青藏高原5种害鼠vkorc1基因的测序分析
2016-07-16张同作苏建平慈海鑫李生庆李志宁林恭华
赵 芳,张同作,苏建平,慈海鑫,李生庆,李志宁,林恭华
(1.中国科学院西北高原生物研究所,青海 西宁 810001; 2.中国科学院大学,北京 100049;3.潍坊滨海开发区管委会,山东 潍坊 261108; 4.青海省畜牧兽医科学院,青海 西宁 810016)
青藏高原5种害鼠vkorc1基因的测序分析
赵 芳1,2,张同作1,苏建平1,慈海鑫3,李生庆4,李志宁4,林恭华1
(1.中国科学院西北高原生物研究所,青海 西宁 810001; 2.中国科学院大学,北京 100049;3.潍坊滨海开发区管委会,山东 潍坊 261108; 4.青海省畜牧兽医科学院,青海 西宁 810016)
摘要:维生素K环氧化物还原酶复合物1基因(vkorc1)的变异是导致鼠类对抗凝血杀鼠剂产生抗性的主要原因。本研究利用转录组测序的方法,分析青藏高原地区5种主要害鼠——高原鼢鼠(Eospalax baileyi)、高原鼠兔(Ochotona curzoniae)、长尾仓鼠(Cricetulus longicaudatus)、青海田鼠(Lasiopodomys fuscus)和喜马拉雅旱獭(Marmota himalayana)的vkorc1基因序列信息。同时,采集5个种群共54只高原鼢鼠,对vkorc1基因全序列进行测序分析。结果显示,从转录组组装结果中成功获得5种动物的vkorc1基因编码区全序列,其中青海田鼠、长尾仓鼠、高原鼢鼠vkorc1基因编码区长度为486 bp,喜马拉雅旱獭和高原鼠兔为492 bp;在DNA序列水平上,5种高原动物存在143个变异位点,与大鼠的序列相似性为80.1%~90.7%,而与小鼠的序列相似性为79.7%~89.8%。在氨基酸序列水平上,5种高原动物存在37个变异位点,大鼠的序列相似性为84.0%~92.0%,而与小鼠的序列相似性为85.9%~92.0%;未发现与已知抗凝血杀鼠剂抗性相关的氨基酸位点。对5个种群54只高原鼢鼠vkorc1基因全序列的测序分析显示,比对后的全长为1 808 bp,共检测到8个变异位点,其中两个为插入缺失位点,全部变异都发生在内含子区。本研究首次以基因序列为分析对象,可以为青藏高原地区的鼠害防治提供关键基础资料。
关键词:抗凝血杀鼠剂抗药性;vkorc1;序列变异;青藏高原;高原鼢鼠
鼠类是繁殖能力最强的哺乳动物,许多鼠类的过度繁殖给人类的健康和经济发展带来灾害性后果[1]。抗凝血杀鼠剂如华法林(warfarin)、敌鼠钠盐(diphacinone sodium salt)、溴敌隆(bromadiolone)等,具有安全、高效和适口性好等优点,是国内外鼠害控制中最常用的化学灭鼠剂[2]。然而,目前很多国家的鼠类对抗凝血杀鼠剂都产生了抗性,严重影响了灭鼠效力[2]。抗凝血杀鼠剂的杀鼠原理是与维生素K环氧化物还原酶(vitamin K epoxide reductase,VKOR)结合,阻止还原型维生素K 的生成,导致凝血功能障碍,同时损害毛细血管,使管壁渗透能力增加,最终造成害鼠大出血而亡[3]。编码VKOR的基因——维生素K环氧化物还原酶复合物1基因(vitamin K epoxide reductase complex subunit 1,vkorc1)的种内变异是导致鼠类对抗凝血杀鼠剂产生抗性的主要原因[4],而特定位点氨基酸的变异则可以用来监测鼠类种群的抗性水平。
尽管vkorc1基因在抗凝血杀鼠剂抗性方面起关键作用,然而迄今为止,对害鼠vkorc1基因的研究仅限于褐家鼠(Rattusnorvegicus)和小家鼠(Musmusculus)及其部分近缘物种,而对其它类群则很少涉及[5]。青藏高原是世界屋脊和中华水塔,也是鼠害发生的重灾区,青藏高原害鼠防治对我国生态系统安全具有重要意义[6]。本研究利用转录组测序的方法,分析青藏高原地区的高原鼢鼠(Eospalaxbaileyi)、高原鼠兔(Ochotonacurzoniae)、长尾仓鼠(Cricetuluslongicaudatus)、青海田鼠(Lasiopodomysfuscus)和喜马拉雅旱獭(Marmotahimalayana)5种主要害鼠[7]的vkorc1基因序列信息。同时,对高原鼢鼠5个种群的54只个体,进行vkorc1基因全序列的测序与分析,本研究首次以抗凝血杀鼠剂抗性基因vkorc1为分析对象,旨在为青藏高原地区的鼠害防治提供关键基础资料。
1材料与方法
1.1样品采集
高原鼢鼠和高原鼠兔采自湟中县拉鸡山,长尾仓鼠采自大通县向化乡,青海田鼠采自同德县城郊,喜马拉雅旱獭采自格尔木市西大滩。所有上述动物活体带回实验室,处死后迅速切取脑组织,置于液氮中固定和临时保存,继而转移至-80 ℃冰箱中长期保存。用死捕法采集祁连县八宝镇、祁连县默勒镇、天峻县新源镇、泽库县巴滩牧场、若尔盖县阿西乡,每个种群采集9~12只鼢鼠,现场解剖后采集腿部肌肉,置于无水乙醇中固定保存。
1.2转录组测序分析
害鼠的脑组织样品用干冰运输至百迈客生物科技有限公司,按照Illumina公司提供的标准流程进行样本制备和上机文库构建,最后用IlluminaHiSeqTM 2000进行高通量测序,指标为100 bp(双向)。对测序得到的原始reads用FASTX-Toolkit软件[8]进行数据评估、去除接头和低质量区域,得到用于生物信息学分析的clean reads,并统计序列基本信息。用Trinity软件[9]对每个物种的clean reads进行从头组装,组装后的序列用CD-HIT-EST[10]进一步去重,最终得到可用的unigene序列。BLAST+软件包中的makeblastdb程序对每个物种的unigene序列文件构建本地blast库,从GenBank中下载大鼠(NM_203335.2)和小鼠(BC031732.1)vkorc1基因的编码区序列,用BLAST+软件包中的blastn程序调取各物种包含vkorc1基因的unigene序列。
将这些unigene序列及上述大鼠、小鼠vkorc1基因的编码区序列合并至一个文本文档,用MEGA软件进行序列比对,同时,翻译成氨基酸序列,最终得到所有物种的vkorc1基因编码区DNA序列及氨基酸序列。用DNasp软件[11]分析DNA序列的变异位点,而氨基酸水平的变异位点直接在MEGA[12]中手工统计。用DNAstar软件包(http://www.dnastar.com/)中的Megalign软件分析DNA序列和氨基酸序列的相似性百分比。此外,收集现有大鼠、小鼠抗药性突变位点资料,在MEGA软件中判断这5个物种是否存在这类氨基酸位点。
1.3高原鼢鼠vkorc1基因测序分析
采用动物组织基因组DNA提取试剂盒(Qiagen,Germany)提取基因组DNA。根据高原鼢鼠基因组序列(待发表)设计扩增vkorc1基因两个片段的引物:vkorc1-1F 5′-TCAAAGAATACCAGACGA-3′/vkorc1-1R 5′-GGAATACACGAAACCATA-3′;vkorc1-2F 5′-GAGAGGGTAGAGTCGGTGA-3′/ vkorc1-2R 5′-GGTTGGAAGAAGGAATTAG-3′。PCR扩增使用40 μL反应体系,包括:40~60 ng的基因组DNA,0.6 mmol·L-1dNTPs,0.2 μmol·L-1的正/反向引物,1 U Taq酶,以及DNA buffer和去离子水的混合反应缓冲液。PCR反应程序:95 ℃预变性5 min;94 ℃变性45 s,55 ℃退火45 s,72 ℃延伸75 s,循环35次;最后72 ℃延伸7 min。PCR产物用CASpure PCR Purification Kit(Casarray,Shanghai,China)纯化,用PCR扩增引物进行Sanger法双向测序。
用MEGA 5中内嵌的CLUSTAL W[13]程序比对测序得到的序列,辅以人工校对,外显子区和内含子区边界参考小鼠的GenBank序列说明(GQ905710)确定。由于少数个体存在多个杂合子位点(简并碱基),无法直接读出其单倍基因型,本研究用二倍体基因型(diplotype)替代,即,两两二倍体序列之间所有座位上碱基(包括简并碱基对)相同则视为同一个diplotype。此外,高原鼢鼠vkorc1基因内含子区有插入缺失位点,而国际上通用的简并碱基命名规则(IUPAC Ambiguity Codes)中并无插入缺失杂合子简并性(如T/-)的定义,因此现有软件无法处理这类位点,将这类位点用小写字母表示(a=A/-,c=C/-,g=G/-,t=T/-),直接用Editplus搜索(区分大小写模式)法确定基因型组成,而序列变异位点信息直接在MEGA软件中人工读取。
2结果与分析
2.1种间vkorc1基因变异情况
转录组测序得到大量的碱基序列,每个物种都得到4千万条Reads,每个物种测得的总碱基数(bp)都在40亿以上。经组装拼接,每个物种都得到9万条以上Unigene序列,N50长度在1 500 bp以上,测序效果比较理想(表1)。经Blast搜索和MEGA比对,得到5个物种vkorc1基因编码区的完整序列,其中青海田鼠、长尾仓鼠、高原鼢鼠为486 bp(与大鼠、小鼠的序列等长),而喜马拉雅旱獭和高原鼠兔为492 bp。在DNA序列水平上,5种动物比对后存在143个变异位点(包括6个插入缺失位点)。5种动物的DNA序列与大鼠的序列相似性为80.1%~90.7%,而与小鼠的序列相似性为79.7%~89.8%(表2)。
在氨基酸序列水平上,5种动物存在37个变异位点(包括两个插入缺失位点)。5种动物的氨基酸序列与大鼠的序列相似性为84.0%~90.8%,而与小鼠的序列相似性为85.9%~92.0%(表2),具体位点变异分布情况见图1。根据文献报道[2,4,14-19],大鼠属(Arg33Pro,Arg35Pro,Ser56Pro,Phe63Cys,Leu120Gln,Leu128Gln/Ser,Tyr139Cys/Ser/Phe)和小鼠属(Arg12Try,Ala26Ser,Ala48Thr,Arg58Gly,Arg61Leu,Leu128Ser,Tyr139Cys)都分别发现7个氨基酸位点变异导致其对抗凝血杀鼠剂的抗性增加。经核对,5个物种在大鼠属抗性突变位点上都完全保守(与野生型相同)。在小鼠属抗性突变位点上,有5个位点保守(位点12、26、58、128和139)。在48号位点,青海田鼠为丝氨酸(Ser)而其余物种为丙氨酸(Ala);而在61号位点,长尾仓鼠和高原鼢鼠分别为赖氨酸(Lys)和谷氨酰胺(Gln),而其余物种都是精氨酸(Arg)(图1);然而,这些位点变异都与小鼠抗性氨基酸不同。
2.2高原鼢鼠vkorc1基因
对54只高原鼢鼠个体进行测序,两对引物都成功扩增,经序列校对和拼接,得到54条vkorc1基因的全序列,比对后的全长为1 808 bp(包括插入缺失位点)。其中1~174、175~929、930~1 040、1 041~1 607、1 608~1 808分别为外显子exon1、intron1、exon2、intron2、exon3。共检测到8个变异位点,其中两个为插入缺失位点。所有3个外显子区都没有变异发生,而intron1和intron2分别存在6个和2个变异位点(表3)

表1 青藏高原5种害鼠转录组测序样品信息和测序结果
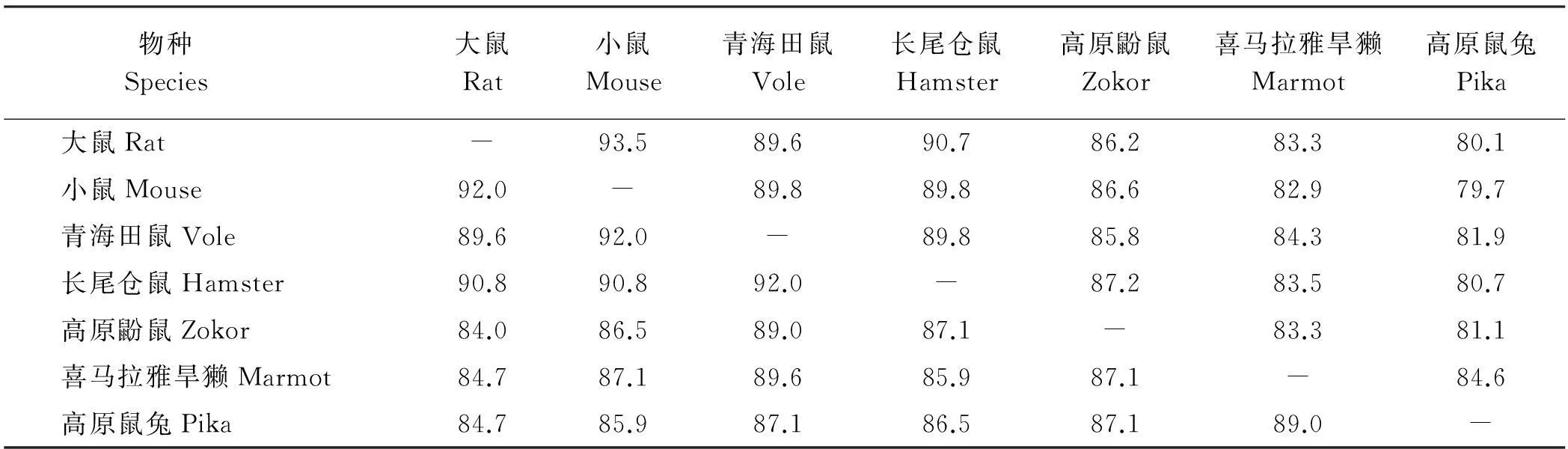
表2 青藏高原5种害鼠及大鼠、小鼠的vkorc1基因编码区DNA序列(上三角)和氨基酸序列(下三角)相似度(%)
基因型分析检测到14个二倍体基因型,其中5个为纯合子(表3)。基因型D01、D06、D09频率超过10%,其余频率较低。祁连县八宝镇遗传多样性最低,而其它区域的个体间都有基因变异。D01、D06、D11在两个或以上种群出现,其余都是各种群独有基因型(表4)。
3讨论
vkorc1基因在抗凝血杀鼠剂抗性方面起关键作用,对其进行遗传变异分析,有助于杀鼠剂的科学合理使用,降低灭鼠成本和延长杀鼠剂更新换代年限[4]。然而长期以来,对大、小鼠之外鼠类的vkorc1鲜见报道,可能的原因是,多数害鼠缺乏基因组数据,难以设计有效的引物进行PCR扩增和测序。近年来,转录组测序分析技术发展迅速,测序成本也大幅下降,使得研究人员通过从头测序的方法(de novo)得到非模式物种的基因序列成为可能。本研究通过单只动物脑组织的转录组测序,即成功得到vkorc1基因的编码区完整序列,足见这种方法在此基因研究方面的优势。值得一提的是,转录组测序产生的数据量极大,除了单份样品测序外,还可将来源于多只动物的样品合并后测序[20],通过SNP鉴定的方法,提高抗性个体发现的几率。

图1 青藏高原5种害鼠及大鼠、小鼠的vkorc1基因氨基酸序列变异位点分布

基因型Genotype序列位点Siteposition11124567228487243614273074类型Type频率FrequencyD01CCAGGG-GHo14D02CCaRGR-GHe2D03CC-GGGtGHe2D04CC-GGGTGHo1D05CCARGG-GHe1D06CC-GGG-GHo7D07CCaGGR-GHe1D08CTAGRG-AHe2D09CTAGGG-RHe2D10CTAGGG-AHo2D11CTAGGG-GHo9D12CTAGRG-RHe1D13YTAGGG-GHe5D14CCaGGG-GHe5
注:小写字母表示插入缺失杂合子,例a=A/-;Ho,纯合子;He,杂合子。
Note: The lower case letters show the indel heterozygotes, e.g. a=A/-; Ho, homozygote; He, heterozygote.
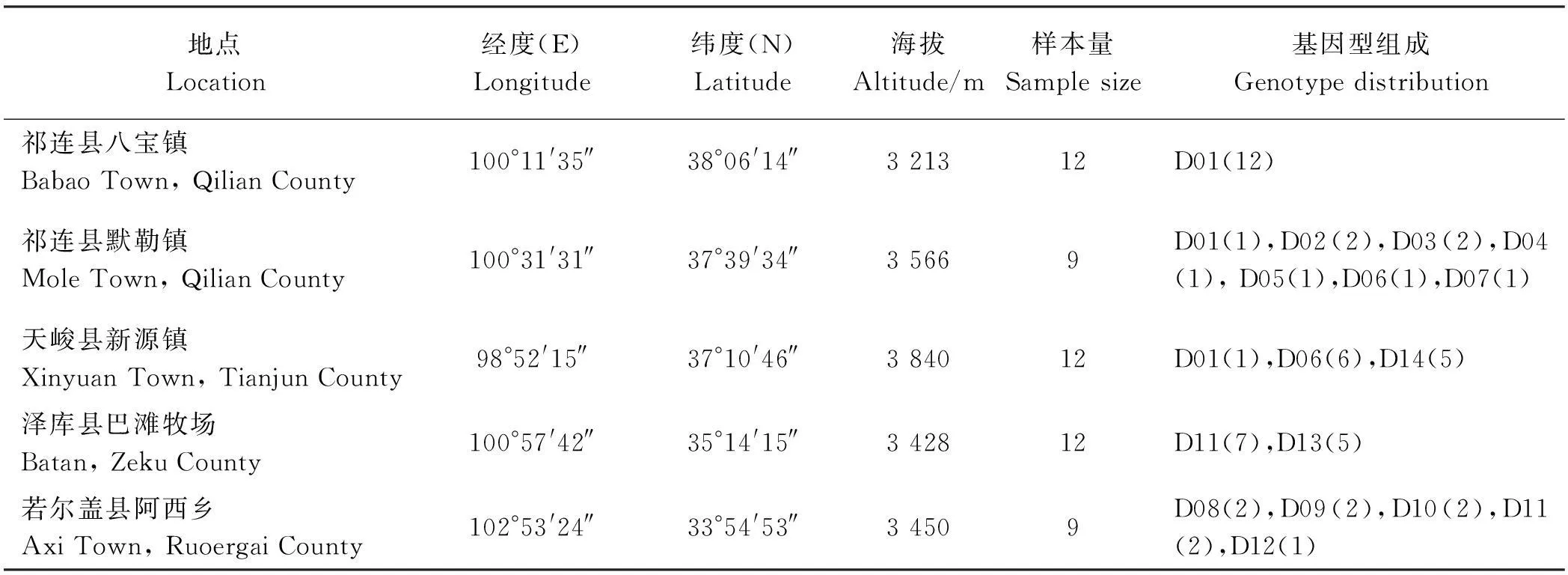
表4 高原鼢鼠种群采样信息和基因型分布
本研究所涉及的动物都是青藏高原及周边地区重要的害鼠类群。其中,高原鼢鼠和高原鼠兔分别是青藏高原高寒草甸生态系统的主要地下和地上害鼠;长尾仓鼠是该地区农田的优势害鼠;喜马拉雅旱獭和青海田鼠对草地的危害相对较轻,然而,两者分别是青藏高原“喜马拉雅旱獭鼠疫自然疫源地”和“青海田鼠鼠疫自然疫源地”的关键疫源动物[21],其种群增长对该地区的鼠疫防控有关键性影响。本研究显示,这5种动物的vkorc1基因序列之间差异很大,例如高原鼢鼠和高原鼠兔之间在氨基酸序列上有近13%(21个氨基酸)的差异。与此同时,不同物种对同一种杀鼠剂的敏感性可能有很大差别[22],例如高原鼢鼠和高原鼠兔对溴敌隆的半致死剂量(LD50)分别为1.31和0.43 mg·kg-1,差异可达3倍之多[23]。有趣的是,高原鼢鼠和高原鼠兔之间的氨基酸差异位点与已知的大鼠或小鼠的抗药性位点并不相同,可见还有许多与抗凝血剂抗性相关的座位有待发掘分析。
本研究涉及的5种害鼠中,高原鼢鼠是唯一的地下鼠。由于鼢鼠类动物的杀鼠剂毒饵投放和毒杀过程都在地下洞道中完成,基本不会发生非靶动物的误伤问题,因此抗凝血剂类药物(如鼢鼠灵)长期在这类动物中频繁使用[24-25]。本研究对高原鼢鼠vkorc1基因的测序结果显示,所有5个种群的54个个体的编码区序列完全一致,同时,未见有与大鼠、小鼠抗性突变相同的氨基酸位点。据此可以推测,高原鼢鼠vkorc1基因编码区非常保守,产生抗药性的可能性较小。有趣的是,高原鼢鼠vkorc1基因的内含子区有丰富的变异。相关研究显示,高原鼢鼠及其它害鼠对抗凝血杀鼠剂敏感性的个体差异较高[23,26]。尽管内含子区不编码氨基酸序列,然而其在相关基因的表达调控方面起重要作用[27]。据此推测,高原鼢鼠vkorc1基因的内含子区序列的变异可能是导致其对灭鼠剂抗性个体差异的重要因素。本研究还显示,高原鼢鼠不同种群之间只有少数几个基因型共享,同时,种群之间遗传多样性差别很大,这与地下鼠较弱的迁移能力(基因流较小)有关。这就意味着,不同种群的高原鼢鼠在杀鼠剂敏感性方面可能有较大差别,对这方面开展深入、细致的研究,将有利于科学合理地规划灭鼠方案和用药标准。
参考文献References:
[1]周延山,花立民,纪维红,楚彬,刘丽.鼢鼠亚科繁殖特性研究进展.草业科学,2015,32(6):1010-1016.
Zhou Y S,Hua L M,Ji W H,Chu B,Liu L.A review about reproductive characteristics of zokors.Pratacultural Science,2015,32(6):1010-1016.(in Chinese)
[2]Buckle A P,Smith R H.Rodent Pests and Their Control.London:CABI,2015.
[3]Watt B E,Proudfoot A T,Bradberry S M,Vale J A.Anticoagulant rodenticides.Toxicological Reviews,2005,24(4):259-269.
[4]Pelz H J,Rost S,Muller E,Esther A,Ulrich R G,Muller C R.Distribution and frequency ofVKORC1 sequence variants conferring resistance to anticoagulants inMusmusculus.Pest Management Science,2012,68:254-259.
[5]Gryseels S,Leirs H,Makundi R,Goüy de Bellocq J.Polymorphism invkorc1 gene of natal multimammatemice,Mastomysnatalensis,in Tanzania.Journal of Heredity,2015,106(5):637-643.
[6]张兴禄,李广.高原鼠兔和高原鼢鼠在高寒草甸生态系统的作用.草业科学,2015,32(5):816-822.
Zhang X L,Li G.Effects of rodents activities on grazingland and ecosystem in alpine meadow.Pratacultural Science,2015,32(5):816-822.(in Chinese)
[7]郭正刚,王倩,陈鹤.我国天然草地鼠害防控中的问题与对策.草业科学,2014,31(1):168-172.
Guo Z G,Wang Q,Chen H.Issues and suggestions for rodent control of the natural grassland in China.Pratacultural Science,2014,31(1):168-172.(in Chinese)
[8]Pearson W R,Wood T,Zhang Z,Miller W.Comparison of DNA sequences with protein sequences.Genomics,1997,46(1):24-36.
[9]Grabherr M G,Haas B J,Yassour M,Levin J Z,Thompson D A,Amit I,Adiconis X,Fan L,Raychowdhury R,Zeng Q D,Chen Z,Mauceli E,Hacohen N,Gnirke A,Rhind N,di Palma F,Birren B W,Nusbaum C,Lindblad-Toh K,Friedman N,Regev A.Full-length transcriptome assembly from RNA-Seq data without a reference genome.Nature Biotechnology,2011,29(7):644-652.
[10]Li W Z,Godzik A.Cd-hit:A fast program for clustering and comparing large sets of protein or nucleotide sequences.Bioinformatics,2006,22(13):1658-1659.
[11]Librado P,Rozas J.DnaSP v5:A software for comprehensive analysis of DNA polymorphism data.Bioinformatics,2009,25:1451-1452.
[12]Tamura K,Peterson D,Peterson N,Stecher G,Nei M,Kumar S.MEGA5:Molecular evolutionary genetics analysis using maximum likelihood,evolutionary distance,and maximum parsimony methods.Molecular Biology and Evolution,2011,28:2731-2739.
[13]Thompson J D,Gibson T J,Plewniak F,Jeanmougin F,Higgins D G.The CLUSTAL_X windows interface:Flexible strategies for multiple sequence alignment aided by quality analysis tools.Nucleotide Acids Research,1997,25:4876-4882.
[14]Pelz H J,Rost S,Hünerberg M,Fregin A,Heiberg A C,Baert K,MacNicoll A D,Prescott C V,Walker A S,Oldenburg J,Muller C R.The genetic basis of resistance to anticoagulants in rodents.Genetics,2005,170:1839-1847.
[15]Rost S,Pelz H J,Menzel S,MacNicoll A D,Leon V,Song K J,Jakel T,Oldenburg J,Muller C R.Novel mutations in theVKORC1 gene of wild rats and mice——A response to 50 years of selection pressure by warfarin?BMC Genetics,2009,10:4.
[16]Grandemange A,Lasseur R,Longin-Sauvageon C,Benoit E,Berny P.Distribution ofVKORC1 single nucleotide polymorphism in wildRattusnorvegicusin France.Pest Management Science,2009,66:270-276.
[17]Song Y,Endepols S,Klemann N,Richter D,Matuschka F,Shih C,Nachman M,Kohn M H.Adaptive introgression of anticoagulant rodent poison resistance by hybridization between Old World mice.Current Biology,2011,21(15):1296-1301.
[18]Tanaka K D,Kawai Y K,Ikenaka Y,Harunari T,Tanikawa T,Fujita S,Ishizuka M.A novel mutation inVKORC1 and its effect on enzymatic activity in Japanese warfarin-resistant rats.Journal of Veterinary Medical Science,2013,75:135-139.
[19]Muller E,Keller A,Fregin A,Muller C R,Rost S.Confirmation of warfarin resistance of naturally occurringVKORC1 variants by coexpression with coagulation factor Ⅸ and in silico protein modeling.BMC Genetics,2014,15:17.
[20]Lemay M A,Henry P,Lamb C T,Robson K M,Russello M A.Novel genomic resources for a climate change sensitive mammal:Characterization of the American pikatranscriptome.BMC Genomics,2013,14:311.
[21]贺雄,王虎.现代鼠疫概论.北京:科学出版社,2010.
[22]李道中.第二代抗凝血杀鼠剂“溴特隆”的生物学试验评价.中国鼠类防制杂志,1987,3(3):140-144.
Li D Z.Biological assessment of the second generation anticoagulant rodenticides bromadiolone.Chinese Journal of Rodent Control,1987,3(3):140-144.(in Chinese)
[23]樊乃昌,景增春,王权业,周文扬.溴敌隆防治高原鼠兔和高原鼢鼠的研究.兽类学报,1986,6(3):211-217.
Fan N C,Jing Z C,Wang Q Y,Zhou W Y.Studies on bromadiolone against the pika and the zokor.Acta Theriologica Sinica,1986,6(3):211-217.(in Chinese)
[24]杨廷勇,周俗,阮芳泽,唐川江,晏学举.鼢鼠灵防治高原鼢鼠的试验研究.四川草原,2005(11):35-36.
[25]唐俊伟,张明,马志贵.3种药剂防治高原鼢鼠的药效研究.现代农业科技,2013(24):144,152.
[26]常文英,郭学林,宁振东,王庭林,邹波,杨新根,侯玉,朱文雅.长尾仓鼠对溴敌隆的抗药性研究.中国植保导刊,2011,31(5):38-39.
[27]Chorev M,Carmel L.The function of introns.Frontiers in Genetics,2012,3:1-15.
(责任编辑武艳培)
Variation in vkorc1 gene of five rodent species endemic to the Qinghai-Tibet Plateau
Zhao Fang1,2, Zhang Tong-zuo1, Su Jian-ping1, Ci Hai-xin3,Li Sheng-qing4, Li Zhi-ning4, Lin Gong-hua1
(1.Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810001, China;2.University of Chinese Academy of Sciences, Beijing 100049, China;3.Management Committee of Weifang Binhai Economic Development Zone, Weifang 261108, China;4.Qinghai Academy of Animal and Veterinary Sciences, Xining 810016, China)
Abstract:The variations of vitamin K epoxide reductase complex subunit 1 (vkorc1) gene play a key role in resistance of rodents to anticoagulant rodenticides. In this study, the transcriptomic methods were applied to analyze the sequence information of five rodents plateau zokor (Eospalax baileyi), plateau pika (Ochotona curzoniae), lesser long-tailed hamster (Cricetulus longicaudatus), Qinghai vole (Lasiopodomys fuscus), and Himalayan marmot (Marmota himalayana), which are endemic to the Qinghai-Tibet Plateau (QTP). We also sampled 54 plateau zokors from five populations and analyzed the variations within the complete vkorc1 gene (including introns). From transcriptome assemblies, we successfully obtained the complete coding sequences of vkorc1 gene of five QTP animals. Of which, the length of vkorc1 CDS in vole, hamster, and zokor was 486 bp, while it was 492 bp in marmot and pika. At the DNA sequence level, there were 143 variable sites among the five QTP animals, the sequence similarities between the five QTP animals and rat (Rattus norvegicus) were 80.1%~90.7%, while between the five QTP animals and mouse (Mus musculus) were 79.7~89.8%. At the amino acid sequence level, there were 37 variable sites among the five QTP animals, the sequence similarities between the five QTP animals and rat (Rattus norvegicus) were 84.0%~92.0%, while between the five QTP animals and mouse were 85.9%~92.0%; no amino acid residue was identical with which involved in anticoagulant resistance in Rattus and Mus. We obtained the complete vkorc1 gene (including introns) sequences of the 54 zokors, based on the 1 808 bp alignment we detected 8 variable sites (including 2 indel sites), all of them were in the intron region. This is the first vkorc1 sequence data from rodent species endemic to the QTP and these results will provide important scientific foundations for rodent control in the QTP regions.
Key words:resistance to anticoagulant rodenticides; vkorc1; sequence variation; Qinghai-Tibet Plateau; plateau zokor
DOI:10.11829/j.issn.1001-0629.2015-0644
*收稿日期:2015-11-17接受日期:2016-02-24
基金项目:青海省科技支撑计划项目——青藏高原草地鼠害综合防控技术集成与示范(2014-NS-113);青海省科技项目——贵南县退化草地恢复治理及生态畜牧业关键技术集成与应用(2014-NS-118)
通信作者:林恭华(1983-),男,浙江江山人,副研究员,博士,主要从事动物生态学研究。E-mail:lingonghua@163.com
中图分类号:S812.6
文献标志码:A
文章编号:1001-0629(2016)6-1206-07*
Corresponding author:Lin Gong-huaE-mail:lingonghua@163.com
赵芳,张同作,苏建平,慈海鑫,李生庆,李志宁.林恭华.青藏高原5种害鼠vkorc1基因的测序分析.草业科学,2016,33(6):1206-1212.
Zhao F,Zhang T Z,Su J P,Ci H X,Li S Q,Li Z N,Lin G H.Variation invkorc1 gene of five rodent species endemic to the Qinghai-Tibet Plateau.Pratacultural Science,2016,33(6):1206-1212.
动物
生产层
第一作者:赵芳(1987-),女,湖南澧县人,在读博士生,主要从事分子生态学研究。E-mail:zhaofang@nwipb.cas.cn
