDA斜面培养基,28 ℃培养,待菌丝长满管后,4 ℃冰箱保藏备用;将活化后的黑木耳菌株,接种到元葱培养基中,250 mL三角瓶装液量为100 mL,28 ℃静置培养20 d。将菌丝体过滤,蒸馏水冲洗,滤纸吸干水分,称质量,放入研钵中,加入液氮充分研磨,分装于离心管,8 000 r/min,4 ℃离心20 min,上清液为酶液。
1.2.2酯酶同工酶的检测采用垂直板聚丙烯酰胺凝胶电泳法[10],配制5%浓缩胶,10%分离胶,电极缓冲液为Tris-Glycine系统,pH值8.3。上样量:酶液6 μL,溴酚蓝指示剂2 μL,10%蔗糖5 μL。浓缩胶电压80 V,室温,电泳45 min,分离胶电压120 V,4 ℃,电泳4 h。取下凝胶,置于α、β-乙酸萘酯和坚牢蓝配制的染色液中,37 ℃,轻微振荡,染色 25 min,将出现红褐色酶带的凝胶置于7%醋酸溶液中脱色、保存,扫描照相。
1.2.3聚类分析相对迁移率Rf=X/Y,X为从点样孔下沿开始,酶带在凝胶中的迁移距离,Y为从点样孔下沿开始,指示剂在凝胶中的迁移距离。在测量迁移距离时,以酶带的中部位置为准。依据酯酶酶谱,按照条带的有无进行标记,在相同迁移率位置上,有条带的记为1,无条带的记为0。利用NTSYS-pc 2.11软件,分析得到秦巴山区41个黑木耳菌株的聚类分析树状图。
2结果与分析
2.1酯酶同工酶酶谱分析
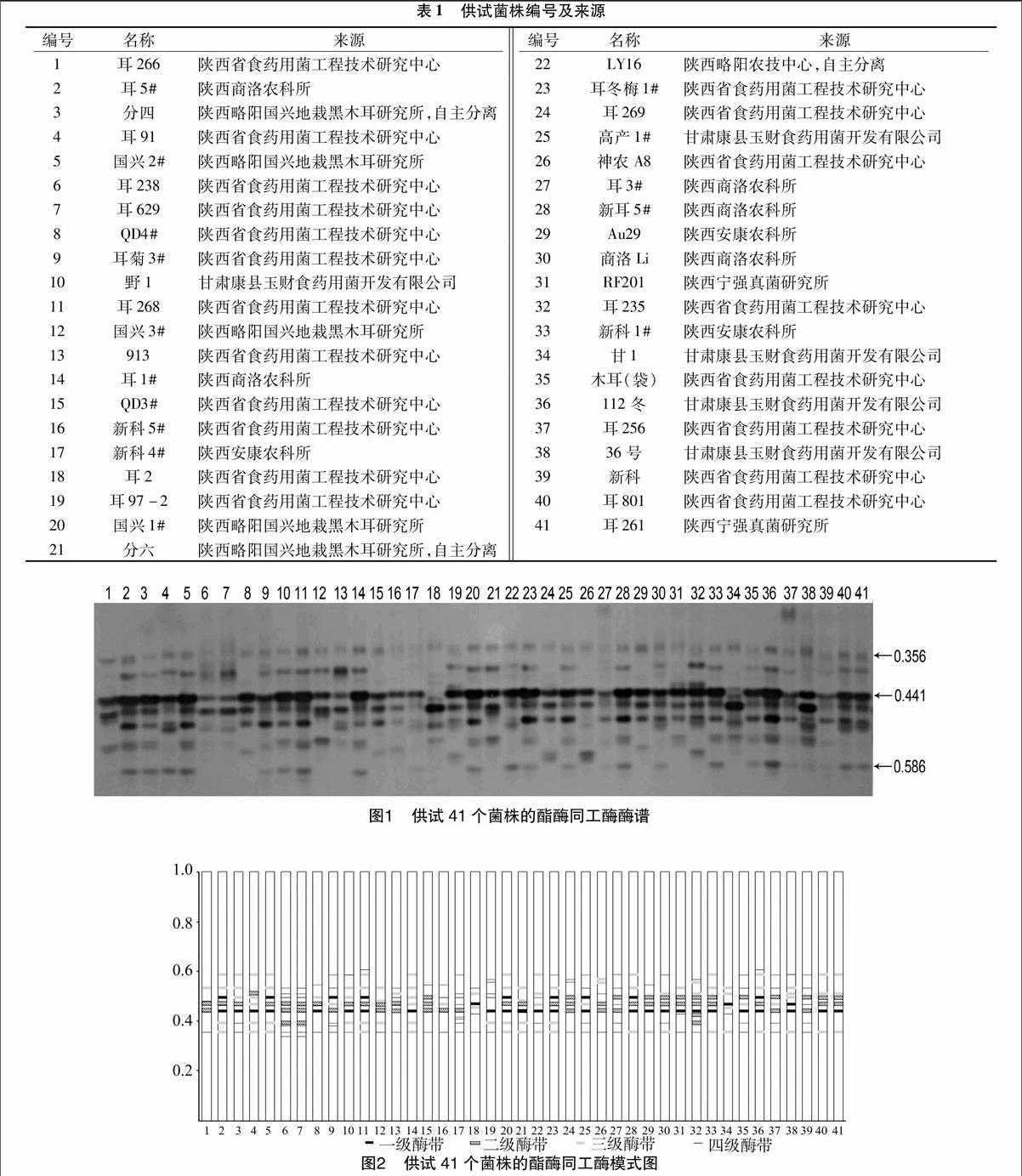
秦巴山区41个黑木耳供试菌株酯酶同工酶酶谱见图1。根据各菌株间酶带颜色和宽窄的差异[9],酯酶酶带可分为:一级酶带(色深而宽);二级酶带(色较深而宽);三级酶带(色较浅而窄);四级酶带(色很浅而窄),结合酯酶酶谱,绘制酯酶同工酶模式图(图2)。
图1显示,秦巴山区41个黑木耳供试菌株共检测到281条酯酶同工酶酶带,各菌株有5~9条不等的酶带,且多数为6~8条。41个供试菌株的迁移率分析表明,共有19条迁移率不同的酶带,迁移率都在0.344~0.606之间。41个供试菌株在Rf值为0.356和0.469这2处均有酶带出现,可能为黑木耳的共有酯酶酶带。41个菌株共得到16个酶谱类型,其中黑木耳36#、耳菊3#、耳266、耳913、耳2、耳269、神农A8、RF201以及甘1各为1个酶谱类型;耳5#、分四、国兴2#、野1#、耳1#、国兴1#、耳冬梅1#、高产1#、耳3#、新耳5#、新科1#、木耳(袋)、耳256以及新科的酶谱带型基本相同;耳91、新科4#、LY16#、商洛Li、耳801以及耳261的酶谱带型基本相同;QD3#、QD4#、新科5#以及Au29的酶谱带型基本相同;耳238与耳629的酶谱带型基本相同;耳268与112冬的酶谱带型基本相同;国兴3#与分六的酶谱带型基本相同;耳235与耳97-2的酶谱带型基本相同。
图2显示,在相同Rf值处,不同黑木耳菌株的酶带等级不同(酶带颜色深浅与宽窄不同)。且41个黑木耳菌株在Rf值为0.441处,一级酶带较多,表明黑木耳菌株该酯酶活性高且含量高;在Rf值为0.469与0.494这2处,二级酶带较多,表明这些酯酶活性较高且含量较高;在Rf值为0.356、0.396以及0.586等处,三级酶带与四级酶带较多,表明这些酯酶活性低且含量低。
2.2聚类分析
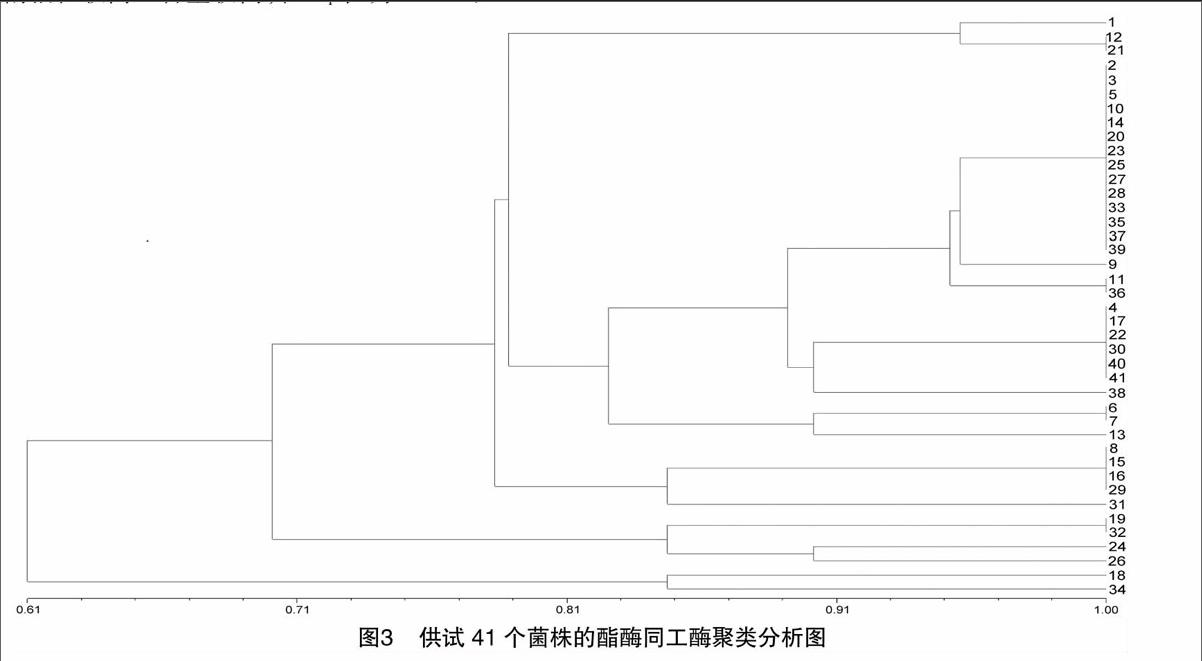
根据酯酶酶谱,应用NTSYS-pc 2.11软件,构建秦巴山区41个黑木耳供试菌株聚类分析树状图(图3)。
图3显示,当遗传相似系数为70%时,秦巴山区41个黑木耳供试菌株被聚类分成3类。当遗传相似系数为80%时,41个黑木耳供试菌株被聚类分成5类,第1类:耳266、国兴3#以及分六;第2类:耳5#、分四、国兴2#、野1#、耳1#、国兴1#、耳冬梅1#、高产1#、耳3#、新耳5#、新科1#、木耳(袋)、256、新科、耳菊3#、耳268、112冬、耳91#、新科4#、LY16#、商洛Li、耳801、耳261、36号、耳238、耳629以及耳913;第3类:QD4#、QD3#、新科5#、Au29以及RF201;第4类:耳97-2、耳235、耳269以及神农A8;第5类:耳2以及甘1。
3讨论
同工酶分子标记是从基因产物水平来研究生物的遗传变异,是一种重要的分子遗传标记技术[11],酶带的变化可作为鉴定物种,研究分类、进化、遗传和变异的重要指标[12],也是对食药用菌进行遗传分析或生化鉴定中的最适标记手段之一。自Markert等提出同工酶概念以来,酯酶同工酶标记技术在食用菌研究中逐步得到广泛的应用[13-15]。
本研究将秦巴山区41个黑木耳供试菌株在严格统一的条件下,如相同的培养温度、时间,相同的酶液制备方法等,采用5%浓缩胶与10%分离胶进行垂直板聚丙烯酰胺凝胶电泳,共检测到19条不同迁移率的酯酶酶带,16个酶谱类型,当遗传相似系数为80%时,聚类分成5类,如耳97-2、耳235、耳269以及神农A8被聚类在一个分支上,表明这4个菌株亲缘关系较近。彭浩等对陕南10种黑木耳主要栽培种进行酯酶同工酶的分析,检测到8条不同迁移率的酯酶酶带,当遗传相似系数为80%时,聚类分成5类[16]。与之相比,本研究选用的供试菌种地域和菌株数更为广泛,且检测出的酯酶酶带较丰富,聚类结果更加精细。
通过酯酶同工酶分析,得到秦巴山区41个黑木耳供试菌株聚类分析树状图,初步鉴定了不同菌株之间的亲缘关系。结果表明,秦巴山区黑木耳菌株的酯酶多态性高,种质资源较为丰富,为秦巴山区黑木耳优良菌种的选育以及菌种的管理工作提供理论依据。由于酯酶活性很容易受到外界因素的影响,酯酶同工酶的稳定性以及试验的可重复性不高,对秦巴山区41个黑木耳供试菌株进行RAPD和酯酶同工酶指纹图谱的分析还有待进一步研究。
参考文献:
[1]卯晓岚. 中国蕈菌[M]. 北京:科学出版社,2009:647-652.
[2]陈文强,王进,邓百万,等. 秦巴山区黑木耳主要栽培种内原生质体分离与融合[J]. 食品与生物技术学报,2014,33(8):856-864.
[3]Peng X B,Li Q,Ou L N,et al. GC-MS,FT-IR analysis of black fungus polysaccharides and its inhibition against skin aging in mice[J]. International Journal of Biological Macromolecules,2010,47(2):304-307.
[4]Nguyen T L,Chen J,Hu Y L,et al. In vitro antiviral activity of sulfated Auricularia auricula polysaccharides[J]. Carbohydrate Polymers,2012,90(3):1254-1258.
[5]Nguyen T L,Wang D Y,Hu Y L,et al. Immuno-enhancing activity of sulfated Auricularia auricula polysaccharides[J]. Carbohydrate Polymers,2012,89(4):1117-1122.
[6]Tang L H,Xiao Y,Li L,et al. Analysis of genetic diversity among Chinese Auricularia auricula cultivars using combined ISSR and SRAP markers[J]. Current Microbiology,2010,61(2):132-140.
[7]李媛媛,隋玉龙,牛淑力,等. SCAR标记在黑木耳栽培菌株分类鉴定中的应用[J]. 菌物研究,2013,11(3):182-185,189.
[8]张旭,李倩,刘华晶,等. SRAP和ITS分子标记在大兴安岭地区野生黑木耳遗传多样性上的应用[J]. 黑龙江科学,2013,4(4):17-21.
[9]唐利华,郭倩,王瑞娟,等. 中国黑木耳主要栽培菌株酯酶同工酶的研究[J]. 食用菌学报,2007,14(4):37-40.
[10]胡能书,万贤国.同工酶技术及其应用[M]. 长沙:湖南科学技术出版社,1985:59-76.
[11]郑素月,张海花. 同工酶技术及其在食用菌种质资源研究中的应用与进展[J]. 邯郸农业高等专科学校学报,2004,21(3):16-18.
[12]张欣倪,王鸿磊,丁强,等. 国外引进双孢菇菌种与国内栽培菌种的 RAPD和酯酶同工酶指纹图谱分析[J]. 江苏农业科学,2015,43(7):254-256.
[13]Bian Y B,Wu K Y,Wu C S. Genetic analysis of isozyme loci of intraspecific hybrid in Auricularia auricula[J]. Acta Genetica Sinica,2003,30(1):76-80.
[14]韦仕岩,王灿琴,覃晓娟,等. 金福菇菌株的酯酶同工酶分析[J]. 基因组学与应用生物学,2014,33(5):1025-1030.
[15]隋玉龙,宋慧,杜金,等. 灵芝栽培菌株酯酶同工酶的酶谱多样性[J]. 菌物研究,2013,11(2):136-140.
[16]彭浩,陈文强,邓百万,等. 陕南10株黑木耳主要栽培种酯酶同工酶的研究[J]. 江苏农业科学,2014,42(9):193-196.
钱雪婷 陈文强 彭浩 解修超 邓百万


摘要:对秦巴山区41个黑木耳供试菌株进行酯酶同工酶垂直板聚丙烯酰胺凝胶电泳分析,结果表明,41个供试菌株共检测到19条不同迁移率酯酶同工酶酶带,各菌株有5~9条不等的酶带,共16个酶谱类型。应用NTSYS-pc 2.11软件分析,41个黑木耳菌株两两之间的相似系数分布在0.474~1.000之间,当遗传相似系数为80%时,41个黑木耳供试菌株被聚类分成5类,初步鉴定了41个供试菌株间的亲缘关系。
关键词:秦巴山区;黑木耳;酯酶同工酶;亲缘关系
中图分类号: S646.603文献标志码: A文章编号:1002-1302(2016)06-0269-03
收稿日期:2015-09-23
基金项目:陕西省科学技术研究发展计划(编号:2014K01-16-03);陕西省科技统筹创新工程计划(编号:2012HBGC-20)。
作者简介:钱雪婷(1990—),女,陕西略阳人,硕士研究生,主要从事微生物资源利用开发等研究。E-mail:qianxueting163@163.com.
通信作者:陈文强,教授,硕士生导师,主要从事微生物资源开发保护及利用相关研究。E-mail:wenqiangc@126.com.黑木耳(Auricularia auricular),别称木耳、细耳、云耳、黑叶、木蛾,分类上属异担子菌纲、木耳目、木耳科、木耳属,子实体一般较小,胶质,浅圆盘形、耳形或不规则形[1],是一种含有极高营养价值和药用价值的美味食用菌[2]。现代研究表明,黑木耳具有抗氧化、抗菌、抗癌,增强体液免疫等多种功效[3-5]。秦巴山区气候温和湿润,适宜菌类生长,是我国黑木耳的主要栽培基地之一。近年来,由于部分地区对黑木耳菌株的管理存在很多缺陷,黑木耳菌株乱引种、乱命名的现象较为严重,使同一菌株冠以不同的品种代号或不同菌株冠以相同的品种代号,造成黑木耳菌种、品种的混乱。目前,黑木耳种质资源遗传多样性的研究逐渐受到重视,Tang等[6]、李媛媛等[7]、张旭等[8]、唐利华等[9]分别利用ISSR和SRAP分子标记、SCAR分子标记、SRAP和ITS分子标记、酯酶同工酶分析对我国不同地区黑木耳栽培菌株以及野生菌株的亲缘关系,结果表明,我国黑木耳菌株具有较高的遗传多样性。本研究利用酯酶同工酶技术对秦巴山区41个黑木耳菌株的亲缘关系进行分析,旨在为黑木耳菌种管理和种质资源遗传多样性的研究提供理论依据。
1材料与方法
1.1材料
1.1.1供试菌株秦巴山区41个黑木耳供试菌株及来源见表1。
1.1.2培养基母种培养基(CPDA):20%马铃薯,2%葡萄糖,0.5% KH2PO4,0.3% MgSO4·7H2O,0.001% 维生素B1,1.2%琼脂,pH值自然。同工酶制备液体培养基(元葱培养基):20%元葱,1%蔗糖,0.006%阿魏酸,pH值自然。
1.1.3主要仪器高压蒸汽灭菌锅(MJ-54A型,上海施都凯仪器设备有限公司);超净工作台(SW-CJ-2F型,苏州净化设备有限公司);电子分析天平(TB-214型,北京赛得利斯仪器系统有限公司);离心机(5418R型,德国Eppendorf);电泳仪(DYCZ-24B型,北京市六一仪器厂);凝胶成像系统(GEL-RAD型,美国伯乐)。
1.2方法
1.2.1样品制备将黑木耳菌株接种于CPDA斜面培养基,28 ℃培养,待菌丝长满管后,4 ℃冰箱保藏备用;将活化后的黑木耳菌株,接种到元葱培养基中,250 mL三角瓶装液量为100 mL,28 ℃静置培养20 d。将菌丝体过滤,蒸馏水冲洗,滤纸吸干水分,称质量,放入研钵中,加入液氮充分研磨,分装于离心管,8 000 r/min,4 ℃离心20 min,上清液为酶液。
1.2.2酯酶同工酶的检测采用垂直板聚丙烯酰胺凝胶电泳法[10],配制5%浓缩胶,10%分离胶,电极缓冲液为Tris-Glycine系统,pH值8.3。上样量:酶液6 μL,溴酚蓝指示剂2 μL,10%蔗糖5 μL。浓缩胶电压80 V,室温,电泳45 min,分离胶电压120 V,4 ℃,电泳4 h。取下凝胶,置于α、β-乙酸萘酯和坚牢蓝配制的染色液中,37 ℃,轻微振荡,染色 25 min,将出现红褐色酶带的凝胶置于7%醋酸溶液中脱色、保存,扫描照相。
1.2.3聚类分析相对迁移率Rf=X/Y,X为从点样孔下沿开始,酶带在凝胶中的迁移距离,Y为从点样孔下沿开始,指示剂在凝胶中的迁移距离。在测量迁移距离时,以酶带的中部位置为准。依据酯酶酶谱,按照条带的有无进行标记,在相同迁移率位置上,有条带的记为1,无条带的记为0。利用NTSYS-pc 2.11软件,分析得到秦巴山区41个黑木耳菌株的聚类分析树状图。
2结果与分析
2.1酯酶同工酶酶谱分析
秦巴山区41个黑木耳供试菌株酯酶同工酶酶谱见图1。根据各菌株间酶带颜色和宽窄的差异[9],酯酶酶带可分为:一级酶带(色深而宽);二级酶带(色较深而宽);三级酶带(色较浅而窄);四级酶带(色很浅而窄),结合酯酶酶谱,绘制酯酶同工酶模式图(图2)。
图1显示,秦巴山区41个黑木耳供试菌株共检测到281条酯酶同工酶酶带,各菌株有5~9条不等的酶带,且多数为6~8条。41个供试菌株的迁移率分析表明,共有19条迁移率不同的酶带,迁移率都在0.344~0.606之间。41个供试菌株在Rf值为0.356和0.469这2处均有酶带出现,可能为黑木耳的共有酯酶酶带。41个菌株共得到16个酶谱类型,其中黑木耳36#、耳菊3#、耳266、耳913、耳2、耳269、神农A8、RF201以及甘1各为1个酶谱类型;耳5#、分四、国兴2#、野1#、耳1#、国兴1#、耳冬梅1#、高产1#、耳3#、新耳5#、新科1#、木耳(袋)、耳256以及新科的酶谱带型基本相同;耳91、新科4#、LY16#、商洛Li、耳801以及耳261的酶谱带型基本相同;QD3#、QD4#、新科5#以及Au29的酶谱带型基本相同;耳238与耳629的酶谱带型基本相同;耳268与112冬的酶谱带型基本相同;国兴3#与分六的酶谱带型基本相同;耳235与耳97-2的酶谱带型基本相同。
图2显示,在相同Rf值处,不同黑木耳菌株的酶带等级不同(酶带颜色深浅与宽窄不同)。且41个黑木耳菌株在Rf值为0.441处,一级酶带较多,表明黑木耳菌株该酯酶活性高且含量高;在Rf值为0.469与0.494这2处,二级酶带较多,表明这些酯酶活性较高且含量较高;在Rf值为0.356、0.396以及0.586等处,三级酶带与四级酶带较多,表明这些酯酶活性低且含量低。
2.2聚类分析
根据酯酶酶谱,应用NTSYS-pc 2.11软件,构建秦巴山区41个黑木耳供试菌株聚类分析树状图(图3)。
图3显示,当遗传相似系数为70%时,秦巴山区41个黑木耳供试菌株被聚类分成3类。当遗传相似系数为80%时,41个黑木耳供试菌株被聚类分成5类,第1类:耳266、国兴3#以及分六;第2类:耳5#、分四、国兴2#、野1#、耳1#、国兴1#、耳冬梅1#、高产1#、耳3#、新耳5#、新科1#、木耳(袋)、256、新科、耳菊3#、耳268、112冬、耳91#、新科4#、LY16#、商洛Li、耳801、耳261、36号、耳238、耳629以及耳913;第3类:QD4#、QD3#、新科5#、Au29以及RF201;第4类:耳97-2、耳235、耳269以及神农A8;第5类:耳2以及甘1。
3讨论
同工酶分子标记是从基因产物水平来研究生物的遗传变异,是一种重要的分子遗传标记技术[11],酶带的变化可作为鉴定物种,研究分类、进化、遗传和变异的重要指标[12],也是对食药用菌进行遗传分析或生化鉴定中的最适标记手段之一。自Markert等提出同工酶概念以来,酯酶同工酶标记技术在食用菌研究中逐步得到广泛的应用[13-15]。
本研究将秦巴山区41个黑木耳供试菌株在严格统一的条件下,如相同的培养温度、时间,相同的酶液制备方法等,采用5%浓缩胶与10%分离胶进行垂直板聚丙烯酰胺凝胶电泳,共检测到19条不同迁移率的酯酶酶带,16个酶谱类型,当遗传相似系数为80%时,聚类分成5类,如耳97-2、耳235、耳269以及神农A8被聚类在一个分支上,表明这4个菌株亲缘关系较近。彭浩等对陕南10种黑木耳主要栽培种进行酯酶同工酶的分析,检测到8条不同迁移率的酯酶酶带,当遗传相似系数为80%时,聚类分成5类[16]。与之相比,本研究选用的供试菌种地域和菌株数更为广泛,且检测出的酯酶酶带较丰富,聚类结果更加精细。
通过酯酶同工酶分析,得到秦巴山区41个黑木耳供试菌株聚类分析树状图,初步鉴定了不同菌株之间的亲缘关系。结果表明,秦巴山区黑木耳菌株的酯酶多态性高,种质资源较为丰富,为秦巴山区黑木耳优良菌种的选育以及菌种的管理工作提供理论依据。由于酯酶活性很容易受到外界因素的影响,酯酶同工酶的稳定性以及试验的可重复性不高,对秦巴山区41个黑木耳供试菌株进行RAPD和酯酶同工酶指纹图谱的分析还有待进一步研究。
参考文献:
[1]卯晓岚. 中国蕈菌[M]. 北京:科学出版社,2009:647-652.
[2]陈文强,王进,邓百万,等. 秦巴山区黑木耳主要栽培种内原生质体分离与融合[J]. 食品与生物技术学报,2014,33(8):856-864.
[3]Peng X B,Li Q,Ou L N,et al. GC-MS,FT-IR analysis of black fungus polysaccharides and its inhibition against skin aging in mice[J]. International Journal of Biological Macromolecules,2010,47(2):304-307.
[4]Nguyen T L,Chen J,Hu Y L,et al. In vitro antiviral activity of sulfated Auricularia auricula polysaccharides[J]. Carbohydrate Polymers,2012,90(3):1254-1258.
[5]Nguyen T L,Wang D Y,Hu Y L,et al. Immuno-enhancing activity of sulfated Auricularia auricula polysaccharides[J]. Carbohydrate Polymers,2012,89(4):1117-1122.
[6]Tang L H,Xiao Y,Li L,et al. Analysis of genetic diversity among Chinese Auricularia auricula cultivars using combined ISSR and SRAP markers[J]. Current Microbiology,2010,61(2):132-140.
[7]李媛媛,隋玉龙,牛淑力,等. SCAR标记在黑木耳栽培菌株分类鉴定中的应用[J]. 菌物研究,2013,11(3):182-185,189.
[8]张旭,李倩,刘华晶,等. SRAP和ITS分子标记在大兴安岭地区野生黑木耳遗传多样性上的应用[J]. 黑龙江科学,2013,4(4):17-21.
[9]唐利华,郭倩,王瑞娟,等. 中国黑木耳主要栽培菌株酯酶同工酶的研究[J]. 食用菌学报,2007,14(4):37-40.
[10]胡能书,万贤国.同工酶技术及其应用[M]. 长沙:湖南科学技术出版社,1985:59-76.
[11]郑素月,张海花. 同工酶技术及其在食用菌种质资源研究中的应用与进展[J]. 邯郸农业高等专科学校学报,2004,21(3):16-18.
[12]张欣倪,王鸿磊,丁强,等. 国外引进双孢菇菌种与国内栽培菌种的 RAPD和酯酶同工酶指纹图谱分析[J]. 江苏农业科学,2015,43(7):254-256.
[13]Bian Y B,Wu K Y,Wu C S. Genetic analysis of isozyme loci of intraspecific hybrid in Auricularia auricula[J]. Acta Genetica Sinica,2003,30(1):76-80.
[14]韦仕岩,王灿琴,覃晓娟,等. 金福菇菌株的酯酶同工酶分析[J]. 基因组学与应用生物学,2014,33(5):1025-1030.
[15]隋玉龙,宋慧,杜金,等. 灵芝栽培菌株酯酶同工酶的酶谱多样性[J]. 菌物研究,2013,11(2):136-140.
[16]彭浩,陈文强,邓百万,等. 陕南10株黑木耳主要栽培种酯酶同工酶的研究[J]. 江苏农业科学,2014,42(9):193-196.
