基于cpDNA trnL-F序列的胡黄连保护遗传学研究
2016-07-25李国栋尹子丽刘小莉
李国栋, 尹子丽, 刘小莉*
( 1. 云南中医学院 中药学院, 昆明 650500; 2. 云南中医学院 民族医药学院, 昆明 650500 )
基于cpDNA trnL-F序列的胡黄连保护遗传学研究
李国栋1, 尹子丽2, 刘小莉1*
( 1. 云南中医学院 中药学院, 昆明 650500; 2. 云南中医学院 民族医药学院, 昆明 650500 )
摘要:胡黄连为特产中国-喜马拉雅特有高山植物,作为常用中、藏药材,受到灭绝性采挖,作为濒危和二级保护植物亟待科学的保护。该研究以云南和西藏7个野生居群91个个体为材料,基于cpDNA trnL-F非编码序列测序分析胡黄连的遗传多样性和遗传结构,分析显著进化单元,确立优先保护居群并提出科学的保护策略。结果表明:胡黄连trnL-F序列长度为871~876 bp,根据序列的核苷酸变异共鉴定出5个单倍型,西藏占有2个单倍型,云南占有3个单倍型,西藏和云南2个地区的所有单倍型均不共享。胡黄连具有较低的单倍型多样性(Hd = 0.434 19)和核苷酸多样性(Dij = 0.004 66)。种群间分化度(Fst=0.864 520)和基因流(Nm=0.04)、居群间的遗传分化水平(GST=0.916)、AMOVA分析(0.78%的遗传变异发生在居群内,60.97%的遗传变异发生在地区内居群间,38.25%的遗传变异发生在地区间)均表明,胡黄连居群间存在明显遗传分化。多数一致性树将胡黄连划分为3个进化分支(Ⅰ、Ⅱ、Ⅲ),这3个分支均与地理相关,分支Ⅰ分布于横断山区的4个居群,分支Ⅱ是分布于东喜马拉雅的一个居群,分支Ⅲ是分布于喜马拉雅中段的2个居群。3个分支分属于3个“进化显著单元(ESU)”。这3个ESU中白马雪山、茨中、定日、波密、聂拉木五个居群都需要保护,建议现阶段应优先保护的居群是云南白马雪山和西藏波密居群,以就地保护为主。
关键词:胡黄连, 濒危, trn L-F, 单倍型, 保护遗传学
胡黄连(Neopicrorhizascrophulariiflora)为玄参科的单种属植物,为中国-喜马拉雅特有高山植物,分布于我国云南西北部、西藏,海拔3 600~4 200 m的高山冷凉生境下。胡黄连具有重要的药用价值,是常用的中、藏药材,根状茎具有清湿热,除骨蒸、消疳热的功效(中华人民共和国药典,2010)。经过课题组全面调查发现,胡黄连资源蕴藏量急剧下降,生存受到严重威胁。已被收载在《中国珍稀濒危植物名录》《国家重点保护野生药材物种名录》中,目前对胡黄连研究主要集中在资源调查(杨少华等,2009)、栽培(杨少华等,2008)、化学(黄开毅等,2008)、药理(高宏伟等,2011)几个方面,对胡黄连遗传变异研究仅限于课题组前期基于ISSR分析(Liu et al, 2011)。近年来,叶绿体DNA (cpDNA)非编码区核苷酸序列变异已被经常用于分析植物的种群遗传变异(刘阳等,2010; 苏英娟等,2004),cpDNA测序法可以避免其它基于PCR的分子标记法经常遇到的长度同塑性问题,在估算种群遗传结构和基因流的能力方面具有较大优势(Chiang et al, 2001; 苏应娟等,2004)。本研究选择用于胡黄连近缘种(Picrorhizakurrooa)并且具有很好的位点变异的trnL-F片段对胡黄连开展了保护遗传学研究,旨在检测胡黄连基于cpDNA遗传多样性水平、遗传结构并确定优先保护种群,分析位于云南和西藏居群在地区水平上是否存在显著分化,划分显著进化单元,提出保护建议,其结果可以为胡黄连这一重要药用资源的可持续利用奠定基础。
1材料与方法
1.1 研究材料
本研究实验材料为采集自云南和西藏7个居群共91个个体,基本覆盖了该物种的所有已知分布点。每个居群的个体之间的距离至少5 m以上, 避免采集到同一株的克隆。选择幼嫩、干净的叶片,放到硅胶中干燥保存。
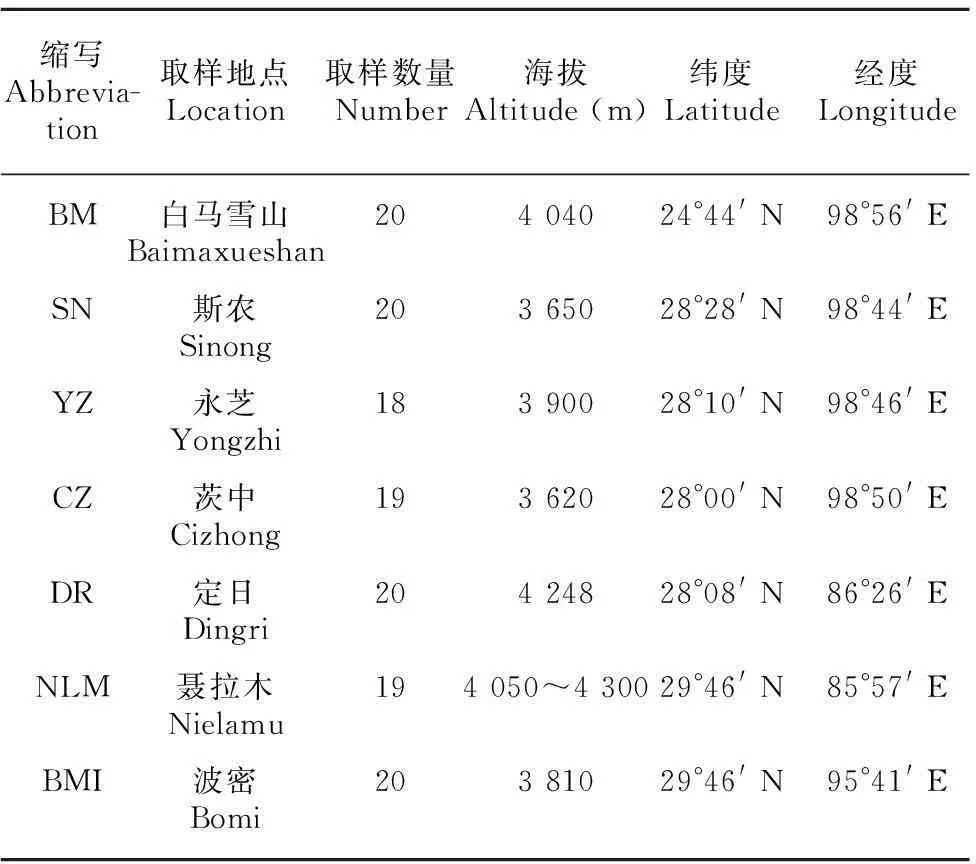
表 1 胡黄连7个居群的地理分布信息
研究材料采集点详细信息见表1,凭证标本存放于云南中医学院。
1.2 DNA提取
参考Doyle(1991) CTAB法,针对胡黄连叶片在研磨过程中极易褐化的问题, 在研磨时加入适量PVP粉,在65 ℃温浴过程中,将2×CTAB溶液与2 μL β-巯基乙醇混合,以有效解决褐化问题。
1.3 PCR扩增
引物序列为trnL: 5′-CGAAATCGGTAGACGCTACG-3′;trnF: 5′-ATTTGAACTGGTGACACGAG-3′,引物由上海生工技术有限公司合成。PCR扩增程序: 94 ℃预变性3 min;94 ℃30 s, 55 ℃ 30 s, 72 ℃ 60 s, 30个循环;末次循环72 ℃延伸7 min, 4 ℃保存。 PCR产物经2%的琼脂糖凝胶检测,扩增成功的样品送中美泰和生物技术(北京)有限公司测序(所有个体均进行双向测序)。
1.4 数据处理与分析
将每条序列与chromas 峰图逐一对比,结合Clustal X 1.83软件对序列进行排列校正。用BioEdit软件统计序列信息和核苷酸组成变异。用DnaSP 4.0软件分析统计单倍型数目、单倍型频率、单倍型多样度(遗传多样度,h)、核苷酸多样性(Dij)、基因流(Nm)等指标。
运用HAPLONST程序计算胡黄连总遗传多样性(HT)和居群内平均遗传多样性 (Hs)以及7个居群间遗传分化系数(GST)。GST和NST的比较采用U统计方法进行。在揭示胡黄连遗传变异的分布以及分化程度时,用Arlequin软件3.01中的分子方差分析AMOVA(Analysis of Molecular Variance)软件计算遗传变异在居群内、居群间及云南和西藏2个地区间的组成和单倍型分布的FST评价。应用PAUP*4.0 b10软件中的最大简约性分析法(maximum parsimony, MP)对单倍型进行系统发育分析, 选择Picrorchizakurroa、Wulfensiopsisamherst、短筒兔儿草(Lagotiskongboensis)、Plantagocoronopus作为外类群, 把空位作为缺失,采用启发式方式搜索,得到一致性系统树。分支的可靠性进行Bootstrap分析, 用1 000次的重复检验各个分支的支持率。
2实验结果
2.1 胡黄连trnL-F序列变异分析
对胡黄连7个居群 91个个体进行了cpDNA片段trnL-F序列的双向测定,序列长度在 871~876 bp之间,排序后长869 bp。共检测到5种单倍型(Hap1~Hap5)。单倍型序列比对后共发现13处变异位点:12处碱基替换和 1处插入或缺失,其中12处替换包括2处碱基转换,10处碱基颠换(表2)。通过统计91个个体的序列发现,碱基A/T含量丰富,占整个序列的比例为64.93%~65.1%,这与大多数植物叶绿体DNA碱基组成成分相符(苏应娟等,2004;陈生云等,2008)
2.2 胡黄连单倍型分布、单倍型多样性、核苷酸多样性
胡黄连每个居群单倍型数目、组成、频率、多样性以及核苷酸多样性指数见表3。胡黄连共有5个单倍型,分别是H1、H2、H3、H4、H5,单倍型H1、H3、H4、H5占绝对优势,单倍型H4的频率最高,在DR居群和NLM2个居群中出现,有27个个体,占总个体数的30%, 而单倍型H2的分布频率最低,仅在居群BM中有2个个体。H3在CZ居群、SN居群和YZ居群中出现。西藏3个居群共有2个单倍型即H4、H5,云南4个居群共有3个单倍型即H1、H2、H3,西藏和云南2个地区的所有单倍型均不共享,均为各自独特的单倍型,这也为胡黄连药材的产地鉴定提供了可能。由单倍型地理分布可见,胡黄连在云南和西藏2个地区间存在一定程度的隔离。

表 2 胡黄连叶绿体DNA trnL-F片段5个单倍型间的序列变异位点
▲=TCAAA

表 3 胡黄连7个居群叶绿体DNA单倍型的遗传多样度、组成和频率
胡黄连具有较低的单倍型多样性(Hd= 0.434 19)和核苷酸多样性(Dij= 0.004 66)。各个居群中,只有BM居群的单倍型多样性较高(Hd= 0.327 27), 其余居群的单倍型多样性均较低,其中SN和YZ居群均为同源组成。地区水平上,云南的单倍型多样性(Hd= 0.543 96)高于西藏(Hd= 0.000 0)。
2.3 遗传多样性和遗传结构
胡黄连7个居群cpDNAtrnL-F多样性在居群间的分化程度,两种中性检验法检验的结果均为正值(Fu and Li′s D*=1.470 34和Tajima′s=1.896 02),并且均呈显著水平(0.10>P>0.05)。同时,对胡黄连居群trnL-F片段序列数据的歧点分布分析显示观测值和期望值两者比较得到的是一个非单峰分布图(图1),此图明显偏离了群体扩张模型,结果表明胡黄连可能未曾经历过居群扩张。
根据胡黄连cpDNAtrnL-F序列变异(gap as the fifth state)采用Nei(1973)的算法估算出的居群间分化度(Fst)为0.864 52,基因流(Nm)为0.04,表明胡黄连各个居群间存的分化较大。在云南和西藏2个地区内,云南居群间的Fst为0~0.904 38,Nm为0.08,西藏居群间的Fst为0~1.000,Nm为0,说明2个地区内居群间的基因交流也近乎为零。
胡黄连总的遗传多样性HT(se)、居群内平均遗传多样性HS(se)、7个居群间遗传分化GST(se)和NST(se)值分别为0.861 (0.0443)、0.073 (0.0496)、0.916 (0.057 5)和0.987 (0.013 3)。使用U统计法检验胡黄连所有单倍型变异的地理结构,结果发现NST>GST(P<0.01),以上分析结果均表明胡黄连在整个中国-东喜马拉雅分布区内,居群间的遗传分化水平非常高(GST=0.916),单倍型的亲缘关系越相近越倾向于分布于同一居群中,并且有着明显的亲缘地理结构存在。将胡黄连按照地区分为云南和西藏2个组后, AMOVA分析也表明,胡黄连居群只有0.78%的遗传变异发生在居群内,而60.97%遗传变异发生在地区内居群间,38.25%遗传变异发生在地区间(表4), 这也揭示了胡黄连的遗传变异主要存在于居群间,而且具有相当高的居群分化水平。
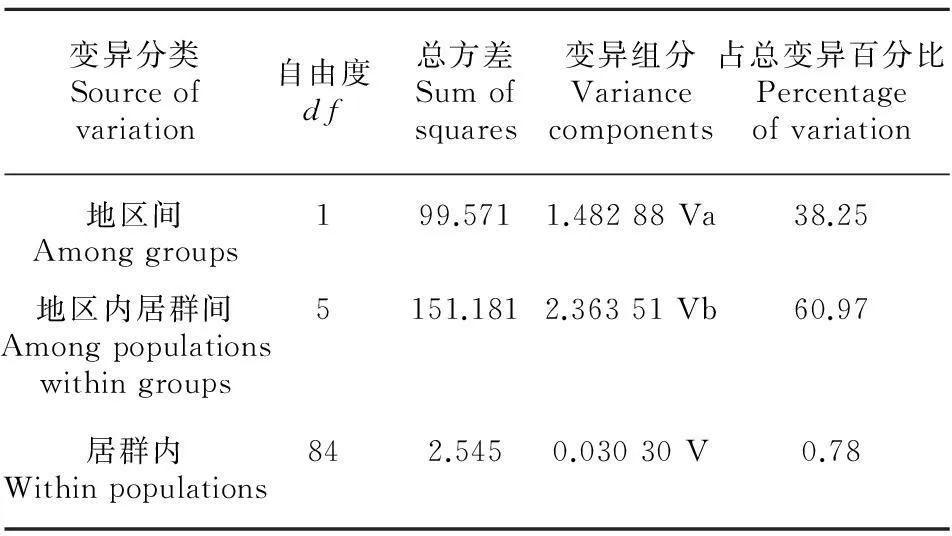
表 4 胡黄连2个地区之间的cpDNA AMOVA分析
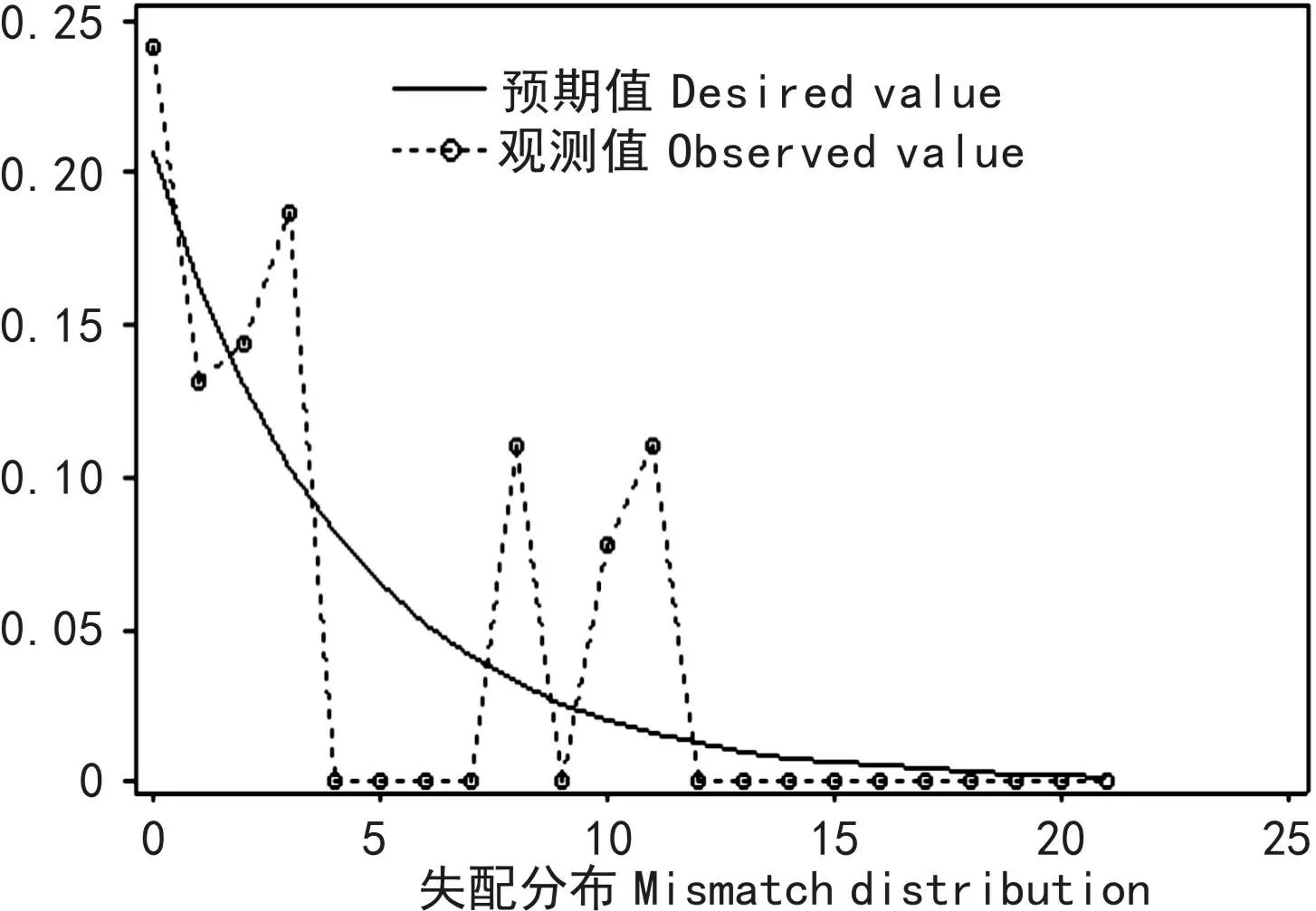
图 1 胡黄连cpDNA序列的失配分布图实线Exp-预期值;虚线Obs-观测值。Fig. 1 Mismatch distribution for cpDNA sequence data of N. scrophulariiflora Solid line represents the expected value; dotted line represents the observed value.
2.4 “显著进化单元”的划分
用粗筒兔儿草等做外类群对5个单倍型构建系统发育多数一致性树,通过启发式搜索得到一棵多数一致性树,得到了3个进化分支(GroupⅠ-Ⅲ),如图2。组Ⅰ和组Ⅱ共同构成一个单系,具有54%的支持率。组Ⅰ的单倍型为分布在横断山区的4个居群(BM、YZ、SN、CZ),构成一个单系,具有90%的支持率,组Ⅱ的单倍型分布在东喜马拉雅北端的2个居群(DR、NLM),组Ⅲ的单倍型分布在东喜马拉雅的居群(BMI),与P.kurrooa聚为一支,具有78%的支持率。因此GroupⅠ、GroupⅡ、GroupⅢ各自划分为一个ESU。
遗传距离计算的结果支持3个ESU的划分(表5),3个ESU内两两居群间的遗传距离≤1.182 (对角线以下,不加粗部分),ESU之间的两两居群间的遗传距离≥1.399(对角线以下,加粗部分)。居群内平均遗传距离为0~0.327,ESUⅠ、Ⅱ、Ⅲ内平均遗传距离分别为0.000、0.001、0.000。91个个体两两的遗传距离为0.000~0.013。
对3个ESUs进行AMOVA分析和相应的F检验的结果(表6)可见,遗传变异主要存在于ESUs间,所占比例高达94.22%,ESUs内居群间的遗传变异仅占5.09%,居群内的遗传变异几乎可以忽略不计,仅为0.69%,因此,AMOVA的结果也支持胡黄连3个ESUs的划分。
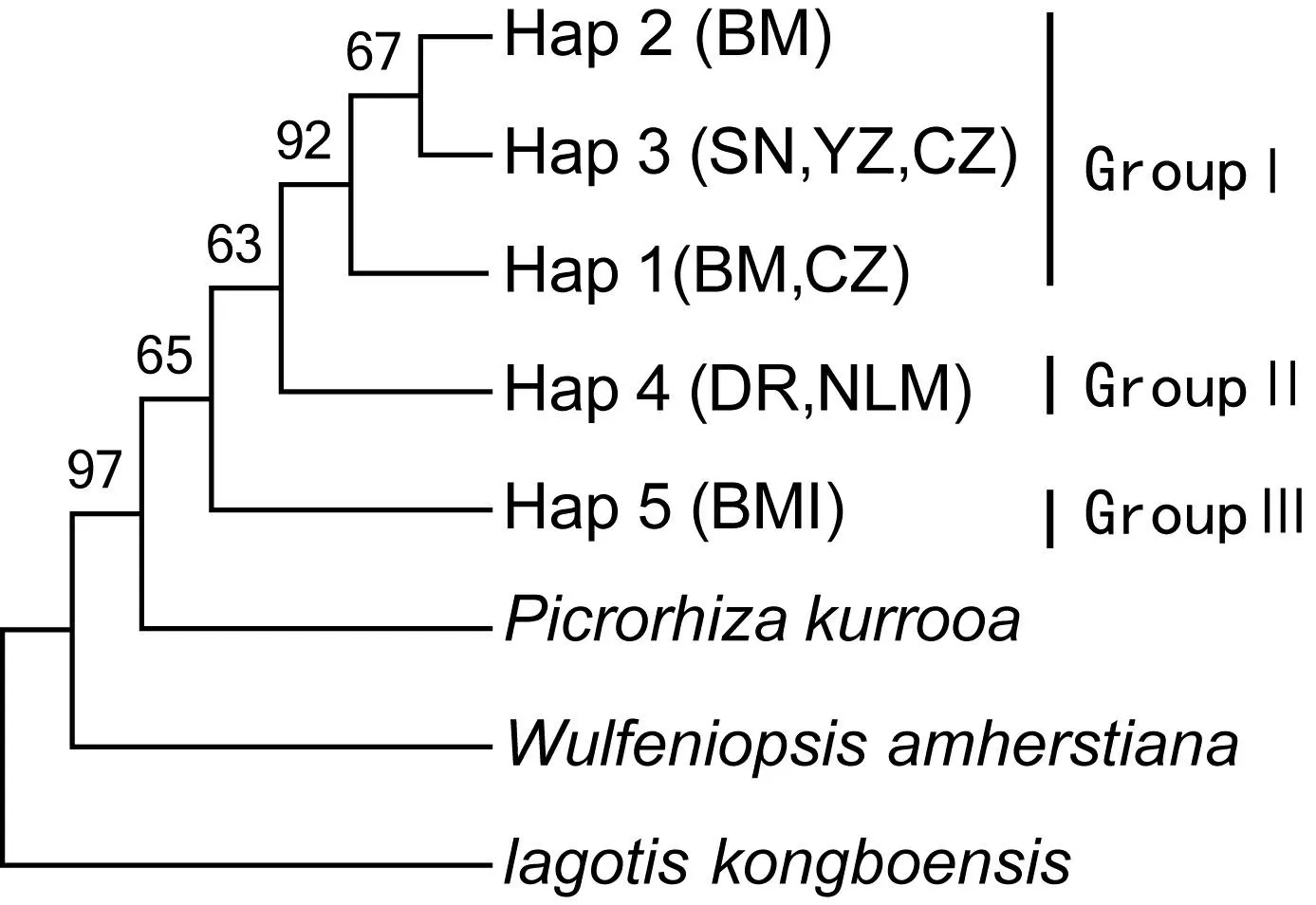
图 2 胡黄连5个单倍型的多数一致性树,显示50%以上的支持Fig. 2 Majority-rule consensus of 5 cpDNA haplotypes of N. scrophulariiflora, showing more than 50% of the support
3讨论
3.1 胡黄连遗传分化程度
胡黄连的cpDNAtrnL-F片段检测结果发现,在单倍型序列的13个变异位点中,小片段的插入/缺失仅占了1个,而点突变占了12个。由此表明了胡黄连单倍型序列间的分化水平较高,种内遗传分化经历的时间较长。
对胡黄连整个地理分布区的7个居群91个个体进行的trnL-F基因间区检测发现,胡黄连总的遗传多样性(HT= 0.861)和居群间遗传分化程度都很高(Gst= 0.916)。这与ISSR分子标记的分析结果一致(liu et al, 2011)。居群间遗传分化程度较之大多数物种高,如条纹狭叶龙胆(Metagentianastriata),Gst=0.859(陈生云等,2008)、偏花报春(Primulasecundiflora), Gst = 0.816(Wang et al, 2008)、肋果沙棘(Hippophaeneurocarpa),Gst= 0.646(孟丽华等,2008)等。胡黄连的cpDNA多样性水平高于Petit et al(2005)所统计的175种种子植物的平均cpDNA多样性HT= 0.67。AMOVA分析也表明,胡黄连99.22%遗传变异发生在居群间,基因流仅为 0.04, 说明胡黄连具有很高的遗传分化水平,而且居群间几乎没有基因交流。Ouborg et al(1999)认为种群间一粒种子或一个花粉粒的交流就会导致Gst值达到0.20,这说明胡黄连种群间存在极低的花粉或种子交流。根据cpDNA得出的Gst值反映种子迁移对基因流的贡献,而根据核基因组分子标记ISSR计算出的Gst值既反映了种子迁移对基因流的贡献,同时也反映了花粉运动对基因流的贡献。因此对于胡黄连而言,花粉运动对基因流的贡献要比种子稍大。
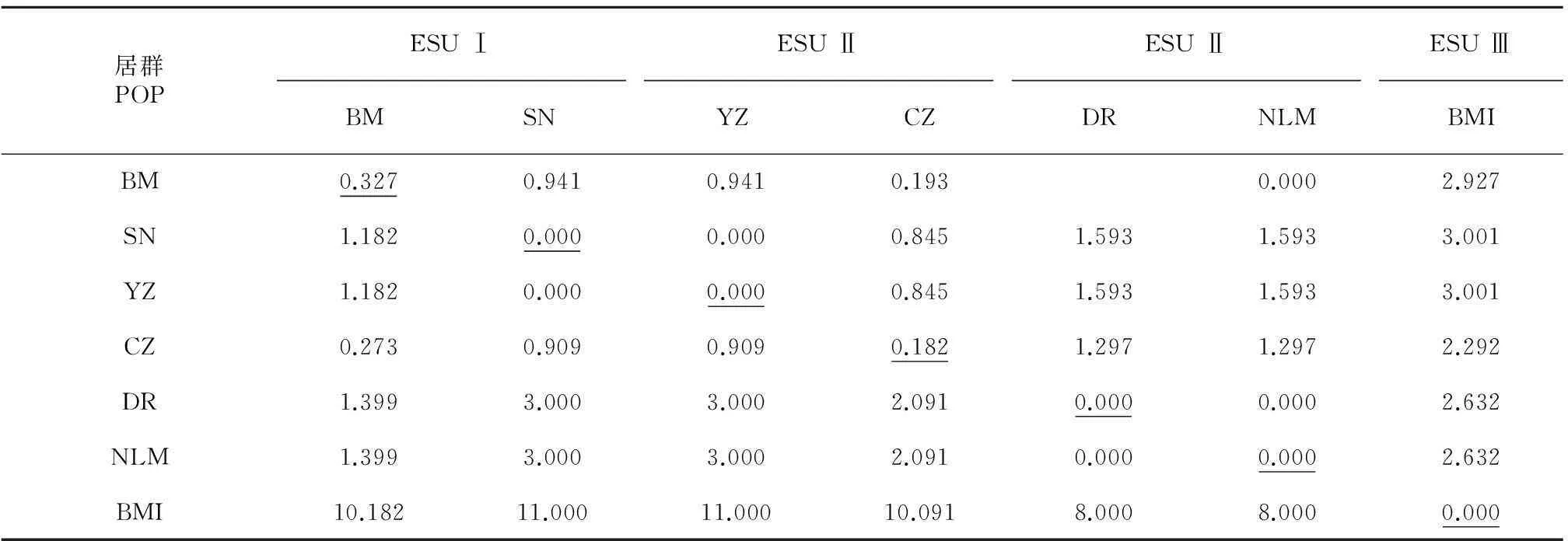
表 5 胡黄连7个居群间和居群内的遗传距离
注:对角线下为居群间遗传距离, 对角线上为标准误,下划线为居群内遗传距离。
Note: Among populations distances are below the diagonal(mean among the ESU). SE values are above diagonal. Within population mean distances are underlined.

表 6 胡黄连7个居群不同层次AMOVA分析
注:显著性检验用1 023次置换。
Note: Significance tests using 1 023 permutations.
胡黄连居群间如此大的遗传分化和如此低的基因流这可能与喜马拉雅和横断山区的生境复杂度有关。5个单倍型在2个地区中的分布呈现较大差异,所有单倍型在2个地区间均不共享,而是为各自独有,在西藏地区,聂拉木(NLM)和定日(DR)共享一个单倍型(H4),居群波密 (BMI) 居群独有一个单倍型(H5),从胡黄连单倍型多数一致性树(图2)可见西藏的2个单倍型不构成一个单系,这一结果显然也说明胡黄连存在分化明显的遗传谱系, BMI与P.kurrooa聚为一支,这是否意味着该属分类地位上存在一定的疑问?有待于深入研究。
3.2 显著进化单元的划分及胡黄连保护
本研究根据多数一致性树划分的3个ESUs, 从居群间的遗传距离矩阵也得到很好的支持,所有3个ESU内两两居群间的遗传距离小于ESU之间的两两居群间的遗传距离(分别是≤1.182和≥1.399),表明ESUs内的居群之间遗传分化程度低,ESUs之间遗传分化程度高。
ESUⅠ 包括了4个居群分别是BM、CZ、YZ、SN,占有3个单倍型(H1、H2、H3), 从单倍型在这个4个居群的分布来看,必须优先保护的是BM和CZ居群。其中BM居群的保护任务是最紧迫的,因为BM居群具有独特的单倍型H2,而CZ包含了H1、H32个单倍型,因此保护这2个居群就等于保护了所有的遗传变异。目前BM居群分布于白马雪山国家级自然保护区内,但由于此居群与胡黄连的其它居群相比,交通相对便利,云南省迪庆州德钦县的藏医日常使用常在此处采挖,保护区由于地域广阔,人力物力限制,对濒危植物的保护相对较薄弱,因此此居群境况堪忧。按照ESU的保护原则是种群的遗传组成越独特,越具有优先保护价值,因而BM居群是需要重点保护的居群,因为这个居群的消失会对胡黄连的遗传多样性造成重大影响。CZ居群离附近的村庄比较远,居群附近生境恶劣,鲜有人烟活动,受到威胁的可能性较小。
ESUⅡ包括了DR和NLM2个居群,只有这2个居群具有单倍型H4,因此也具有独特性。NLM居群分布于聂拉木县城附近,每年都有药材收购商到当地收购胡黄连药材,当地药农往往组织成小分队的形式进行采挖,因此对NLM居群的干扰特别大,而DR居群是位于日喀则地区定日县融霞乡的一个居群,多年前被定日县的藏医采收过,由于资源已严重匮乏,当地藏医已多年未曾来此地采集,此居群由于地处偏远、交通不便受人类干扰相对较小,但此居群很小,能够自然复壮的几率很低。这2个居群已经处于珠穆朗玛峰国家级自然保护区内,尽管此保护区主要保护高山、高原生态系统,但由于地域广阔,对胡黄连的保护没有受到足够重视。综上所述,DR居群可以采用就地保护措施,而对严重受人类干扰的NLM居群的最佳保护措施则是迁地保护,由于两者具有共同的单倍型,因此2个居群能够保护好一个居群即可。
ESUⅢ 只包含一个居群即BMI居群,并且具有一个独特的单倍型H5, 结合ISSR分子标记研究结果,BMI居群是所有居群中遗传多样性水平最低的一个(PPB=4.88),因而此居群胡黄连具有重要的保护价值。在调查中发现此居群胡黄连数量已非常少,呈零星分布,距离最近的是波密县城,波密县以旅游业为主,未见藏医院以及藏医,因而此地对胡黄连的利用很少,但胡黄连为何会数量如此之少,原因现无从得知。从保护遗传差异性居群角度,此居群由于遗传的特异性,需要就地保护。
由于胡黄连的遗传多样性主要分布在居群间,若有条件在高海拔地区建设胡黄连种质资源圃对胡黄连进行迁地保护时,样品采集应该覆盖尽可能多的居群而避免从一2个居群采集多个个体。
参考文献:
CHINA PHARMACOPOEIA COMMITTE, 2010. Pharmacopoeia of the People′s Republic of China [M]. 2010 Ed. Beijing:Chemical Industry Press. [国家药典委员会, 2010. 中华人民共和国药典 [M]. 2010版. 北京:化学工业出版社.]
CHEN SY, WU GL, ZHANG DY, et al, 2008. Molecular phylogeography of alpine plantMetagentianastriata(Gentianaceae) [J]. J Syst Evol, 46(4): 573-585. [陈生云, 吴桂莉, 张得钧, 等, 2008. 高山植物条纹狭蕊龙胆的分子亲缘地理学研究 [J]. 植物分类学报, 46(4): 573-585.]
CHEN SH, CHEN C, GUO CG, et al, 2008. Domestic study ofPicrorhizascrophulariilforaPennell at different elevation [J]. SW Chin J Agric Sci, 21(5): 1 391-1 394. [杨少华, 陈翠, 郭承刚, 等, 2008. 胡黄连不同海拔定位栽培研究 [J]. 西南农业学报, 21(5): 1 391-1 394.]
CHEN SH, CHEN C, GUO CG, et al, 2009. Study of biological characters,endangered reasonsand protecting measures ofPicrorhizascrophulariifloraPennel in Yunnan [J]. SW Chin J Agric Sci, 22(5): 1 482-1 485. [杨少华, 陈翠, 郭承刚, 等, 2009. 云南胡黄连生物学特性、濒危原因及保护对策 [J]. 西南农业学报, 22(5): 1 482-1 485.]
CHIANG TY, CHIANG YC, CHEN YJ,et al, 2001. Phylogeography ofKandeliacandelineast Asiatic mangroves based on nucleotide variation of chloroplast and mitochondrial DNAs [J]. Mol Ecol, 10(11): 2 697-2 710.GAO HW,KUANG HX,YAN XY, 2011. Absorption characteristics of picroside Ⅱ in rat intestine in situ [J]. Drugs & Clinic,26(4):284-286. [高宏伟,匡海学,阎雪莹, 2011. 胡黄连苦苷Ⅱ大鼠在体肠吸收特征研究 [J]. 现代药物与临床,26(4):284-286.]DOYLE JJ, 1991. DNA protocols for plants-CTAB total DNA isolation [M]// HEWITT GM, Johnston A (Eds). Molecular Techniques in Taxonomy, Berlin, Germany, Spinger-Verlag: 283-293.
HUANG KY,HE L, WANG DC, et al, 2008. Chemical constituents ofPicrorhizascrophulariilfora[J]. Chin Pharm J, 43(18): 1 382-1 385. [黄开毅,何乐, 王大成, 等, 2008. 西藏胡黄连的化学成分 [J]. 中国药学杂志, 43(18): 1 382-1 385.]
LIU XL, QIAN ZG, LIU FH,et al, 2011.Genetic diversity within and among populations ofNeopicrorhizascrophulariiora(Scrophulariaceae) in China, an endangered medicinal plant [J]. Biochem Syst Ecol , 39(4-6): 297-301.LIU Y, YANG SX,GAO LZ, 2010. Comparative study on the chloroplast RPL 32-TRNL nucleotide variation within and genetic differentiation among ancient tea plantations ofCamelliasinensisvar.assamicaand C1 taliensis ( Theaceae) from Yunnan, China [J]. Acta Bot Yunnan, 32(5): 427- 434. [刘阳, 杨世雄,
(Continueonpage706)(Continuefrompage697)
高立志, 2010. 云南古茶园栽培大叶茶和大理茶群体的叶绿体 [J]. 云南植物研究, 32 ( 5): 427- 434.]
MENG LY, YANG HL, WU GL, et al, 2008. Genetic structure and phylogeography of populations ofAlsophilapodophyllainHainan and Guangdong,southern China, based on cpDNA trnL-F noncoding sequences [J]. J Syst Evol, 46(1): 32-20. [孟丽华, 杨慧玲, 吴桂丽, 等, 2008. 基于叶绿体DNA trnL-F序列研究肋果沙棘的谱系地理学 [J]. 植物分类学报, 46(1): 32-20.]
MORITZ C, 1994. Defining “Evolutionarily Significant Units” for conservation [J]. Trend Ecol Evol, 9(10): 373-375.
OUBORG NJ, PIQUOT Y,VAN GROENENDAEL JM, 1999. Population genetics, molecular markers and the study of dispersal in plants [J]. J Ecol, 87(4): 551-568..
PETIT RJ, DUMINIL J, FINESCHI S,et al, 2005.Comparative organization of chloroplast, mitochondrial and nuclear diversity in plant populations [J]. Mol Ecol, 14(3): 689-701.
SU YJ, WANG T, ZHENG B, et al, 2004. Genetic structure and phylogeography of populations ofAlsophilapodophyllainHainan and Guangdong, southern China, based on cpDNA trnL-F noncoding sequences[J]. Acta Ecol Sin, 24(5): 914-919. [苏应娟, 王艇, 郑博, 等, 2004. 根据cpDNA trnL-F非编码区序列变异分析黑桫椤海南和广东种群的遗传结构与系统地理. 生态学报, 24(5): 914-919.]
WANG FY, GONG X, HU CM, et al, 2008. Phylogeography of an alpine speciesPrimulasecundiflorainferred from the chloroplast DNA sequence variation [J]. Acta Phytotax Sin, 46(1):13-22.
Conservative genetics ofNeopicrorhizascrophulariiflorabased on cpDNAtrnL-F
LI Guo-Dong1, YIN Zi-Li2, LIU Xiao-Li1*
( 1.CollegeofTraditionalChinesePharmacy,YunnanUniversityofTraditionalChineseMedicine, Kunming 650500, China;2.CollegeofTraditionalMinorityMedicine,YunnanUniversityofTraditionalChineseMedicine, Kunming 650500, China )
Abstract:Neopicrorhiza scrophulariiflora (Scrophulariaceae), a monotypic genus perennial species, is endemic to the Eastern Himalayas and the Hengduan Mountains region. It only distributes in Yunnan and Tibet in China, ranging from 3 600 m to 4 200 m in elevation. The long and creep rhizomes (Rhizoma Neopicrorhizae) are of high medicinal value and dysentery by traditional Chinese and Tibetan medicine. Mainly because of large-scale acquisitions activity, natural populations of this species have suffered rapid declines and now it is classified as an endangered species under second category of key protected wild plants in China. In order to protect the decreasing natural genetic resources of N. scrophulariiflora, in this study, the chloroplast DNA (cpDNA) trnL-F noncoding sequence was used to estimate the genetic diversity and genetic structure and the evolutionary significant units (ESU) were proposed. A total of 91 individuals of N. scrophulariiflora were collected from seven populations, covering almost all areas of its distribution ranges. Of these seven populations, four were from Yunnan Province and three populations were from Tibet. The statistical results showed that the haplotype sequences length varied from 871 bp to 876 bp. A total of five haplotypes were detected based on trnL-F nucleotide variation. Yunnan contains three haplotypes and Tibet contains two. However, none of common haplotypes were shared between the populations from Yunnan and Tibet. A normal low level of genetic diversity (Hd = 0.434 19) and nucleotide diversity (Dij = 0.004 66) were identified at the species level. A high level of genetic differentiation (0.96) among populations was revealed. AMOVA results from chloroplast data indicated that 0.78% of the genetic variation was partitioned within population, 60.97% among populations within groups, and 38.25% among groups under the condition that N. scrophulariiflora was divided into two groups including Yunnan and Tibet. The U-statistic test for phylogeographical structure showed that NSTwas significantly higher than GST(NST>GST, P < 0.01), which suggested a distinctly phylogeographical pattern The gene flow (Nm) was extremely low with only 0.04. The higher NSTthan GST(P<0.01) suggested a distinctly phylogeographical pattern. Conjoint Fst (0.864 520), gene flow, GSTand AMOVA results all indicated a significant high level of genetic differentiation among populations, which could be a consequence of the limited gene flow caused by geographic isolation among populations. Phylogenetic analysis of the haplotypes sequences identified three tentative clades (Ⅰ,Ⅱ and Ⅲ) according to Majority-rule consensus tree. All of which had distinct geographic range: Clade Ⅰ comprised four populations (CZ, YZ, SN, BM) which were located at the Hengduan Mountains region; Clade Ⅱ comprised one population (BMI), which was located at the Eastern Himalayas region; and Clade Ⅲ comprised two populations (DR, NLM) located at the Central Himalayas region. Based on the phylogenetic analyses and uniqueness of the populations, three evolutionary significant units (ESU) were identified and conservation strategies were discussed for this endangered species. Baimaxueshan and Cizhong, Bomi, Nielamu and Dingri populations respectively concluded in the three evolutionary significant units and the five populations all contained special haplotypes. Based on these findings, all the populations should be protected. However, in consideration of the actual distribution of every population, Baimaxueshan population from Yunnan and Bomi population from Tibet should be given priority for conservation and in situ conservation should be the ideal implement.
Key words:Neopicrorhiza scrophulariiflora, endangered, trnL-F, haplotype, conservative genetics
DOI:10.11931/guihaia.gxzw201410004
收稿日期:2014-10-8修回日期:2015-03-29
基金项目:云南省自然科学基金面上项目(2010CD073);国家科技基础性工作专项重点项目(SB2007FY0200);云南省应用基础研究青年项目(2014FD035)[Supported by the Natural Science Foundation of Yunnan(2010CD073); National Science and Technology Basic Special Fund (SB2007FY0200); Yunnan Youth Program for the Application Foundamental Research (2014FD035)]。
作者简介:李国栋(1984-), 江西鄱阳人,博士,讲师,主要从事植物谱系地理学研究,(E-mail)gammar116@163.com。 *通讯作者:刘小莉,博士,副教授,硕士生导师,主要从事药用植物资源研究,(E-mail)kmxunzi@aliyun.com。
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2016)06-0691-07
李国栋,尹子丽,刘小莉. 基于cpDNAtrnL-F序列的胡黄连保护遗传学研究 [J]. 广西植物, 2016, 36(6):691-697
LI GD,YIN ZL,LIU XL. Conservative genetics ofNeopicrorhizascrophulariiflorabased on cpDNAtrnL-F [J]. Guihaia, 2016, 36(6):691-697
