紫薇异倍体的杂交亲和性分析及子代倍性鉴定
2016-07-18段丽君叶要妹
王 滑,陈 放,段丽君,叶要妹
(园艺植物生物学教育部重点实验室/华中农业大学园艺林学学院,湖北 武汉 430070)
紫薇异倍体的杂交亲和性分析及子代倍性鉴定
王滑,陈放,段丽君,叶要妹*
(园艺植物生物学教育部重点实验室/华中农业大学园艺林学学院,湖北 武汉 430070)
摘 要:以二倍体红薇(H,2n)、三倍体紫薇(Z,3n)、四倍体紫薇(Z,4n)及四倍体银薇(Y,4n)为试材,进行杂交试验,检测紫薇异倍体杂交的亲和性,并采用细胞学倍性鉴定法对杂交子代的倍性进行鉴定和比较。结果显示:不同杂交组合子代的结实特性以及种子的出苗率均存在差异,其中 Z(4n)×H(2n)的单果重(0.22 g)、单果种子数(40.1)、千粒重(1.79 g)及出苗率(36.4%)均高于 H(2n)×Y(4n),且差异达到显著水平,表明在二倍体与四倍体的杂交中,四倍体作为母本的亲和性更高;H(2n)×Z(3n)后代倍性主要为非整倍体,占80%以上,其次是2n,占11.8%;Y(4n)×H(2n)后代倍性主要为3n,占50%,其次是非整倍体,占40%;H(2n)×Z(3n)后代中,1株为3n,2株为非整倍体;Y(4n)×H(2n)后代中,6株为3n,3株为非整倍体;H(2n)×Y(4n)后代幼苗中,1株为2n,1株为3n;Z(4n)×Z(4n)后代中,1株为4n;在杂交组合H(2n)×Z(3n)、Y(4n)×H(2n)及H(2n)×Y(4n)后代中还出现三子叶变异的幼苗。
关 键 词:紫薇;异倍体;杂交亲和性;倍性鉴定法
投稿网址:http://xb.ijournal.cn
紫薇(Lagerstroemia indica)是千屈菜科紫薇属多年生落叶灌木或小乔木。紫薇属植物约有55种, 260个品种,中国有18个种和40多个品种,占世界紫薇总资源的32.7%[1]。紫薇具有花色艳丽,花期长等特点,是主要的木本观赏植物之一,在国内外园林中得到广泛应用[2]。但中国的栽培种中缺少大叶、大花、密花的品种,而多倍体植株具有树势健壮、抗性强、叶大、花大、无籽或少籽等优良性状[3–4],能拓宽遗传资源。目前,多倍体诱变技术已经成功地应用于紫薇育种,主要方式是利用秋水仙素培育出四倍体紫薇,将其与二倍体紫薇进行杂交获得三倍体植株。与二倍体紫薇相比,三倍体及四倍体植株的花蕾、花径增大,花瓣变长变宽,观赏价值大大优于二倍体[5–6]。紫薇种间杂交授粉结实容易。蔡明等[7]利用尾叶紫薇、紫薇与散沫花进行种间杂交,培育出具有早花、香花性状的紫薇品种。随着杂交育种研究工作的不断深入,杂交成功率显著提高,杂交亲本的应用范围不断扩大,远缘杂交育种与多倍体育种技术相结合,已成为观赏植物商业育种的重要手段[8–9]。利用倍性不同的材料杂交有望培育优良性状的紫薇新品种。然而,异倍体杂交存在一定的亲和性障碍[10–11]。目前,对于不同倍性紫薇的杂交亲和性以及杂交子代倍性鉴定鲜见报道。本研究以紫薇为试材,通过测试不同倍性紫薇杂交对亲和性的影响,拟筛选出多倍体紫薇杂交的最佳亲本组合,同时利用已有的细胞学倍性鉴定方法[12]对多倍体杂交子代的倍性进行鉴定和比较,旨在为紫薇异倍体杂交育种提供参考依据。
1 材料与方法
1.1 材料
以华中农业大学园艺林学学院花卉基地5~6年生,长势健壮的二倍体红薇(H,2n)、三倍体紫薇(Z,3n)、四倍体紫薇( Z,4n)及四倍体银薇(Y,4n)为试验材料,其中四倍体紫薇与银薇是由二倍体紫薇与银薇经秋水仙素诱导加倍得到,三倍体紫薇是二倍体紫薇与四倍体紫薇的天然杂交种。
1.2 方法
1.2.1 异倍体杂交试验
选择长势健壮、株型良好H(2n)、Z(3n)、Z(4n)及Y(4n)作为母株,采用完全双列杂交试验设计进行异倍杂交。但由于受材料限制,所做的异倍杂交组合见表1。于2010年9月紫薇盛花期08:00—10:00人工授粉,套袋标记,1周后去袋。每组合3次重复。每重复授15朵花。11月采种,晾晒,净种后于4 ℃冰箱中贮藏,备用。

表1 异倍杂交组合设计Table 1 Designs of alloploid cross combinations
于2011年3月田间播种、栽培管理、定株编号。最终获得的异倍杂交幼苗:H(2n)×Z(3n)杂交后代幼苗17株;Y(4n)×H(2n)杂交后代幼苗20株;H(2n)×Y(4n)杂交后代幼苗3株;Z(4n)×Z(4n)杂交后代幼苗3株。
1.2.2 不同杂交组合亲和性分析
采种后按不同杂交组合编号分类,用游标卡尺测量蒴果形态特征,并在2011年4月,观测紫薇种子出苗率,以此作为杂交亲和性的指标。
1.2.3 异倍杂交后代幼苗倍性分析
对异倍杂交后代幼苗定株编号,8—9月生长盛期,于晴天10:00采集不同编号植株的功能叶3~5片,立即带回实验室,进行幼苗形态特征、保卫细胞及气孔大小观测[12],具体方法如下:用透明胶带粘贴叶片下表皮中部,取3~5 mm×3~5 mm的矩形块,下表皮朝上平放在载玻片上,滴2滴I2–KI染色,30 min后置于倒置显微镜下观察测定。在10×40倍显微镜下拍照,用Image–Pro Plus 6.0软件测量保卫细胞及气孔的长度和宽度,每株植株随机观察10个气孔,取其平均值。
室内试验于华中农业大学园艺林学学院森林培育与林木育种实验室进行。
1.3 数据分析
数据采用Excel2003进行整理,运用SPSS17.0软件进行方差分析。
2 结果与分析
2.1 不同杂交组合的杂交亲和性
2.1.1 异倍杂交组合的结实特性
9个杂交组合的结实特性(表2)显示,杂交后代的结实特性,如果实长径、短径、单果重、单果种子数都有一定的变异。以二倍体为母本的H(2n)×H(2n)、H(2n)×Z(3n)及H(2n)×Y(4n)组合中,果实长径以 H(2n)×H(2n)组合的最长(10.32 mm),H(2n)×Z(3n)组合的果实长径变异系数最大(11.81%);果实短径以H(2n)×Y(4n)组合的最长(9.23 mm),果实短径变异系数最大(11.42%)的组合是H(2n)×Z(3n),H(2n)×Z(3n)组合的单果质量最大(0.19 g),单果重的变异系数最大(41.39%)的组合是H(2n)×Z(3n),单果种子数以H(2n)×Y(4n)组合的最多,为 32.1粒,单果种子数变异系数最大(30.3%)的组合是H(2n)×Z(3n),以三倍体为母本的 Z(3n)× H(2n)、Z(3n)×Z(3n)及Z(3n)×Z(4n)组合中,果实长径最长的组合是Z(3n)×Z(4n),为10.67 mm,果实长径的变异系数最大的是Z(3n)×H(2n),为10.79%;果实短径以Z(3n)×Z(4n)的最大,为9.33 mm,果实短径的变异系数最大的是Z(3n)×H(2n),为15.15%;单果以Z(3n)×Z(4n)的质量最大,为0.19 g,单果质量的变异系数最大的组合是 Z(3n)×H(2n),为41.49%;单果种子数及其变异系数均是Z(3n)×H(2n)的最大,分别为28.7粒和33.1%。以四倍体为母本的Z(4n)×H(2n)、Z(4n)×Z(3n)及Z(4n)×Z(4n)组合中,以Z(4n)×Z(4n)组合的果实长径最长(10.42 mm),短径最长,单果质量最大,单果种子数最多,分别是10.42 mm、9.94 mm、0.25 g和40.1粒,果实长径的变异系数最大(16.13%)的组合是Z(4n)×H(2n),果实短径的变异系数最大的组合是 Z(4n)×H(2n),为11.04%,单果重的变异系数最大的是Z(4n)×Z(3n),为34.35%,种子数的变异系数最大的是Z(4n)×H(2n),为28.03%。9个杂交组合的种子千粒重以Z(4n)×Z(4n)组合最大(1.97 g),Z(3n)×H(2n)的千粒重最小(1.01 g)。
从表2可见,不同杂交组合间的种子特性有较大的差异。Z(4n)×H(2n)的果实长径(9.64 mm)最小,显著地小于其他杂交组合的。Z(3n)×H(2n) 的果实短径最小,为7.92 mm,显著地小于其他组合的。Z(4n)×Z(4n)的杂交后代单果质量最大,为0.25 g,而H(2n)×H(2n)杂交后代的单果质量最小,为 0.13 g。H(2n)×Y(4n)单果种子数最多,为 40.1粒,Z(3n)×Z(4n)杂交的单果种子数最少,为22.3粒。二倍体为母本的组合,2n×3n单果质量和单果种子数均显著地低于2n×4n组合,且单果质量和单果种子数的变异系数也最大;以三倍体为母本的组合,3n×4n的单果质量显著地高于其他2个组合。

表2 不同杂交组合的果实性状及变异特征Table 2 Fruiting characteristics and variation features among different cross combinations in L. indica
2.1.2 异倍体杂交组合的出苗率
所有杂交组合穴盘播种的出苗率显示,Z(4n)× H(2n)的出苗率最高,为36.4%;其次为H(2n)×H(2n)出苗率为35.2%;Z(3n)×Z(3n)组合的出苗率最低为8.7%。从以上结果可以看出来,以二倍体为母本的组合,2n×3n穴盘出苗率(21.9%)高于2n×4n(12.5%)的;以三倍体为母本的组合,3n×4n穴盘出苗率最高,3n×3n的穴盘出苗率最小;以四倍体为母本的组合,4n×4n穴盘出苗率较低,为 20.5%,4n×2n的穴盘出苗率最高,为36.4%。异倍杂交组合2n×4n、4n×4n虽能够获得较多的种子,但是正常发育的种子较少,导致出苗率较低。而异倍杂交组合4n×2n获得正常种子数较多,出苗率也较高,这说明了异倍杂交组合2n×4n、4n×4n的杂交亲和性低于4n×2n杂交组合。为了获得更多的杂交后代幼苗,异倍杂交时应以4n为母本,2n为父本。
2.2 异倍杂交后代幼苗子叶的变异
在所有杂交组合中,只在 H(2n)×Z(3n)、Y(4n)×H(2n)及 H(2n)×Y(4n)杂交后代的幼苗期,分别发现有三子叶变异的幼苗3、2和1株(图1)。观察发现,3个子叶大小相近,在同一节间分布均匀,呈120°。第一轮3片真叶分布在同一节间,同时长出。第二轮3片真叶有2片位于同一节间,而另一片的着生位置稍高于其他2片,以后随着幼苗的生长逐渐变为正常植株。

图1 紫薇杂交后代中的三子叶幼苗Fig. 1 Tricotyledon seedlings of cross progeny in L. indica
2.3 异倍杂交后代幼苗倍性分析
参照Ye 等[12]的标准(表3),对17株H(2n)× Z(3n)、20株Y(4n)×H(2n)、3株H(2n)×Y(4n)及3株Z(4n)×Z(4n)杂交后代幼苗气孔宽度和密度指标进行分类(表4)。结果显示,H(2n)×Z(3n)杂交后代倍性分离均主要集中在非整倍体,比例为80%,其次是二倍体,比例为11.8%。Y(4n)×H(2n)杂交后代幼苗倍性三倍体的比例最大,比例为50%;其次是非整倍体,比例占40%。H(2n)×Y(4n)杂交后代幼苗2株为三倍体,1株为非整倍体。Z(4n)×Z(4n)杂交后代幼苗倍性均为四倍体。
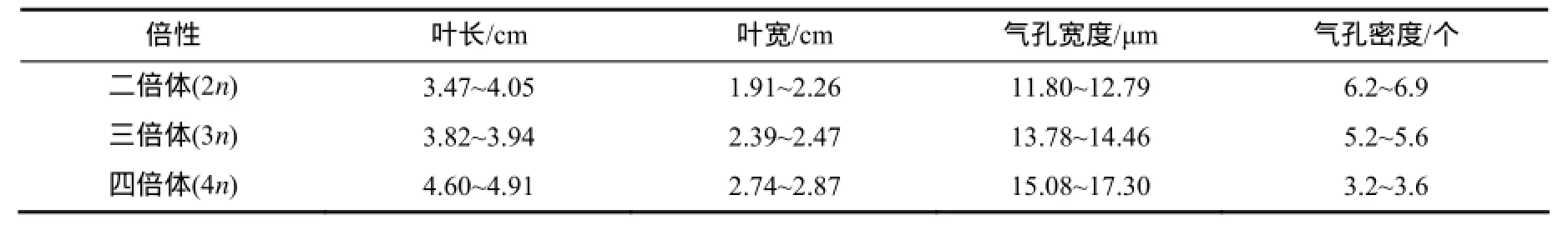
表3 紫薇倍性鉴定的初步参考标准Table 3 Primary reference standards of ploidy identification in L. indica
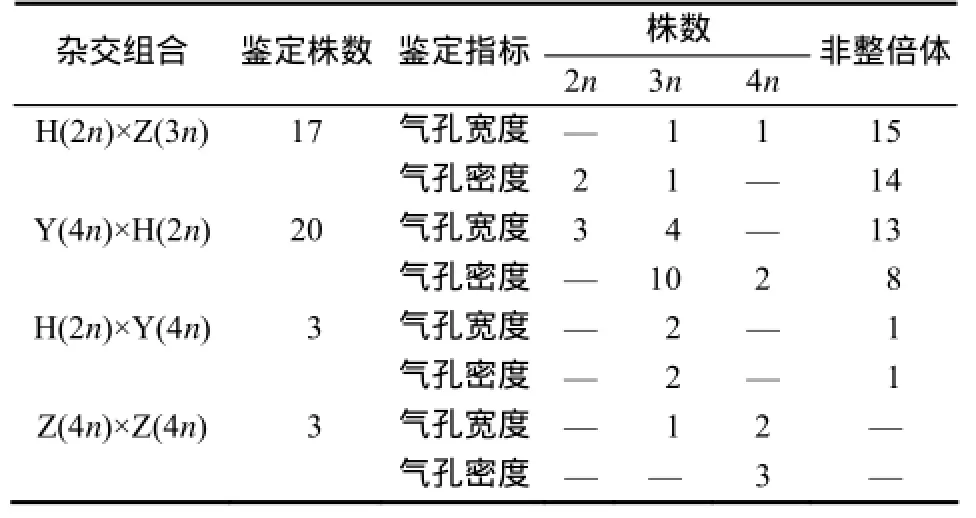
表4 不同异倍体杂交组合后代倍性归类结果Table 4 Classification of ploidy level in different alloploidcrossprogeny
3 讨论与结论
不同倍性杂交亲和性试验发现,异倍杂交2n×4n、4n×4n虽能够获得较多的种子,但是正常发育的种子较少,导致出苗率较低。而异倍杂交组合4n×2n获得正常种子数较多,出苗率也较高,所以异倍杂交组合 2n×4n、4n×4n的杂交亲和性低于4n×2n杂交组合。异倍杂交亲和性的研究在其他植物上也有报道,但杂交亲和性结果因材料不同而不一致。本试验由杂交组合 2n×4n、4n×2n分别获得的3n在大多数性状上没有差异,但是4n×2n杂交组合获得的种子萌发率高于 2n×4n杂交组合,与Burton和Husband[13]在对柳兰进行异倍杂交时发现结果一致。Viloria[14]也曾报道,Citrus aurantifolia异倍杂交组合4n×2n与2n×4n比较,杂交后代中三倍体的发生率要高。结合遗传理论和本试验结果来看,培育三倍体紫薇应以四倍体作母本的4n×2n为重点杂交组合。郭印山等[15]通过人工授粉和常规压片法对2n和4n葡萄杂交组合的亲和性进行研究,结果 2n×4n后代的种子形成率及出苗率都高于4n×2n的后代,本研究结果与其不一致。
根据Johnston[16]提出的EBN假说,不同倍性间杂交不育,可能与胚或胚乳染色体比例被破坏有关。胚乳中,只有当母本与父本的基因组成比例为2∶1时,胚乳才能正常发育,形成健全的种子,否则胚乳发育中途停滞,造成败育。在本研究中,紫薇异倍杂交组合4n×2n的亲本在生殖过程中产生的配子比例为2∶1,形成的种子胚乳中母本与父本的基因组成比例为4∶1,而2n×4n产生的配子比例为1∶2,种子胚乳中母本和父本的基因比例为1∶1,所以导致种子的胚乳和胚发育不全。然而杂交组合4n×2n的种子出苗率却高于杂交组合2n×4n的,这可能是因为用染色体数目高的作母本,胚与胚乳染色体比例较正常的染色体比例偏离少的缘故。异倍杂交后代中,倍性均出现分离。二倍体、三倍体与四倍体紫薇由于细胞内染色体组的倍性差异,导致H(2n)×Z(3n)后代倍性主要为非整倍体,Y(4n)×H(2n)后代倍性主要为3n,其次是非整倍体。这在鸭茅[17]、水稻[18]也发现相似的倍性分离。
子叶数是植物进化及分类的一个重要指标,生物学工作者发现在自然情况下可发生子叶变异,但频率较低。三子叶现象已在桑树[19]、凤仙花[20]、向日葵[21]等异倍杂交后代中都有发现。本研究发现在H(2n)×Z(3n)后代、Y(4n)×H(2n)后代及H(2n)×Y(4n)后代幼苗中均有三子叶幼苗出现,直至第3轮真叶,以后随着幼苗的生长逐渐变为正常植株。根据Sacita[22]提出的三子叶形成原因,结合本试验是进行异倍杂交,后代三子叶幼苗产生的原因可能是异倍杂交过程中细胞异常分化,相连的2个胚部分融合导致的。
参考文献:
[1] 陈俊愉.中国花卉品种分类学[M].北京:中国林业出版社,1999.
[2] 张启翔,田苗.我国紫薇品种调查研究[C]//中国园艺学会观赏园艺专业委员会.中国园艺学会观赏园艺专业委员会学术年会论文集.北京:中国林业出版社,2008:56–65.
[3] 康向阳.林木多倍体育种研究进展[J].北京林业大学学报,2003,25(4):70–74.
[4] 王尚德,王晓川,刘佳棽,等.桃三倍体种质的花粉形态及萌发特性[J].北方园艺,2015(12):80–84.
[5] 童俊,叶要妹,冯彪,等.秋水仙素诱导三种紫薇多倍体的研究[J].园艺学报,2009,36(1):127–132.
[6] 袁玮.紫薇授粉生物学特性和三倍体育种初步研究[D].武汉:华中农业大学,2009.
[7] 蔡明,孟锐,潘会堂,等.紫薇属与散沫花属远缘杂交亲和性的研究[J].园艺学报,2010,37(4):637–642.
[8] Van T J M,Kibuyng L.Interspecific hybridisation and polyploidisation as tools in ornamental plant breeding[J]. Acta Hort,2003,612:13–22.
[9] 王金凤,柳新红,陈卓梅.紫薇属植物育种进展研究[J].园艺学报,2013,40(9):1795–1804.
[10] 段超,张启翔.观赏植物远缘杂交和多倍体育种研究进展[J].安徽农业科学,2009,37(15):6954–6956.
[11] Nico de Storme,Linda Z,Martin M,et al.Volume-based pollen size analysis:an advanced method to assess somatic and gametophytic ploidy in flowering plants[J]. Plant Reproduction,2013,26(2):65–81.
[12] Ye Y M,Tong J,Shi X P,et al. Morphological and cytological studies of diploid and colchicines-induced tetraploid lines of crape myrtle(Lagerstroemia indica L.)[J].Scientia Horticulture,2010,124(1):95–101.
[13] Burton T L,Husband B C.Fitness differences among diploids,tetraploids,and their triploid progeny in Chamerionangustifolium:mechanisms of inviability and implications for polyploid evolution[J].Evolution,2000,54(4):1182–1191.
[14] Viloria Z.Acid Citrus fruit improvement via interploid hybridization using allotetraploid somatic hybrid and autotetraploid breeding parents[J].Journal of American Society for Horticultural Science,2005,130(3):392–402.
[15] 郭印山,郭修武,李坤,等.葡萄不同倍性品种间杂交亲和性和后代染色体变异的研究[J].沈阳农业大学学报,2004,35(3):192–194.
[16] Johnston S A.The significance of genic balance to endosperm development in interspecific crosses[J]. Theoretical and Applied Genetics,1980,57(1):5–9.
[17] 钟声.鸭茅不同倍性杂交及后代发育特性的初步研究[J].西南农业学报,2006,19(6):1034–1038.
[18] 黄群策,吴福林,向茂成.异倍性水稻间异交结实特性的研究[J].湖南农业科学,1997(4):18–19.
[19] 马国平.不同倍数体桑杂交性能的研究[J].江苏蚕业,1999(1):59.
[20] 陈德灿,汤泽生,杨军.航天诱导凤仙花 SP3代子叶变化的研究[J].热带亚热带植物学报,2006,14(3):202–206.
[21] Hu J,Miller J F,Vick B A.Registration of a tricotyledon sunflower genetic stock[J].Crop Science,2007,46(6):2734– 2735.
[22] Sacita Kandya.A note on tricotyledonousseedlings of copper pod plant(Peltophorumferrugineum Benth.)[J]. Journal of Tropical Forestry,1988,4(4):339–400.
责任编辑:尹小红
英文编辑:梁 和
The crossability and progeny ploidyidentification of hybridization among the heteroploidincrape myrtle
Wang Hua, Chen Fang, Duan Lijun, Ye Yaomei*
(Key Laboratory of Horticultural Plant Biology, Ministry of Education/College of Horticulture and Forestry Science,Huazhong Agricultural University, Wuhan 430070, China)
Abstract:In the present study, we used H(2n), Z(3n), Z(4n ) and Y(4n) as the parents of hybridization to indicated the crossing-compatibility between them and identify the ploidy of offspring by cytological examination. The results suggested that there are obvious differences on fruiting and sprout among them. Capsule weight(0.22 g), seeds per capsule(40.1), thousand seed weight(1.79 g) and germination rate(36.4%) in the hybridization between Z(4n) and H(2n)were significantly higher than that of hybridization between H(2n) and Y(4n), which suggested the crossingcompatibility of 4n×2n was higher than the reciprocal cross. Most of lines in the progeny of H(2n)×Z(3n) were aneuploidy(80%)with a little diploid(11.8%); 50% lines in the progeny of cross Y(4n)×H(2n) were triploid, and 40% were aneuploidy. In the seedlings of cross H(2n)×Z(3n), 1 seedling was identified as 2n, 2 seedlings were aneuploidy; in the cross Y(4n)×H(2n), 6 seedlings were triploid, 3 seedlings were aneuploidy; in the cross H(2n)×Y(4n), 1 seedling as diploid, 1 seedling as triploid; in the cross Z(4n)×Z(4n), 1 seedling as tetraploid. Besides, tricotyledon mutant seedlings were founded in the cross of H(2n)×Z(3n), Y(4n)×H(2n) and H(2n)×Y(4n).
Keyword:crape myrtle; heteroploid; crossing-compatibility; ploidy identification
中图分类号:S685.99
文献标志码:A
文章编号:1007−1032(2016)03−0291−05
收稿日期:2015–08–14 修回日期:2016–03–15
基金项目:国家博士后基金项目(2014M550105);武汉市科技攻关项目(3013020705070350–4)
作者简介:王滑(1981—),男,重庆人,博士,讲师,主要从事种质资源收集与评价研究,near1981@mail.hzau.edu.cn;*通信作者,叶要妹,教授,主要从事园林植物育种研究,yymld@mail.hzau.edu.cn
