3个披碱草属牧草对低温胁迫的生理响应及苗期抗寒性评价
2016-07-16张尚雄尼玛平措徐雅梅苗彦军包赛很那张卫红
张尚雄,尼玛平措,徐雅梅,苗彦军,包赛很那,张卫红
(1.西藏大学农牧学院,西藏 林芝 860000; 2.西藏自治区农牧科学院畜牧兽医科学研究所,西藏 拉萨 850000)
3个披碱草属牧草对低温胁迫的生理响应及苗期抗寒性评价
张尚雄1,尼玛平措2,徐雅梅1,苗彦军1,包赛很那1,张卫红1
(1.西藏大学农牧学院,西藏 林芝 860000; 2.西藏自治区农牧科学院畜牧兽医科学研究所,西藏 拉萨 850000)
摘要:探讨低温胁迫下西藏野生披碱草(Elymus)苗期抗寒性及其生理变化,可为优良牧草选育提供理论依据。以披碱草属3个牧草为试验材料,出苗21 d后在人工气候箱内进行低温胁迫,设18、10、5和0 ℃共4个低温处理,其中以18 ℃为对照。结果表明,随着低温胁迫的加剧,相对膜透性,同德老芒麦(E. sibiricus cv. Tong De)的增幅最大,巴青垂穗披碱草(E. nutans)增幅最小。叶绿素含量,巴青垂穗披碱草先升高后降低,同德老芒麦与那曲垂穗披碱草显著升高(P<0.05)。巴青垂穗披碱草可溶性蛋白(SP)含量增加幅度最小,为对照的2.59倍;同德老芒麦与那曲垂穗披碱草SP含量显著上升(P<0.05),均为对照的5.31倍。巴青垂穗披碱草与同德老芒麦的可溶性糖(SS)含量先升高后降低,而那曲垂穗披碱草SS含量显著增大(P<0.05),为对照的2.14倍。同德老芒麦的丙二醛(MDA)含量增幅最大,巴青垂穗披碱草MDA含量0 ℃较5 ℃时显著降低(P<0.05)。3种供试材料游离脯氨酸(Pro)含量均升高,同德老芒麦增幅最大,为对照的2.58倍;巴青垂穗披碱草最小,为对照的1.44倍。巴青垂穗披碱草超氧化物歧化酶(SOD)活性先增强后减弱,那曲垂穗披碱草SOD活性显著增强(P<0.05),为对照的1.65倍。那曲垂穗披碱草过氧化物酶(POD)活性上升幅度最大,为对照的4.13倍;同德老芒麦最小,为对照的2.91倍。隶属函数法综合评价表明,3种野生披碱草的抗寒性依次为巴青县野生垂穗披碱草>同德老芒麦>那曲县野生垂穗披碱草。
关键词:披碱草;低温胁迫;生理特性;抗寒性评价;西藏
西藏是青藏高原的主要组成部分,平均海拔4 000 m以上,气温年较差小,日较差大,夏季气温不高,冬季干燥少雨,具有独特的高原气候[1]。温度是西藏植物正常生长发育的关键因子之一,其影响范围包括植物的新陈代谢、酶促反应、休眠等[2]。西藏植物往往会遭受极端气温从而影响其生长发育。目前低温是制约西藏披碱草(Elymusspp.)生长发育的一个关键因素。因此,如何提高披碱草的抗寒性,最大限度地提高披碱草的经济价值成为西藏草产业发展面临的一大难题。
披碱草属植物在西藏分布极广,甚至在5 200 m以上的喜马拉雅山区也有生长[3]。西藏披碱草属牧草种类极为丰富,但原始生境不同的披碱草,其抗寒性能也有差异。西藏当雄县野生垂穗披碱草(E.nutans)和甘南垂穗披碱草在低温胁迫下,两种垂穗披碱草的细胞膜受损伤程度与抗氧化酶活性负相关,当雄县野生垂穗披碱草的抗寒能力较强,由于其抗氧化酶活性在冷胁迫时显著增强,减轻了细胞膜系统受损伤的程度[4]。早熟禾(Poa)在低温胁迫下,可溶性蛋白、可溶性糖、抗氧化酶活性、海拔与其耐寒能力正相关[5]。因为植物体内膜的透性、可溶性物质、丙二醛、叶绿素以及抗氧化酶等指标是评价植物抗寒性强弱的优良指标,所以在前人的研究基础上,本研究以同德老芒麦(E.sibiricus)、巴青县披碱草、那曲县披碱草为材料,其中以同德老芒麦为对照,通过低温胁迫观察材料的相对电导率,叶绿素、渗透调节物质以及抗氧化酶的活性等生理指标,来比较评价西藏当地野生披碱草的抗寒性能。本研究对西藏本地采集的野生披碱草进行抗寒性强弱的相关性分析,指出西藏本地野生披碱草的抗寒性强弱,旨为西藏披碱草抗寒种质筛选和抗寒新品种选育提供理论依据。
1材料与方法
1.1试验材料
同德老芒麦由青海省畜牧兽医科学院草原所提供,其它两份材料分别采集于西藏的巴青县和那曲县(表1)。
1.2试验方法
1.2.1试验设计低温处理设有3个处理水平(10、5、0 ℃),并以室温18 ℃为对照(CK)。试验采用盆栽供试材料,播种量为15 g·m-2,花盆直径为20 cm、深15 cm,混合栽培基质为农田土∶羊粪=9∶1(质量比),花盆内装等量混合栽培基质[5]。2014年11月22日将全部盆栽材料置于25 ℃(白天)/18 ℃(晚上)的人工物候箱内培养,2014年12月1日全部出苗,经过21 d的培养,2014年12月22日开始进行低温胁迫处理。处理时对照组直接放入18 ℃培养箱中培养,其它处理组先在15 ℃的培养箱中预冷24 h,然后,每隔2 h降5 ℃,直至试验处理温度,再持续低温胁迫8 h,试验处理结束后,立刻剪取叶片,将其混匀,根据试验指标中的鲜样称重,锡纸包裹,并用液氮迅速冷冻,放入-84 ℃低温冰箱保存,备用[5]。
1.2.2试验测定指标相对电导率、可溶性糖(SS)、可溶性蛋白(SP)的测定以《植物生理学实验技术》为指导[6]。其中细胞膜透性测量方法如下:将去离子水洗净的材料叶片剪成1cm长的条段120段,分为6组,对照与试验组各3个重复,相对电导率=(处理电导率-对照电导率)/(煮沸电导率-对照电导率)×100%。可溶性糖测定时取样均在0.10~0.12 g,3次重复,可溶性糖含量=C×V×n/1 000×a×W,C为标准方程求得糖量(μg),a为吸取样品液体积(mL),V为提取液量(mL),n为稀释倍数,W为组织重量(g)。可溶性蛋白测定时取样均在0.25~0.30 g,3次重复,样品中蛋白质含量=C×VT/(VS×WF×1 000)。式中:C为查标准曲线值(μg),VT为提取液总体积(mL),VS为测定时加样量(mL),WF为样品鲜重(g)。

表1 供试的3个牧草
注:下文中图和表中材料名称均以编号表示。
Note: The codes of material names will be used in the following figures and tables.
丙二醛(MDA)采用硫代巴比妥酸(TBA)法测定[7]。丙二醛测定时取样均在1.00~1.10 g,3次重复。
叶绿素、游离脯氨酸(Pro)、超氧化物歧化酶(SOD)、过氧化物酶(POD)参照《植物生理学实验教程》测定[8]。叶绿素测定时采样量均在2.00~2.10 g,3次重复,CT=20.29A645+8.05A663,叶绿体色素含量=CT×V×n/W。式中:CT为色素的质量浓度(mg·L-1),A645和A663分别为645和663 nm处的吸光度,V为提取液体积(L),n为稀释倍数,W为样品鲜重(g)。游离脯氨酸测定时采样量均在2.00~2.10 g,3次重复,脯氨酸含量=c×VT/W×V1。式中:c为由标准曲线上查得的脯氨酸含量,VT为提取液总体积(mL),W为样品鲜重(g),V1为测定系统中提取液的加入量(mL)。超氧化物歧化酶活性U·(g·min)-1测定时取样量均在0.50~0.51 g,3次重复,NBT光还原的抑制率=1-(A1-A2)/A1×100%,超氧化物歧化酶=1 000×60×V/B×W×t。式中:A1为对照杯在560 nm的吸光度,A2为加酶杯在560 nm的吸光度,B为一个酶活单位的酶液量(μL),V为酶提取液总量(mL),t为反应时间(min),W为样品鲜重(g)。过氧化物酶活性U·(g·min)-1测定时采样量均在1.00~1.10 g,3次重复,过氧化物酶活性=ΔA470×V/0.01×W×Vt×t。式中:ΔA470为反应时间内吸光度的变化,W为材料鲜重(g),V为酶提取液总量(mL),Vt为反应系统中加入酶液量(mL),t为反应时间(min)。其中,超氧化物歧化酶活性与过氧化物酶活性的单位中的U表示一个酶活性单位,即反应体系NBT的光化还原率为对照的50%时所用的酶液量。
1.3供试材料苗期抗寒性综合评价
1.3.1抗寒性评价方法试验材料的抗寒性评价采用隶属函数法[9-10]。供试牧草各指标的隶属函数值:
μ(Xj)=(Xj-Xmin)/(Xmax-Xmin).
式中:Xj表示第j个指标值,Xmin表示第j个指标的最小值,Xmax表示第j个指标的最大值,如某一指标与抗寒性呈负相关,可用反隶属函数计算其抗寒隶属函数值[9]。
μ(Xj)=(Xmax-Xj)/(Xmax-Xmin).
标准差系数Vj:

各指标的权重Wj:

式中:Wj表示第j个指标的权重[9]。
综合评价值:

式中:D表示各材料抗寒性综合评价值,D值越大表明抗寒性越强[9]。
1.3.2抗寒性评价分级标准植物综合抗寒性D值可以分为3级,Ⅰ级:D值0.7以上,为抗寒性强;Ⅱ级:D值在0.5~0.7,抗寒性中等; Ⅲ级:D值在0.5以下,抗寒性较弱[5,9]。
1.4数据分析
统计分析采用SPSS 19.0,采用DUNCAN法多重比较;Excel 2013制作图表。
2结果与分析
2.1低温胁迫对披碱草叶片膜透性的影响
3种供试牧草随低温胁迫加剧,相对膜透性整体呈增加的趋势,但变化规律不尽相同(图1)。10 ℃时,所有供试牧草的相对膜透性显著增大,显著高于对照(P<0.05)。其中TD相对膜透性上升最快,约为对照的5.71倍。BQ相对膜透性上升最慢,约为对照的2.37倍。温度降至5 ℃时,所有供试材料的相对膜透性仍与对照差异显著,其中TD与NQ的相对膜透性较10 ℃处理有所减小,而BQ的相对膜透性显著升高,是对照的8.18倍。温度降至0 ℃时,TD与那NQ的相对膜透性较5 ℃时显著升高(P<0.05),并且TD相对膜透性高于10 ℃处理组,达到所有处理水平的最大值,约为对照的7.52倍,受到的伤害也最大。BQ相对膜透性与10 ℃的处理比较显著减小,受到的伤害也最小。
十一点钟的时候,敦礼已经坐在他上次坐的那个位子了。他记得她当时对服务生说的是老位子。他也要把这个位子变成属于自己的老位子吗?很快,他又觉得自己这个想法很荒谬,很可笑。点了一杯蓝山后,他就闭上眼睛靠在沙发靠背上。

图1 3种供试牧草叶片相对膜透性的变化
注:不同小写字母同一供试材料不同温度处理间差异显著(P<0.05)。下同。
Note:Different lower case letters for the same germplasm indicate significant difference among different temperatures at 0.05 level. The same below.
2.2低温胁迫对披碱草叶绿素含量的影响
供试材料叶绿素的含量随低温胁迫加剧,叶绿素含量均呈上升趋势(图2)。10 ℃处理下,除BQ叶绿素含量与对照显著差异外(P<0.05),其它供试牧草的叶绿素含量与对照差异均不显著(P>0.05)。5 ℃处理下,各供试材料叶绿素含量与对照显著差异。其中NQ叶绿素含量上升最快,为对照的1.86倍;TD叶绿素含量上升的最慢,为对照的1.46倍。0 ℃处理下,各供试材料叶绿素含量与对照显著差异(P<0.05),其中BQ叶绿素含量低于5和10 ℃时的含量,但显著低于5 ℃处理下的叶绿素含量,与10 ℃下的叶绿素含量差异不显著(P>0.05)。NQ叶绿素含量上升最快,为对照的2.13倍,BQ叶绿素含量上升最慢,为对照的1.32倍。

图2 3种供试牧草叶片叶绿素含量的变化
2.3低温胁迫对披碱草叶片可溶性蛋白(SP)含量的影响
各供试材料随低温胁迫的加剧,可溶性蛋白含量整体呈上升趋势(图3)。10 ℃处理下,除NQ以外,其余供试材料SP含量与对照显著差异(P<0.05)。5℃处理下,所有供试材料可溶性蛋白含量显著高于对照(P<0.05);其中NQ的SP含量上升最快,为对照的2.59倍。0 ℃处理下,NQ的SP含量增加最多,为对照的5.31倍;BQ的SP含量增加最少,是对照的1.68倍。

图3 3种供试牧草叶片可溶性蛋白(SP)含量的变化
2.4低温胁迫对披碱草叶片可溶性糖(SS)含量的影响
3个材料随低温胁迫加剧,SS含量均呈升高趋势(图4)。10 ℃处理下,3个材料SS含量与对照差异显著(P<0.05),其中BQ的SS含量上升最快,是对照的1.24倍,TD的SS含量上升最慢,是对照的1.04倍。5 ℃处理下,所有供试材料SS含量显著高于对照,TD上升幅度最大,为73.40%,BQ上升幅度最小,为53.5%。0 ℃处理下,除NQ的SS含量显著增加以外,其余材料SS含量较5 ℃处理有所下降,但3种供试材料SS含量均与对照差异显著(P<0.05),其中NQ的SS含量达到最高,为对照的2.14倍。
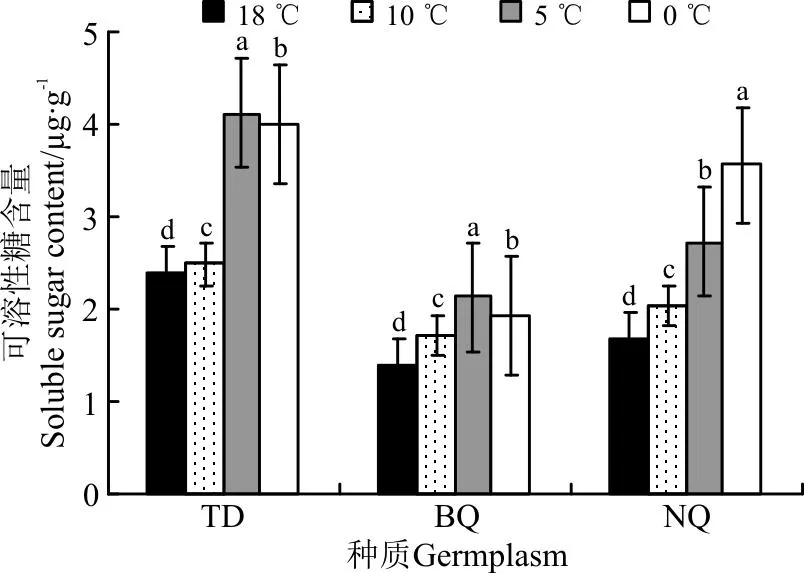
图4 3种供试牧草叶片可溶性糖(SS)含量的变化
2.5低温胁迫对披碱草叶片丙二醛(MDA)含量的影响
3个供试材料随着温度降低,MDA含量均显著高于对照(P<0.05),但变化规律不一(图5)。10 ℃处理下,所有供试材料MDA显著升高,其中TD的MDA含量上升最快,并且达到3个处理水平的最大值,是对照的2.14倍;NQ的MDA上升最慢,为对照的1.11倍。5 ℃处理下,TD的MDA含量较10 ℃时下降,而BQ与NQ的MDA含量都显著升高,并且达到3个处理水平的最高值,BQ的MDA含量为19.75 μmol·g-1,是对照的1.81倍;NQ的MDA含量为20.63 μmol·g-1,是对照的1.58倍。0 ℃处理下,TD的MDA含量较5 ℃时有所回升,达到对照的1.65倍;其余材料MDA含量反而较5 ℃时降低,尤其是BQ,其MDA含量降低幅度较大,为19.74%。

图5 3种供试牧草叶片丙二醛(MDA)含量的变化
2.6低温胁迫对披碱草叶片游离脯氨酸(Pro)含量的影响
3个材料随温度的降低,Pro含量逐渐升高(图6)。10 ℃处理下,所有供试材料Pro含量显著高于对照(P<0.05),18 ℃到10 ℃所有供试材料Pro含量增长缓慢,增幅在2.29%~23.12%,平均增幅为11.29%。当温度降至5 ℃,所有材料Pro含量显著高于对照(P<0.05),10 ℃到5 ℃之间供试材料Pro含量增长迅速,其中TD的Pro含量增幅最大,为102.77%,其余材料增幅均低于50%,平均增幅为55.36%。温度降至0 ℃,3个材料Pro含量继续上升,但增加幅度较小,其中TD的Pro含量增幅最大,为17.28%,达到对照的2.58倍;BQ的Pro含量增幅最小,为1.28%,达到对照的1.44倍。
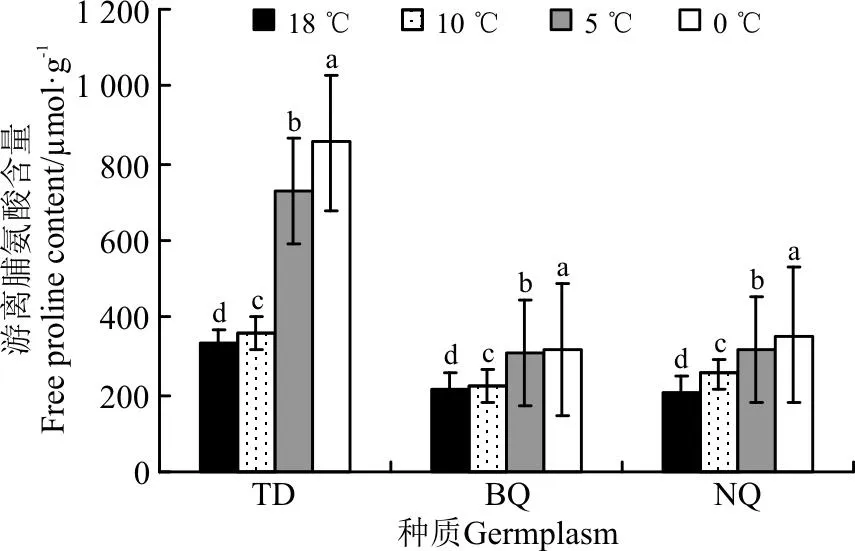
图6 3种供试牧草叶片游离脯氨酸(Pro)含量的变化
2.7低温胁迫对披碱草叶片超氧化物歧化酶(SOD)活性的影响
供试牧草叶片的超氧化物歧化酶活性随温度的降低呈现出不同的变化规律,但是整体呈上升的趋势(图7)。10 ℃时,所有供试材料SOD活性较对照显著增强(P<0.05)。温度下降到5 ℃时,TD的SOD活性相对10 ℃有所减弱,且差异不显著(P>0.05);BQ的SOD活性明显增强,并达到所有处理水平的最大值,为271.47 U·(g·min)-1;NQ的SOD活性较10 ℃显著降低(P<0.05)。温度降低到0 ℃时,除BQ的SOD活性减弱外,其余材料的SOD活性显著增强,其中NQ的SOD活性增强最快,达对照的1.65倍。

图7 3种供试牧草叶片超氧化物歧化酶(SOD)活性的变化
2.8低温胁迫对披碱草叶片过氧化物酶(POD)活性的影响
3个材料叶片的POD活性随温度的降低而逐渐增强(图8)。10 ℃处理下,各供试材料POD活性显著高于对照(P<0.05),TD的POD活性增幅最大,为97.2%。温度降至5 ℃,TD的POD活性达到对照组的2.31倍;BQ的POD活性较10 ℃差异不显著(P>0.05),增幅为1.6%;NQ的POD活性较10 ℃时显著升高,增幅为151.91%。温度降至0 ℃,BQ的POD活性较10 ℃显著增大,增幅为204.15%,是对照的3.18倍,TD与NQ的POD活性分别是对照的2.91、4.13倍。

图8 3种供试牧草叶片过氧化物酶(POD)活性的变化
2.93个披碱草属牧草苗期抗寒性综合评价
植物的抗寒性不能用单一指标来判定,本研究中5 ℃时3种供试牧草受冷害较小并且各指标变异系数较大,故采用5 ℃时3种牧草的各项指标作为其苗期抗寒能力综合评价的实测值,并通过权重值衡量比较。结果表明:BQ抗寒综合评价值最大,为0.71,抗寒性最强;TD抗寒综合评价值为0.52,抗寒性中等;NQ抗寒综合评价值为0.48,抗寒性弱(表2)。
2.103个材料各生理指标与苗期抗寒相关性分析
将5 ℃时3种供试牧草各指标的相对值与综合评价值进行相关性分析,结果显示(表3), 3种供试牧草在低温胁迫下可溶性蛋白含量的相对值与综合评价值显著负相关(P<0.05),相关系数为-1.00。相对膜透性、丙二醛、超氧化物歧化酶、可溶性糖、游离脯氨酸、过氧化物酶的相对值与抗寒综合评价值差异不显著(P>0.05)。本研究还对叶绿素含量相对值与抗寒综合评价值间进行了相关性分析,叶绿素含量相对值与抗寒综合评价值间呈负相关(P>0.05)。

表2 3个供试牧草苗期叶片抗寒性各指标隶属函数值、权重及评价值
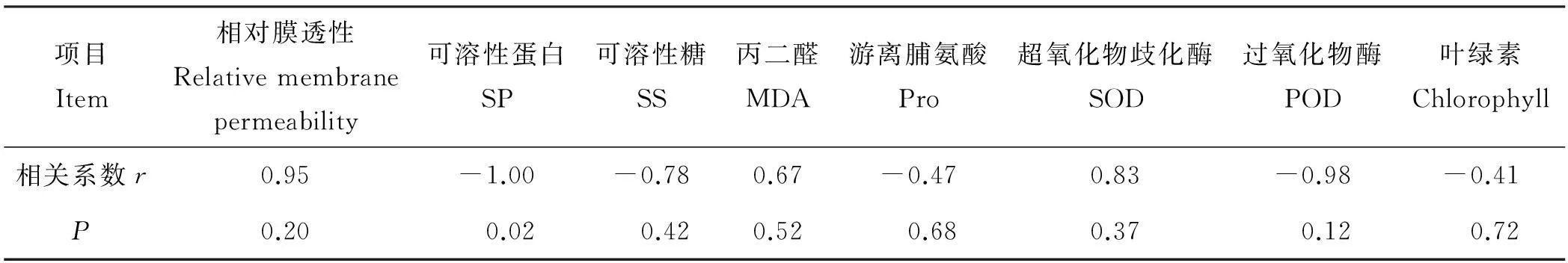
表3 3个供试披碱草各生理指标与苗期抗寒相关性分析结果
3讨论
3.1质膜透性与披碱草抗寒性的关系
植物耐寒性的强弱与质膜透性的变化息息相关。有研究表明,质膜透性与细胞受损程度呈正比,植物质膜透性越强,其细胞与组织就越容易受到损害,故而其抗逆性就越差[11]。本研究表明,3个供试牧草叶片的相对膜透性随温度的降低呈先增大后减小的趋势,其中同德老芒麦相对膜透性最大,受到的伤害也最大;巴青垂穗披碱草在5 ℃时,相对膜透性显著升高,但温度降到0 ℃时,相对膜透性显著减小,降幅远大于其余两种材料,表明巴青垂穗披碱草有很强的抗寒调节能力;那曲垂穗披碱草相对膜透性0 ℃较5 ℃时显著升高,但幅度比同德老芒麦小。仅从相对膜透性这一指标来看,巴青垂穗披碱草的抗寒性最强,西藏当地品种巴青垂穗披碱草和那曲垂穗披碱草的抗寒性要强于青海同德老芒麦。
3.2可溶性糖、可溶性蛋白、游离脯氨酸与3种牧草抗寒性的关系
细胞内重要的渗透调节物质和营养物质对其抗寒性有关键的调节作用,其中可溶性糖、可溶性蛋白、游离脯氨酸对细胞的保水能力以及保护生物膜起关键作用[12]。正常状态下,植物体内的可溶性糖、可溶性蛋白、游离脯氨酸含量较少,当遇到逆境时游离脯氨酸含量会急速增加,同时植物中的淀粉可转化为可溶性糖、可溶性蛋白也相对增多以抵御冷害,当细胞内的可溶性糖、可溶性蛋白、游离脯氨酸含量积累到一定量的时候会维持相对稳定[13-16]。本研究中,随着低温胁迫的加剧,那曲垂穗披碱草和同德老芒麦的可溶性糖与可溶性蛋白含量急剧增加,以适应寒冷胁迫;巴青垂穗披碱草可溶性糖含量与可溶性蛋白含量也有增加,但增加幅度很小,表明巴青垂穗披碱草自身耐受寒冷胁迫的能力较强。本研究中,3个材料的叶片游离脯氨酸含量随温度的降低而显著升高,这与吕优伟等[5]在低温胁迫下9个早熟禾叶片游离脯氨酸含量变化规律基本一致,其中同德老芒麦的游离脯氨酸含量随低温胁迫的加剧急剧升高,巴青垂穗披碱草与那曲垂穗披碱草游离脯氨酸含量也有升高,但升高幅度远不如同德老芒麦,表明巴青垂穗披碱草与那曲垂穗披碱草具有较强的抗寒能力,同德老芒麦抗寒能力较弱。仅从这3个指标来看,3种供试材料的可溶性糖、可溶性蛋白、游离脯氨酸含量随着寒冷胁迫的加剧而不断增加,同时说明巴青垂穗披碱草对寒冷的耐受能力很强。
3.3丙二醛与披碱草抗寒性的关系
植物体内膜脂的过氧化会导致冷害,这种冷害最明显的变化就是MDA的大量积累,所以,MDA成为鉴别植物抗逆性的重要指标之一[17-18]。本研究中,3种供试材料的MDA含量随着低温胁迫的加剧都有一个急剧积累的过程,10 ℃时,同德老芒麦MDA含量急剧增加,增加幅度远大于另外两种供试材料,说明同德老芒麦的抗寒能力相对较弱,10 ℃的寒冷胁迫足以让其MDA大量增加以抵御寒冷,同时也说明另外两种材料的抗寒能力较强。随着温度降低到5 ℃时,同德老芒麦MDA含量显著降低,表明同德老芒麦的抵御外界寒冷调节机制起了保护作用,但不能说明其抗寒性能强,因为温度降到0 ℃时MDA含量又显著升高且叶片受到的伤害程度最大。然而另外两种材料在0 ℃时MDA含量显著降低,并且巴青垂穗披碱草含量降低幅度最大,所以仅从MDA这项指标来看,寒冷胁迫会导致植物体内MDA的积累,同时也说明巴青垂穗披碱草抗寒能力最强,同德老芒麦最弱。
3.4超氧化物歧化酶、过氧化物酶与披碱草抗寒性的关系
植物受到胁迫时会产生氧自由基,而过氧化物酶和超氧化物歧化酶可以清除植物体内产生的活性氧,进而减少逆境对植物的伤害[19-20]。本研究中,3种供试材料的叶片SOD活性随低温胁迫的加剧都出现了急剧增加的过程,表明供试材料受到寒冷胁迫时产生了氧自由基以抵御寒冷。从变化规律来看,那曲垂穗披碱草活性氧产生的最多,叶片受到的伤害也越大,巴青垂穗披碱草次之,同德老芒麦最小,说明那曲垂穗披碱草抵御寒冷的能力较弱,试验给予的温度迫使其产生大量的氧自由基来消除体内活性氧,进而抵御寒冷;同时也说明巴青垂穗披碱草与同德老芒麦对试验低温耐受力强,无需产生大量的氧自由基来抵御寒冷。 3种供试材料叶片POD活性随着低温胁迫的加剧而急剧增加,处理温度在10和5 ℃时,巴青垂穗披碱草POD活性变化不大,而且受伤害程度最小;同德老芒麦与那曲垂穗披碱草POD活性显著增加,其中那曲垂穗披碱草POD活性增加幅度最大,受伤害程度也最大。当温度继续降低到0 ℃时,巴青垂穗披碱草POD活性急剧增加,为对照的3.18倍,但受伤害程度较小,而那曲垂穗披碱草POD活性达到对照的4.13倍,受伤害程度最大。仅从这两个指标来看,植物受到寒冷胁迫时过氧化物酶和超氧化物歧化酶的活性会增强,同时说明巴青垂穗披碱草具有较强的抗寒性,那曲垂穗披碱草的抗寒性在三者中最弱。
3.5叶绿素与披碱草抗寒性的关系
叶绿素是植物光合作用的主要器官,逆境胁迫下,部分光合机构失活或者遭到破坏,为了适应逆境,植物体内的叶绿素含量会发生相应的变化[21]。有研究表明,随着植物受到的寒冷胁迫加剧,叶绿素含量呈下降趋势[22-23]。本研究表明,叶绿素含量随着低温胁迫的加剧,各材料叶绿素含量呈上升趋势,这与前人研究的结果相反。本研究中,温度是逐步降低到目标温度,并且到达目标温度后,各处理仅有8 h,只是模拟了一个冷胁迫环境,而植物在逆境胁迫下叶绿素部分光合机构失活或者遭到破坏需要一个很长的过程,加之前人的试验设计中不是逐步降低温度,而是直接冷冻,这样植物失去了一个适应的过程,可能导致叶绿素不可逆的破坏,故而前人试验中叶绿素含量降低了,而本研究中叶绿素含量反而升高了。仅从本研究可以看出,随着寒冷胁迫的加剧,叶绿素含量升高了,同时结合前人的研究结果,也能说明低温胁迫的方式不同,植物体内的叶绿素含量会发生相应的变化。所以,叶绿素与抗寒性的关系还需要进一步跟踪研究。
3.6披碱草抗寒性与衡量指标的关系
植物的抗寒性与多种生理因素有关,因此不能用单一的指标来衡量[24]。本研究经过综合分析,不同程度的寒冷胁迫影响了3种披碱草属牧草的生理特性,巴青垂穗披碱草抗寒性优于同德老芒麦和那曲垂穗披碱草。巴青垂穗披碱草抗寒性指标主要在相对膜透性、可溶性糖、可溶性蛋白、游离脯氨酸、丙二醛和超氧化物歧化酶活性方面优于另外两种材料,而过氧化物酶活性衡量指标明显低于同德老芒麦与那曲垂穗披碱草,故而巴青垂穗披碱草在各个指标的相互影响、综合调控下具有较强的抗寒性。同德老芒麦过氧化物酶活性衡量指标占主要优势,可溶性蛋白含量高于其它材料,可溶性糖、超氧化物歧化酶活性衡量指标强于那曲垂穗披碱草。那曲垂穗披碱草的相对膜透性、游离脯氨酸、丙二醛衡量指标强于同德老芒麦,但弱于巴青垂穗披碱草。综合衡量,同德老芒麦的抗寒性大于那曲县垂穗披碱草,所以仅从本研究的结果可以反映出:可溶性糖、可溶性蛋白、超氧化物歧化酶活性、过氧化物酶活性是衡量植物抗寒性强弱的主要因素。因为可溶性糖、可溶性蛋白在植物细胞中起渗透调节作用,提高细胞的保水能力;超氧化物歧化酶、过氧化物酶能有效清除植物体内积累的氧自由基,从而减轻质膜的受伤程度,所以本研究认为这4个指标是衡量植物抗寒性强弱的主要因素。
4结论
低温影响了3份供试材料的生理特性,经过综合评价得出巴青县披碱草抗寒性最强,同德老芒麦次之,那曲县披碱草抗寒性最弱。
参考文献References:
[1]徐宗学,孟翠玲,巩同梁,隋彩虹.西藏自治区气温变化趋势分析.自然资源学报,2009,24(1):162-170.
Xu Z X,Meng C L,Gong T L,Sui C H.Long-term trend analysis for air temperature in the Tibetan region.Journal of Natural Resources,2009,24(1):162-170.(in Chinese)
[2]王代军,温洋.温度胁迫下几种冷季型草坪草抗性机制的研究.草业学报,1988,7(1):75-80.
Wang D J,Wen Y.Effects of temperature on growth of cold-season turfgrasses.Acta Prataculturae Sinica,1988,7(1):75-80.(in Chinese)
[3]卢宝荣.披碱草属与大麦属系统关系的研究.植物分类学报,1997,35(3):193-207.
Lu B R.A study on systematic relationships betweenElymusandHordeum(Poaceae).Acta Phytotaxonomica Sinica,1997,35(3):193-207.(in Chinese)
[4]付娟娟,刘建,孙永芳,褚希彤,苗彦军,呼天明,许岳飞.冷胁迫对2种垂穗披碱草生长和生理特性的影响.草地学报,2014,22(4):789-795.
Fu J J,Liu J,Sun Y F,Chu S T,Miao Y J,Hu T M,Xu Y F.Effects of cold stress on the growths and physiological characteristics of twoElymusnutansvarieties.Acta Agrestia Sinica,2014,22(4):789-795.(in Chinese)
[5]吕优伟,贺佳圆,白小明,董沁,雷娅伟.9个野生早熟禾对低温胁迫的生理响应及苗期抗寒性评价.草地学报,2014,22(2):326-333.
Lyu Y W,He J Y,Bai X M,Dong Q,Lei Y W.Evaluation of physiological responses and resistances of nine wildPoato low temperature.Acta Agrestia Sinica,2014,22(2):326-333.(in Chinese)
[6]孔祥生,易现峰.植物生理学实验技术.北京:中国农业出版社,2008:132-250.
Kong X S,Yi X F.Experiment Technology of Plant Physiology.Beijing:China Agriculture Press,2008:132-250.(in Chinese)
[7]中国科学院上海植物生理研究所,上海市植物生理学会.现代植物生理学实验指南.北京:科学出版社1999:305-306.
[8]张立军.植物生理学实验教程.北京:中国农业大学出版社,2012:32-104.
Zhang L J.The Tutorial of Plant Physiology Experiment.Beijing:China Agricultural University Press,2012:32-104.(in Chinese)
[9]王赞,李源,吴欣明,高洪文,孙桂芝.PEG渗透胁迫下鸭茅种子萌发特性及抗旱性鉴定.中国草地学报,2008,30(1):50-54.
Wang Z,Li Y,Wu X M,Gao H W,Sun G Z.Study on germination characteristics and drought-resistance evaluation ofDactylisglomerataL. under osmotic stress.Chinese Journal of Grassland,2008,30(1):50-54.(in Chinese)
[10]贺佳圆.9个野生早熟禾种质材料抗寒性研究.兰州:甘肃农业大学硕士学位论文,2012.
He J Y.Studies on cold-resistance of nine varieties of wildPoaL.Master Thesis.Lanzhou:Gansu Agricultural University,2012.(in Chinese)
[11]魏臻武,王槐三.2种草坪草抗寒特性及其超氧化物酶的作用.草业科学,1998,15(2):62-66.
Wei Z W,Wang H S.Cold acclimation and cold hardiness property of various types of turfgrasses.Pratuacultural Science,1998,15(2):62-66.(in Chinese)
[12]Jian L C,Sun L H,Shi G S.Acytochemical study on the glycoprotein satcellular membranes in different cold resistant winter wheat cultivars during cold acclimation.Acta Biologiae Experimentalis Sinica,1991,24(3):249-257.
[13]刘香萍,崔国文,李国良,杜广明.紫花苜蓿主根内非结构性碳水化合物累积及其与抗寒性的关系.中国草地学报,2010,32(2):113-116.
Liu X P,Cui G W,Li G L,Du G M.Relationships between non-structure carbohydrates accumulation in taproot of alfalfa and coldhardiness.Chinese Journal of Grassland,2010,32(2):113-116.(in Chinese)
[14]陶雅,孙启忠.不同紫花苜蓿品种可溶性糖、全氮、丙二醛含量动态变化及其与抗寒性关系研究.中国农业科技导报,2008,10(S1):56-60.
Tao Y,Sun Q Z.Dynamic variation of soluble sugar,total nitrogen and malondialdehydein different alfalfa varieties and their effect on alfalfa’s cold resistance.Journal of Agricultural Science and Technology,2008,10(S1):56-60.(in Chinese)
[15]张小英,卫智军,陈立波,刘磊,褚文彬.四个紫花苜蓿品种对低温条件的生理适应性.中国草地学报,2008,30(3):48-51.
Zhang X Y,Wei Z J,Chen L B,Liu L,Chu W B.The physiological adaptability of four alfalfa cultivars to lower temperature in autumn and winter.Chinese Journal of Grassland,2008,30(3):48-51.(in Chinese)
[16]李轶冰,杨顺强,任广鑫,冯永忠,张强,李鹏.低温处理不同禾本科牧草的生理变化及其抗寒性比较.生态学报,2009,29(3):1341-1347.
Li Y B,Yang S Q,Ren G X,Feng Y Z,Zhang Q,Li P.Changes analysis in physiological properties of several gramineous grass species and cold-resistance comparison on under cold stress.Acta Ecologica Sinica,2009,29(3):1341-1347.(in Chinese)
[17]Wise R R,Naylar A W.Chilling-enhanced photooxidation:Evidence for the role of singlet oxygen and superoxide in the breakdown of pigments and endogenous antioxidants.Plant Physiology,1987,83(2):278-282.
[18]Pan X Y,Cao Q D,Wang G X.Evaluation of lipid peroxidation for use in selection of cold hardiness cultivars of almond.Acta Ecologica Sinica,2002,22(11):1902-1911.
[19]张新兰.不同品种紫花苜蓿叶片离体干旱胁迫过程中抗氧化酶活性动态.草业科学,2008,25(2):77-83.
Zhang X L.Study on activity of antioxidative enzymes and drought resistance of alfalfa leaf under drought stress.Pratuacultural Science,2008,25(2):77-83.(in Chinese)
[20]周瑞莲,张承烈.水分胁迫下紫花苜蓿叶片含水量质膜透性、活性变化与抗旱性关系研究.中国草地,1991,13(2):20-24.
Zhou R L,Zhang C L.Alfalfa drought resistance:Correlated with changes of relative water content,membrane permeability and activities of superoxide dismutase and catalase in different water stress.Grassland of China,1991,13(2):20-24.(in Chinese)
[21]周蕴薇,刘艳萍,戴思兰.用叶绿素荧光分析技术鉴定植物抗寒性的剖析.植物生理学通讯,2006,42(5):945-950.
Zhou Y W,Liu Y P,Dai S L.Identification of cold resistant plants by chlorophyll fluorescence analysis technique.Plant Physiology Communications,2006,42(5):945-950.(in Chinese)
[22]刘琳.四川野生假俭草的抗寒性研究.雅安:四川农业大学硕士学位论文,2004.
Liu L.Study on cold resistance of Sichuan wild Centipedegrass.Master Thesis.Ya’an:Sichuan Agricultural University,2004.(in Chinese)
[23]萧洪东,王惠珍,喻敏.草坪抗逆增绿剂对细叶结缕草抗寒性的影响.草业科学,2013,30(9):1344-1348.
Xiao H D,Wang H Z,Yu M.Influence of turfass stress resistance and green enhancer on chilling resistance ofZoysiatenuifolia.Pratuacultural Science,2013,30(9):1344-1348.(in Chinese)
[24]南丽丽,师尚礼,陈建纲,朱新强,郭全恩,赵文翰.不同根型苜蓿根系对低温胁迫的响应及其杭寒性评价.中国生态农业学报,2011,19(3):619-625.
Nan L L,Shi S L,Chen J G,Zhu X Q,Guo Q E,Zhao W H.Field evaluation of the response and resistance to low temperature of alfalfa root with different root types during over-wintering.Chinese Journal of Eco-Agriculture,2011,19(3):619-625.(in Chinese)
(责任编辑张瑾)
Physiological responses to low temperature stress and cold tolerance evaluation in threeElymusspecies
Zhang Shang-xiong1, Nimapingcuo2, Xu Ya-mei1,Miao Yan-jun1, Baosaihenna1, Zhang Wei-hong1
(1.College of Agriculture and Animal Husbandry of Tibet University, Linzhi 860000, China;2. Tibet Academy of Agricultural and Animal Science, Lhasa 850000, China)
Abstract:In order to screencold tolerant Elymus germplasm, the 21 days aged seedlings of three Elymus species from Tibet were used to investigate the physiological, biochemical responses and resistanceunder different low temperature treatments(18, 10, 5, 0 ℃) with 18 ℃ as check.With the increase of low temperature stress Elymus sibiricus cv. Tongde had maximum relative membrane permeability, but Baqing (E. nutans) is the minimum. The contents of chlorophyll in Baqing showed a trend of rising first then falling, but the content of Chlorophyll in Tongde and Naqu (E. nutans) increased significantly (P<0.05). The soluble protein content in Baqing had the smallest increase and it only increased 2.59 times when compared with control; The soluble protein content in Tongde and Naqu increased significantly (P<0.05), and they increased 5.31 times than the control. The soluble sugar content in Baqing and Tongde showed a trend of rising first and then falling, but the soluble sugar content in Naqu increased significantly (P<0.05) and it is 2.14 times of the control. The MDA content in Tongde had the biggest increase; the MDA content in Baqing decreased significantly at 0 ℃ when compared with that at 5 ℃(P<0.05). The free proline in all materials tested increased.Tongde had the maximum Pro content, and it increased 2.58 times when compared with control; Baqing had the minimum Pro content, it only increased 1.44 times when compared with control. The superoxide dismutase activities of Baqing showed a trend of rising first then falling. But the superoxide dismutase activities of Naqu increased significantly (P<0.05) and it is 1.65 times of control. The peroxidase activities in Naqu is the maximum and it increased 4.13 times when compared with control; The peroxidase activities in Tongde is the maximum and it is 2.19 times of control. Membership function analysis showed the order of cold resistance of three Elymus species: Baqing>Tongde>Naqu.
Key words:Elymus; cold stress; physiological characteristics; cold tolerance evaluation; Tibetan
DOI:10.11829/j.issn.1001-0629.2015-0489
*收稿日期:2015-08-28接受日期:2015-10-19
基金项目:国家科技支撑计划项目(2011BAD17B05-4);西藏自治区科技厅基金项目(2013NM016);西藏自治区科技重点项目(Z2013C02N02_04);西藏大学农牧学院高原作物栽培与耕作学重点实验室
通信作者:苗彦军(1971-),男,内蒙古四子王旗人,副教授,硕士,主要从事西藏野生牧草驯化、选育及植被恢复的教学与科研工作。
中图分类号:S543+.903.4;Q945.78
文献标志码:A
文章编号:1001-0629(2016)6-1154-10*
Corresponding author:Miao Yan-junE-mail:myj666@126.com
张尚雄,尼玛平措,徐雅梅,苗彦军,包赛很那,张卫红.3个披碱草属牧草对低温胁迫的生理响应及苗期抗寒性评价.草业科学,2016,33(6):1154-1163.
Zhang S X,Nimapingcuo,Xu Y M,Miao Y J,Baosaihenna,Zhang W H.Physiological responses to low temperature stress and cold tolerance evaluation in threeElymusspecies.Pratacultural Science,2016,33(6):1154-1163.
第一作者:张尚雄(1988-),男,甘肃古浪人,在读硕士生,主要从事西藏野生牧草方面的研究。E-mail:zhangshangx@126.com
E-mail:myj666@126.com
