2种不同生活型杉叶藻的光合特性
2016-07-14高丽楠
高丽楠
(成都大学 旅游与经济管理学院,成都 610106)
2种不同生活型杉叶藻的光合特性
高丽楠
(成都大学 旅游与经济管理学院,成都610106)
利用Junior-PAM调制叶绿素荧光仪测定九寨沟箭竹海沉水和挺水杉叶藻(Hippurisvulgaris)的快速光响应曲线、最大光化学效率(Fv/Fm)、光化学淬灭(qP)和非光化学淬灭(NPQ)的日动态,探讨不同生活型杉叶藻的光合特性对九寨沟高原湖泊环境适应的特性和机制。结果表明,沉水杉叶藻的最大相对电子传递速率(rETRmax)和半饱和光强(Ek)的日变化均呈双峰型曲线,而挺水杉叶藻rETRmax和Ek日变化呈单峰曲线;沉水和挺水杉叶藻光能利用效率均呈早晚高、中午低的近“V”型曲线。挺水杉叶藻的rETRmax、Ek、qP和NPQ日平均值极显著高于沉水杉叶藻,挺水杉叶藻通过增加光耗散尽量降低光抑制程度,具有更高光合能力和对强光的耐受能力。以上结果说明:不同生活型杉叶藻的叶绿素荧光参数有一定差异,挺水杉叶藻对强光的适应能力强于沉水杉叶藻。
沉水杉叶藻;挺水杉叶藻;快速光响应曲线
九寨沟自然保护区位于青藏高原东缘,具有脆弱性和敏感性,是高寒地区独具特色的喀斯特地貌景观。近年来,九寨沟的旅游活动呈现空前繁荣,但是在带来巨额经济收入的同时,巨大的游客人数、大量的旅游活动以及修建旅游基础设施给景区的植被、水体和土壤等造成一定的负面影响。水生植物生物量急速增长,植物残体不断淤积于湖底,湖泊生态系统开始向陆生生态系统演变,沼泽化趋势日益严重。因此,湖泊沼泽化进程主要受制于水生植物初级生产力的变化。目前,对该区域湖泊水生植物的研究多集中在植被多样性调查、植物区系地理分析等基础研究[1-2],缺乏对水生植物生长状况的研究。
九寨沟箭竹海处于湖泊沼泽化过程的挺水植物阶段,湖泊沉积速度加快。如果水量继续下降,湖泊湿地萎缩,沼泽化过程将更快。九寨沟分布广泛的水生植物杉叶藻为北极—高山分布类型,是青藏高原亚寒带地区水生植被典型的挺水植物群落,是高原高寒环境的一种指示群落[3]。本试验以九寨沟优势水生植物杉叶藻(Hippurisvulgaris)为研究对象,探讨箭竹海的不同生活型植物—挺水和沉水杉叶藻的叶绿素荧光参数日变化特征,讨论不同生活型植物对湖泊水环境生态适应的差异,有助于理解植物从水生生境向陆地生境过渡的可变适应性,为九寨沟水生植物的生长过程、湖泊沼泽化治理和生态与环境保护提供科学依据。
1材料与方法
1.1试验样地
九寨沟自然保护区位于四川省阿坝藏族羌族自治州九寨沟县中南部(103°46′~104°05′E,32°55′~33°16′N),地处青藏高原东缘岷山山脉南段尕尔纳峰北麓,是长江水系嘉陵江源头的支沟,地貌为高山峡谷区,地势南高北低,海拔1 996~4 764 m。九寨沟具有冷凉干燥的季风气候特征,年平均气温6.5 ℃,极端低温为-19.1 ℃,极端高温为33.7 ℃;年平均降水量681 mm,降雨多集中在5-10月;年日照时数为1 600 h,相对湿度为60 %~70 %[4]。
1.2测定方法及数据处理
1.2.1叶绿素荧光参数野外试验选在水生植物生长旺季的7月14-24日进行。选择晴朗无云的天气,分别测定箭竹海沉水和挺水杉叶藻叶绿素荧光参数的日变化,7:00-17:00,每2 h测定1次。叶绿素荧光测定由脉冲调制叶绿素荧光仪Junior-PAM (Walz, Germany)连接到手持三星Q1U-000电脑上进行,由WinControl-3软件控制。叶绿素荧光参数测定过程中,用磁性叶夹将叶片夹住,并且调节光纤末端到样品的距离为2 mm。
用叶绿素荧光仪Junior-PAM测定快速光响应曲线(Rapid light curves, RLCs),随机选择5株植株,测定植株顶端以下第3~4片叶片。测量由电脑控制,沉水植物的光化光强度(Photosynthetic active radiation,PAR)梯度设置为66、90、125、190、285、 420、625、820 μmol·m-2·s-1,挺水植物的光化光强度梯度设置为125、190、285、420、625、820、1 150、1 500 μmol·m-2·s-1。打开测量光和光化光,适应10 s后打开饱和脉冲,之后提升光化光强度并适应10 s,再打开饱和脉冲,如此重复8 次。打开饱和脉冲前的荧光Fs,打开饱和脉冲得到最大荧光Fm′,由此可以得出PSⅡ的有效量子产量 [Yeild= (Fm′-Fs)/Fm′]。根据Yeild和PAR计算相对电子传递速率 (Relative electron transport rate,rETR=Yeild×PAR×0.5×0.84, μmol·m-2·s-1)[5],其中0.5是假设吸收光能被光系统Ⅰ(PSⅠ)和光系统Ⅱ(PSⅡ)平均分配,0.84是植物的经验性吸光系数[6]。
快速光曲线拟合采用Platt等的方程[7]:
rETR=rETRmax×(1-e-a×PAR/rETRmax) ×e-β×PAR/rETRmax,式中,rETRmax(Maximum relative electron transport rate, μmol·m-2·s-1)为无光抑制时的最大潜在相对电子传递速率,α(Photosynthetic efficiency)为快速光曲线的初始斜率,β为光抑制参数。由此可以得出半饱和光强(Saturating photon flux,Ek, μmol·m-2·s-1),Ek=rETRmax/α。
用叶绿素荧光仪Junior-PAM测定最大光化学效率(Maximal quantum yield,Fv/Fm)和荧光淬灭参数,随机选择生长一致且叶片受光方向相同的 5 株植株,测定植株顶端以下第3~4片叶片。叶片暗适应 20 min后,首先开启检测光,得到暗适应条件下PSⅡ最小荧光(Fo),再由饱和脉冲光激发测得暗适应的最大荧光(Fm)。随后打开内源光化光,10 min后获得光下的稳态荧光(Fs),并再次照射饱和脉冲光以获得光下最大荧光(Fm′);最后关闭光化光,打开远红光的同时测定光下的最小荧光(Fo′)。根据这些参数可以计算出:PSⅡ最大光化学效率(Fv/Fm) = (Fm-Fo) /Fm[5];光化学淬灭Photochemical quenching (qP) = (Fm′-Fs) / (Fm′-Fo′);非光化学淬灭Non-photochemical quenching (NPQ) = (Fm-Fm′) /Fm′[8]。
1.2.2光合有效辐射强度和水温 与叶绿素荧光参数同步测定,采用AccuPAR Lp-80 Ceptometer测定沉水和挺水杉叶藻顶端的光合有效辐射,利用温度计测定水体温度(Water temperature,Tw,℃)。
1.2.3叶绿素质量分数将沉水和挺水杉叶藻放在装有少量湖水的低温箱,并在最短时间内运至九寨沟实验室。称取新鲜植株顶端叶片约1.6 g,擦净组织表面,剪碎,以φ=80%的丙酮进行研磨。用紫外分光光度计(UV-1240-PC, Shimadzu)测定663 nm和645 nm处的吸光度,重复测量6次,计算叶绿素a(Chla, mg·g-1)、叶绿素b(Chlb, mg·g-1)和叶绿素(a+b)[Chl(a+b), mg·g-1]质量分数[9]。
1.2.4数据分析数据以“平均数±标准差”表示,采用Spss 17.0和Origin 7.5统计软件进行数据处理及统计分析。用One-way ANOVA进行显著性分析。
2结果与分析
2.1箭竹海环境因子日变化特征
PAR的日变化都呈明显的单峰型曲线,7:00最低,随后迅速升高,在13:00达到峰值。水上和水下PAR在7:00都小于70 μmol·m-2·s-1,13:00水上PAR大于2 100 μmol·m-2·s-1,水上与水下PAR日平均值存在显著性差异,水上PAR日平均值显著大于水下PAR(P<0.01)。湖泊Tw日变化也呈明显的单峰型曲线,7:00左右为最低值,随着PAR增强而逐渐升高,在13:00左右达到最高值,为11.16 ℃,之后随着PAR减小而缓慢下降(图1)。
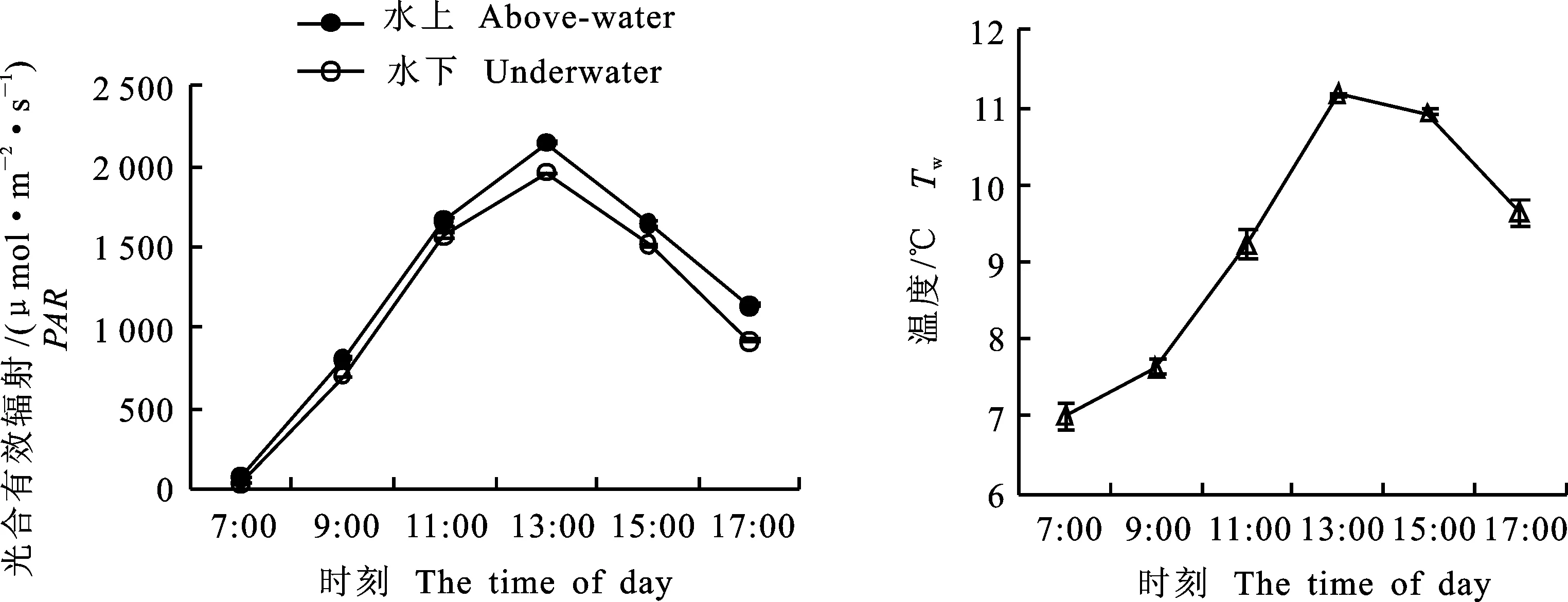
图1 水上和水下光合有效辐射和水体温度
2.2箭竹海挺水和沉水杉叶藻叶绿素荧光参数日变化特征
2.2.1快速光响应曲线从图2 可以看出,沉水杉叶藻rETRmax日变化呈典型的双峰型曲线,分别在9:00和13:00出现高峰,峰值分别为47.98和53.13 μmol·m-2·s-1,11:00出现低谷,仅为24.45 μmol·m-2·s-1。挺水杉叶藻rETRmax日变化呈单峰型曲线,7:00-13:00,随着环境因子的变化而不断上升,在13:00左右达到最大值,为156.63 μmol·m-2·s-1,之后不断下降。沉水和挺水杉叶藻Ek日变化趋势与其rETRmax基本一致(图2),挺水杉叶藻Ek(404.29 μmol·m-2·s-1)、rETRmax(113.19 μmol·m-2·s-1)日均值都极显著高于沉水杉叶藻(P<0.01)。初始斜率(α)可以反映植物叶片对光能的利用效率,表现植物叶片捕光能力的大小。沉水杉叶藻α呈早晚高、中午低的近“V”型变化趋势,11:00时α最低。挺水杉叶藻α日变化表现为:清晨α随PAR升高而迅速上升,在9:00达到峰值;此后α随PAR升高而下降,并在13:00到达谷底;之后随PAR下降α逐渐升高。挺水杉叶藻α日均值显著高于沉水杉叶藻(P<0.01)。

图2 沉水和挺水杉叶藻的最大相对电子传递速率、初始斜率和半饱和光强
2.2.2最大光化学效率 由图3看出,沉水和挺水杉叶藻Fv/Fm日变化趋势呈现先降低后升高的趋势,7:00最高,分别为0.73和0.78,之后随着PAR增强而逐渐降低,沉水和挺水杉叶藻Fv/Fm均在13:00达到最低值,表明两者均发生光抑制现象。之后随着PAR的降低,Fv/Fm开始恢复,17:00沉水杉叶藻Fv/Fm相对于7:00恢复95.71 %,可见傍晚未恢复到清晨水平,而挺水杉叶藻Fv/Fm基本恢复到清晨时的水平。挺水杉叶藻Fv/Fm日均值显著高于沉水杉叶藻(P<0.01)。
2.2.3光化学淬灭和非光化学淬灭由图4可以看出,沉水杉叶藻qP的日变化中,7:00-11:00迅速上升,之后随着PAR的增强qP降低,在13:00左右出现低谷,之后呈上升趋势。7:00-9:00,挺水杉叶藻qP迅速升高,最高峰出现在9:00,峰值为0.95,之后迅速下降,在13:00降至最低点,随后又回升。
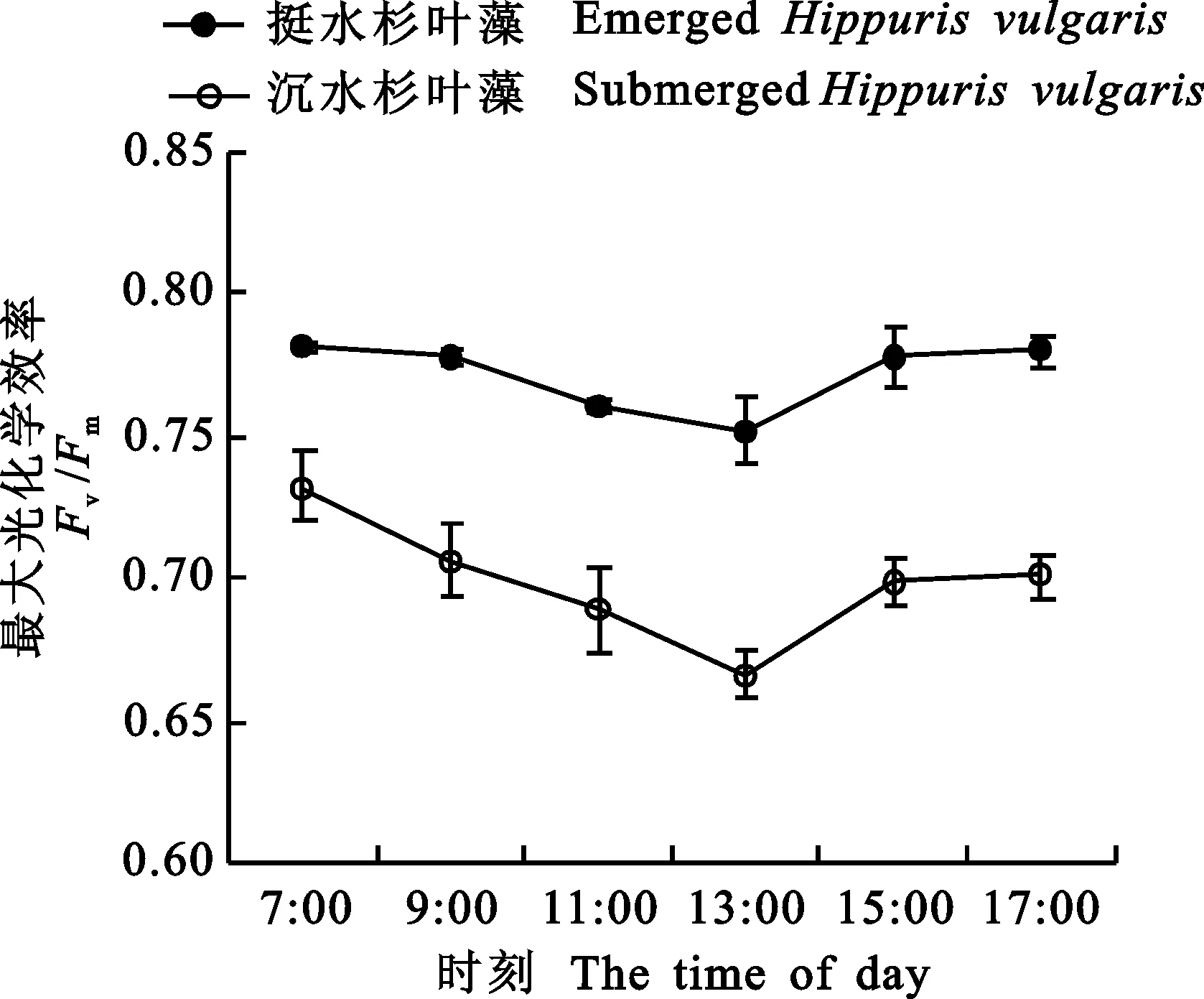
图3 沉水和挺水杉叶藻的Fv/Fm
沉水杉叶藻NPQ日变化,7: 00-11: 00逐渐降低,之后呈上升趋势,在13:00达到峰值,为0.45,之后逐渐降低(图4)。挺水杉叶藻NPQ的日变化呈单峰型曲线,随着PAR的增强,NPQ逐渐增大,在13:00达到峰值(0.52),之后随着光强的减弱,NPQ逐渐下降。挺水杉叶藻qP和NPQ日均值均极显著高于沉水杉叶藻(P<0.01)。
2.3沉水和挺水杉叶藻叶绿素质量分数的比较
叶绿素是绿色植物叶绿体内参与光合作用的重要色素,叶片中的光合色素是植物光合作用的基础,植株功能叶的叶绿素质量分数的高低在很大程度上反映植株的光合能力和生长状况。试验结果显示,Chla、Chlb、Chl(a+b)和Chla/b在沉水和挺水杉叶藻之间均有显著性差异(P<0.05),挺水杉叶藻均显著高于沉水杉叶藻(表1)。

图4 沉水和挺水杉叶藻的qP和NPQ

叶绿a/(mg·g-1)Leafchlorophylla叶绿素b/(mg·g-1)Leafchlorophyllb叶绿素(a+b)/(mg·g-1)Leafchlorophyll(a+b)叶绿素a/bLeafchlorophylla/b沉水杉叶藻 SubmergedHippurisvulgaris057±0.03a0.26±0.03a0.83±0.06a2.26±0.13a挺水杉叶藻 EmergedHippurisvulgaris1.03±0.05b0.34±0.02b1.36±0.09b3.16±0.21b
注: 同列不同小写字母表示差异显著(P<0.05)。
Note:Different lowercase letters in same column indicate significantly different according atP<0.05.
3讨 论
箭竹海挺水杉叶藻rETRmax日变化呈单峰型曲线,在13:00达到峰值,与沉水杉叶藻的rETRmax最高峰出现时间相同,水生植物都有适合其正常生命活动的温度范围,当外界环境温度在此范围内时,酶促反应会随温度升高而加快,在一定程度上能促进光合作用。而日变化趋势却不相同,沉水杉叶藻rETRmax日变化呈典型的双峰曲线,11:00出现低谷,因箭竹海湖泊水体的pH(8.4)较高[10]。有研究表明,湖泊水具有较高pH,作为溶解无机碳源的可利用CO2浓度会降低[11],从而影响沉水植物的光合作用。挺水杉叶藻日变化呈单峰型曲线,因其露出水面生长,可获得光合作用所需稳定而充足的CO2,接受的PAR没有通过水面散射进行衰减[12],所以挺水杉叶藻rETRmax显著大于沉水杉叶藻。沉水和挺水杉叶藻光能利用效率α日变化趋势表现为清晨和傍晚较高、正午较低,表明其具有调控光合作用和高效利用PAR的能力。挺水杉叶藻rETRmax、Ek和α均显著高于沉水杉叶藻,表明挺水杉叶藻具有高光合能力和对强光的耐受能力,光保护能力更强。
本研究中,7:00 沉水和挺水杉叶藻Fv/Fm分别为0.73和0.78,有研究[13]表明,Fv/Fm在非胁迫条件下比较恒定,一般为0.80~0.85,表明沉水和挺水杉叶藻可能受到胁迫,且外界胁迫对沉水植物PSⅡ反应中心造成一定损伤。目前,已有关于UV-B辐射、多环芳烃污染以及网状藻的附着对水生植物Fv/Fm影响的报道[14-15],其他因子胁迫对Fv/Fm影响还未见相关报道。结合采样区域的环境状况,可能引起采样区域沉水和挺水植物Fv/Fm降低的原因为高原较强UV-B辐射和植物体上附着的藻类[16],至于植物本身Fv/Fm对胁迫因子的耐受性如何,还有待进一步研究。其次,7:00-17:00,挺水杉叶藻的Fv/Fm大于沉水杉叶藻,说明PSⅡ的实际光能捕获效率较高,能够把所捕获的光能更多地用于光化学反应[17],因此挺水杉叶藻的光合能力(rETRmax)较高。
挺水杉叶藻qP日均值最大,即PSⅡ的电子传递活性最大,这与挺水杉叶藻的rETRmax相一致。本研究中,九寨沟夜间水温低使沉水和挺水杉叶藻qP在7:00均较低,已有报道[18]证实夜间低温抑制PSⅡ光合电子传递活性,从而造成PSⅡ反应中心开放数目下降。此后,PAR增强,随着温度的上升可能又进一步解除低温对PSⅡ光化学效率和电子传递活性的抑制作用[18],使得qP大幅度上升。另外,7:00时挺水杉叶藻qP大于沉水杉叶藻,可能是完全浸入湖泊中的沉水杉叶藻遭受夜间低温抑制作用较强。沉水杉叶藻11:00 之后qP逐渐下降,但挺水杉叶藻在9:00之后开始下降,由于PAR过强,即PSⅡ天线色素吸收的光能用于光化学反应的份额均降低,同时PSⅡ反应中心的开放程度减少,增加过剩激发能的耗散,以保护光合机构免受光抑制的破坏[19]。13:00 之后随着PAR逐渐降低,之前反应中心由于强光而处于半关闭状态向打开状态转化,qP逐渐上升,反应中心恢复。有研究[20]表明,南京固城湖苦草qP的日变化从7:00 开始,PAR逐渐增高,qP迅速降低;在13:00达到最低值,之后升高。此研究与九寨沟湖泊沉水和挺水杉叶藻qP日变化恰好相反,沉水和挺水杉叶藻qP日变化从7:00开始,随着PAR逐渐增高qP升高,之后下降。这主要是由于南京固城湖水体温度日变幅为22~25 ℃,而九寨沟高原湖泊水体温度日变幅为7~12 ℃,可能是低温抑制PSⅡ光合电子传递活性的原故。
沉水杉叶藻遭受夜间的低温Tw后,在7:00刚接受微弱的PAR,所以低温抑制吸收的光能不能用于光合电子传递而以热的形式耗散掉,所以弱光低温导致清晨时较高的NPQ和较低的qP[21]。7:00-11:00随着PAR的增加,温度上升,低温的抑制作用得到恢复,NPQ下降而qP上升;11:00-13:00 随着PAR越来越大,植物叶片发生较强的光抑制作用,所以植物通过NPQ等非光化学途径将过多的激发能耗散掉,NPQ上升qP下降,保护光合机构免受光抑制的破坏;13:00之后光照减弱,植物的光抑制作用降低,NPQ逐渐降低。与此相反,挺水杉叶藻的NPQ日变化曲线基本上为单峰型(图4),7:00时NPQ最低,上午随PAR升高而上升,表明热耗散增多,这是植物叶片防止光破坏的重要机制之一;下午随PAR减弱逐渐降低。挺水杉叶藻NPQ日均值显著高于沉水杉叶藻,因挺水杉叶藻叶片照射的PAR较强,其叶温较高使NPQ增大,植株发生光抑制的程度降低,光保护能力更强[22]。
本研究结果表明,挺水杉叶藻叶片露出水面生长,使其有充足的CO2和较高叶温,在一定程度上可以促进叶绿素a的合成[23]。挺水杉叶藻的叶绿素a质量分数较高,而叶绿素a的功能主要是将汇聚的光能转变为化学能进行光化学反应,保持体内有相对较高的叶绿素a质量分数,可以保证植物体对光能的充分利用,提高转化率[24],由此可见,挺水杉叶藻光能利用效率α较高。叶绿素a/b可以反映捕光色素复合体Ⅱ(LHCⅡ)在叶绿素结构中的比重,挺水杉叶藻叶绿素a/b高于沉水杉叶藻, 表明LHCⅡ降低[25],说明挺水杉叶藻对较强PAR产生胁迫效应导致这种变化,减少叶片对光能的捕获,降低光合机构遭受破坏的风险,这也是挺水植物适应较强PAR生境的一种自身调节机制。
综上所述,生活型不同可能是沉水和挺水杉叶藻叶绿素荧光特性显著差异的重要原因。研究结果显示,挺水杉叶藻rETRmax日均值都极显著高于沉水杉叶藻,因挺水杉叶藻露出水面生长,可以获得光合作用所需稳定而充足的CO2,接受的光强没有通过水面散射进行衰减。挺水杉叶藻的rETRmax、Ek、qP和NPQ日均值都极显著高于沉水杉叶藻,所以其通过增加光耗散尽量降低光抑制程度,具有更高光合能力和对强光的耐受能力。
Reference:
[1]齐代华,王力,钟章成.九寨沟水生植物群落β多样性特征研究[J].水生生物学报,2006,30(4):446-452.
QI D H,WANG L,ZHONG ZH CH.Studies on the β diversity analyses of aquatic plant community in Jiuzhaigou[J].ActaHydrobiologicaSinica,2006,30(4):446-452(in Chinese with English abstract).
[2]邓洪平,陈亚飞,谢大军,等.九寨沟自然保护区种子植物区系特征研究[J].西南师范大学学报(自然科学版),2005,30(3):543-547.
DENG H P,CHEN Y F,XIE D J,etal.A floristic study of seed plants on Jiuzhaigou Valley Natural Reserve[J].JournalofSouthwestChinaNormalUniversity(NaturalScienceEdition),2005,30(3):543-547(in Chinese with English abstract).
[3]王东.青藏高原水生植物地理研究[D].武汉:武汉大学, 2003.
WANG D.The geography of aquatic vascular plants of Qinghai-Xizang (Tibet) plateau[D].Wuhan:Wuhan University,2003(in Chinese with English abstract).
[4]邓贵平,颜磊,章小平.九寨沟自然保护区景观变化与保护[J].山地学报,2011,29(2):173-182.
DENG G P,YAN L,ZHANG X P.The changes of landscape and the adaptive management strategies at Jiuzhaigou Nature Reserve in Sichuan, China[J].JournalofMountainScience,2011,29(2):173-182(in Chinese with English abstract).
[5]SCHREIBER U,GADEMANN R,RALPH P,etal.Assessment of photosynthetic performance ofProchloroninLissoclinumpatellain hospite by chlorophyll fluorescence measurements[J].PlantandCellPhysiology,1997,38(8):945-951.
[6]COHEN I,NEORI A.Ulvalactucabiofilters for marine fishpond effluents.I.Ammonia uptake kinetics and nitrogen content[J].BotanicaMarina,1991,34(6):475-482.
[7]PLATT T,GALLEGOS C L,HARRISON W G.Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton[J].JournalofMarineResearch,1980,38(4):687-701.
[8]ROHACEK K.Chlorophyll fluorescence parameters.The definitions,photosynthetic meaning and mutual relationships[J].Photosynthetica,2002,40(1):13-29.
[9]ARNON D I.Copper enzymes in isolated chloroplasts.Polyphenoloxidase inBetavulgaris[J].PlantPhysiol,1949,24(1):3-15.
[10]苏君博.九寨沟水文地球化学特征及对景观演化影响研究[D].成都:成都理工大学,2005.
SU J B.Hydrogeochemistry and its influence on evolution at Jiuzhai Valley[D].Chengdu:Chengdu University of Technology,2005(in Chinese with English abstract).
[11]INVERS O,ROMERO J,PRES M.Effects of pH on seagrass photosynthesis:a laboratory and field assessment[J].AquaticBotany,1997,59(314):185-194.
[12]SAND-JENSEN K,FROST-CHRISTENSEN H.Plant growth and photosynthesis in the transition zone between land and stream[J].AquaticBotany,1999,63(63):23-35.
[13]DAWSON S P,DENNISON W C.Effects of ultraviolet and photosynthetically active radiation on five seagrass species[J].MarineBiology,1996,125(4):629-638.
[14]MARWOOD C A,SOLOMON K R,GREENBERG B M.Chlorophyll fluorescence as a bioindicator of effects on growth in aquatic macrophytes from mixtures of polycyclic aromatic hydrocarbons[J].EnvironmentalToxicologyandChemistry,2001,20(4):890-898.
[15]李强,王国祥,马婷,等.水网藻附着对亚洲苦草光合特性的影响[J].湖泊科学,2007,19(3):315-320.
LI Q,WANG G X,MA T,etal.Changes of photosynthetic characters ofVallisneriaasiaticaadhered byHydrodictyonreticulatum[J].JournalofLakeSciences,2007,19(3):315-320(in Chinese with English abstract).
[16]宋玉芝,蔡炜,秦伯强.太湖常见浮叶植物和沉水植物的光合荧光特性比较[J].应用生态学报,2009,20(3):569-573.
SONG Y ZH,CAI W,QIN B Q.Photosynthetic fluorescence characteristics of floating-leaved and submersed macrophytes commonly found in Taihu Lake[J].ChineseJournalofAppliedEcology,2009,20(3):569-573(in Chinese with English abstract).
[17]张杰,杨传平,邹学忠,等.蒙古栎硝酸还原酶活性、叶绿素及可溶性蛋白含量与生长性状的关系[J].东北林业大学学报,2005,33(3):20-21.
ZHANG J,YANG CH P,ZOU X ZH,etal.Relationships between nitrate reductase activity,chlorophyll,soluble protein contents,and growth characters ofQuercusmongolica[J].JournalofNortheastForestryUniversity,2005,33(3):20-21(in Chinese with English abstract).
[18]胡文海,黄黎锋,肖宜安,等.夜间低温对2种光强下榕树叶绿素荧光的影响[J].浙江林学院学报,2005,22(1):20-23.
HU W H,HUANG L F,XIAO Y A,etal.Effects of low temperature stress at night on chlorophyll fluorescence characteristics in leaves ofFicusmicrocarpagrowing under two levels of irradiance[J].JournalofZhejiangForestryCollege,2005,22(1):20-23(in Chinese with English abstract).
[19]宗梅,谈凯,吴甘霖.两种石楠叶绿素荧光参数日变化的比较研究[J].生物学杂志,2010,27(1):27-30.
ZONG M,TAN K,WU G L.Comparative study on diurnal changes of chlorophyll fluorescence parameters of two types ofPhotiniaserrulataLind[J].JournalofBiology,2010,27(1):27-30(in Chinese with English abstract).
[20]王立志,王国祥,俞振飞,等.苦草光合作用日变化对水体环境因子及磷质量浓度的影响[J].生态与环境学报,2010,19(11):2669-2674.
WANG L ZH,WANG G X,YU ZH F,etal.Effects of diurnal change ofV.natansphotosynthesis on environmental factors and phosphorus concentration in water[J].EcologyandEnvironmentalSciences,2010,19(11):2669-2674(in Chinese with English abstract).
[21]周艳虹,黄黎锋,喻景权.持续低温弱光对黄瓜叶片气体交换、叶绿素荧光淬灭和吸收光能分配的影响[J].植物生理与分子生物学报,2004,30(2):153-160.
ZHOU Y H,HUANG L F,YU J Q.Effects of sustained chilling and low light on gas exchange,chlorophyll fluorescence quenching and absorbed light allocation in cucumber leaves[J].JournalofPlantPhysiologyandMolecularBiology,2004,30(2):153-160(in Chinese with English abstract).
[22]李强.环境因子对沉水植物生长发育的影响机制[D].南京:南京师范大学,2007.
LI Q.Influence mechanism of environment factors on the growth and development of submerged macrophytes[D].Nanjing: Nanjing Normal University,2007(in Chinese with English abstract).
[23]蒋利鑫,于苏俊,魏代波,等.湖泊富营养化评价中的灰色局势决策法[J].环境科学与管理,2006,31(2):10-12.
JIANG L X,YU S J,WEI D B,etal.Grey situation decision method aplying to eutrophication evaluation for lakes[J].EnvironmentalScienceandManagement,2006,31(2):10-12(in Chinese with English abstract).
[24]衣艳君,李芳柏,刘家尧.尖叶走灯藓(Plagiomniumcuspidatum) 叶绿素荧光对复合重金属胁迫的响应[J].生态学报,2008,28(11):5437-5444.
YI Y J,LI F B,LIU J Y.Physiological response of chlorophyll fluorescence in mossPlagiomniumcuspidatumto mixture heavy metal solution[J].ActaEcologicaSinica,2008,28(11):5437-5444(in Chinese with English abstract).
[25]ANDERSON J M,ARO E M.Grana stacking and protection of photosystem II in thylakoid membranes of higher plant leaves under sustained high irradiance: an hypothesis[J].PhotosynthesisResearch,1994,41(2):315-326.
The authorGAO Linan, female, lecturer. Research area: plant physiological ecology. E-mail:gaolinan20081212@163.com
(责任编辑:顾玉兰Responsible editor:GU Yulan)
Photosynthetic Characteristics in Two Different Life Forms ofHippurisvulgaris
GAO Linan
(College of Tourism and Economic Management, Chengdu University,Chengdu610106, China)
Submerged and emergedHippurisvulgariswere selected in Jiuzhaigou to examine diurnal variations of rapid light curves (RLCs), maximal quantum yield (Fv/Fm), photochemical quenching (qP) and non-photochemical quenching (NPQ) using pulse-amplitude modulated fluorometer (Junior-PAM) in Jianzhu Lake.The purpose is to investigate the photosynthetic characteristics of different life forms ofH.vulgarisand adaptive strategies against environmental in alpine lakes. The results showed that the diurnal variations of submergedH.vulgarisof maximum relative electron transport rate (rETRmax) and saturating photon flux (Ek) presented a typical bimodal curve, while those of emergedH.vulgarispresented unimodal curve. The photosynthetic efficiency (α) of submerged and emergedH.vulgariswas V pattern diurnal cycle, α was the highest at sunrise and sunset, α was the lowest at noon. The daily averages ofrETRmax,Ek,qPandNPQof emergedH.vulgariswere significantly greater than that of submergedH.vulgaris.Therefore it dissipated excess light energy depending on the increase ofNPQ, emergedH.vulgariswas a plant with a strong photosynthetic capacity for acclimation to high light intensity. The results indicated that significant differences in chlorophyll fluorescence parameters were found between submerged and emergedH.vulgaris, emergedH.vulgariswas more tolerant to strong light intensity than submergedH.vulgaris.
SubmergedHippurisvulgaris; EmergedHippurisvulgaris; Rapid light curves
2016-04-01
2016-05-11
Project of Introducing Talents of Discipline to Universities (No.B08037).
1004-1389(2016)07-1067-07
2016-04-01修回日期:2016-05-11
高等学校学科创新引智计划 (B08037)。
高丽楠,女,讲师,主要从事植物生理生态学研究。E-mail:gaolinan20081212@163.com
Q945.79
A
网络出版日期:2016-06-30
网络出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20160630.1634.034.html
