甘蓝型油菜产量性状的遗传分析
2016-07-07关周博董育红李少钦张耀文田建华李殿荣陕西省杂交油菜研究中心国家油料作物改良中心陕西油菜分中心陕西杨凌712100
关周博,董育红,李少钦,张耀文,田建华,李殿荣(陕西省杂交油菜研究中心/国家油料作物改良中心陕西油菜分中心,陕西杨凌712100)
甘蓝型油菜产量性状的遗传分析
关周博,董育红,李少钦,张耀文,田建华,李殿荣
(陕西省杂交油菜研究中心/国家油料作物改良中心陕西油菜分中心,陕西杨凌712100)
摘 要:探索甘蓝型油菜亲本及杂种在不同生态区的遗传效应,为品种选育提供理论基础。本研究利用加性—显性及其与环境互作的遗传模型,分析在不同生态区域亲本和杂交组合的产量性状遗传规律,估算亲本产量性状的加性遗传效应值、亲本在杂交组合中的显性遗传效应方差值、各项方差分量的比率及对F1和F2基因型与环境效应值的预测。结果表明,产量性状受到加性和显性遗传效应的影响;在遗传效应与环境互作中,单株产量、角果数和千粒重的各遗传效应都与环境互作达到了显著性,而角粒数的遗传稳定与环境互作没有显著性。不同环境下甘蓝型油菜产量性状在F1和F2世代差异较大,总体表现为大荔试验点基因型与环境的效应值高于张掖试验点基因型与环境的效应值,但千粒重却与之相反,环境条件对千粒重的后代表型影响较大。
关键词:甘蓝型油菜;产量性状;遗传分析
甘蓝型油菜在我国种植的历史较短,但却是目前我国主栽油菜类型。自李殿荣成功选育秦优2号并成为全世界第一个大面积推广的杂交油菜品种,油菜杂种优势的突破推进了我国油菜品种从常规种向杂交种的转变,实现了油菜产量的跨越式发展,油菜生产面积和产量均占到世界油菜生产面积的三分之一[1~3]。
目前,高产杂交油菜品种的选育途径主要是利用亲本之间的配合力,选择配合力较好的杂交组合以发挥其种间杂种优势,从而实现高产的目标。研究亲本性状的遗传、强化亲本性状的改良以及杂种后代性状表现的筛选,特别是与产量相关性状遗传规律的研究,是配制高产组合成功与否的关键。邓武明等[4]发现,油菜千粒重性状的遗传符合加性—显性遗传模型,单株有效角果数、角果粒数以及单株千粒重的遗传行为符合加性—显性—上位性模型。李少钦等[5]认为,产量相关性状间存在着显著的加性遗传效应和显性遗传效应,且都存在着与环境的互作,其中千粒重的广义遗传率和狭义遗传率都较高。王瑞等[6]认为,母体效应对产量的影响较小,产量性状的遗传主要受到加性和显性效应共同控制。还有大量研究结果表明,F1代在产量与相关性状上表现出较强的杂交优势[7~9]。但由于年份不同或研究试验的区域不同,油菜产量性状的遗传可能也会随着环境的变化而表现不同。在大多数的研究中较少涉及到F2基因型与环境的互作,而且在产量性状遗传分析中,多数采用双列杂交的配合力分析方法,利用亲本和F1数据分析F2产量性状的遗传报道尚少。本文拟采用朱军[10]的加性(A)—显性(D)及其与环境(E)互作的遗传模型来分析不同环境下油菜产量性状的遗传效应,估算遗传方差分量及F1和F2群体产量性状的遗传表现,为选育高产、稳产和优质的杂交油菜品种提供理论依据。
1 材料与方法
1.1试验材料
供试的6个甘蓝型油菜亲本为性状优良的纯合自交系(品种),均来源于陕西省杂交油菜研究中心种质资源库(表1)。

表1 供试亲本(系)系谱Table 1 Pedigree of parent varieties(lines)
1.2试验方法
2012年1月于加代温室内对6个亲本(系)进行杂交,共获得15个F1杂交组合种子,同年4月到9月将所得到的杂交F1及亲本种植于张掖试验点(春播);2012年9月到2013年6月将所得到的杂交F1及亲本种植于大荔试验点(秋播)。试验采取随机区组设计,每个组合种植5行,3次重复,行长2.5 m,行距0.4 m,株距0.14 m。成熟期调查每小区单株角果数、角粒数、单株产量和千粒重。
1.3数据分析
数据处理采用朱军经典数量遗传学分析软件[10~15]。
2 结果与分析
2.1不同地区甘蓝型油菜亲本与F1产量性状的表现
6个亲本和15个杂交组合的产量性状在两个生态区的平均表现见表2。由表2可知,4个产量性状杂种F1的表型值均值均高于其亲本的表现型值,说明杂种F1的产量性状表现出了一定的正向杂种优势。在4个产量性状中,2个生态区亲本和F1的平均表现型值不同,单株产量、角果数、角粒数大荔试验点高于张掖试验点,千粒重性状表型值张掖试验点高于大荔试验点,说明存在着基因型与环境的互作现象。

表2 不同地区油菜亲本与F1产量性状的平均表现Table 2 Average phenotype values for yield traits of rapeseed parents and their F1in different areas
2.2产量性状的加性遗传效应值(A)和显性遗传效应值(D)
组合测配或品种改良中,既要注重一般配合力大小又要考虑其特殊配合力大小,亲本材料的加性遗传效应与测配组合的显性遗传效应应作为选择的依据。在表3中,1、4和6号亲本的4个产量性状在加性遗传中表现为显著或极显著;单株产量和角果数以5号亲本的加性遗传值最高,角粒数和千粒重以4号亲本的加性遗传效应值最高。遗传效应值越高说明在后代早期遗传表现越突出,4号和5号亲本可作为后代改良的理想亲本,在早期世代选择更有效。
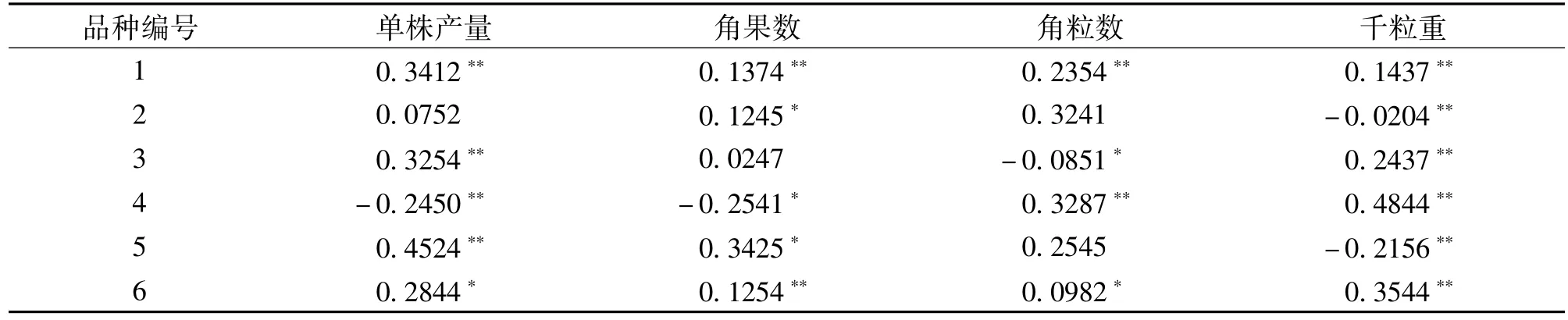
表3 6个甘蓝型油菜亲本产量性状的加性遗传效应值(A)Table 3 Additive genetic effect for yield traits of rapeseed parents
基因的加性效应可以稳定遗传,显性效应是要在具体组合中才能体现出来的,但在杂种优势利用中有着更为重要的意义。由表4信息可知,组合1 ×4、1×6、3×4、3×5、3×6在4个产量性状中显性遗传效应值都达到了显著或极显著,与加性遗传效应相对比,4号和6号亲本的加性遗传效应值较高,在组合测配中与其它亲本所配的杂种后代也出现了较高的显性效应值,但并不是亲本的加性遗传效应值都高且具有显著性其所配组合的显性遗传效应值也高,如组合4×6,两个亲本都具有较高的加性遗传效应值,但组合却不是所有性状都表现出较高的显性遗传效应值。所以,组合测配不仅要考虑到亲本的加性遗传效应值,而且还要结合组合的实际表现。
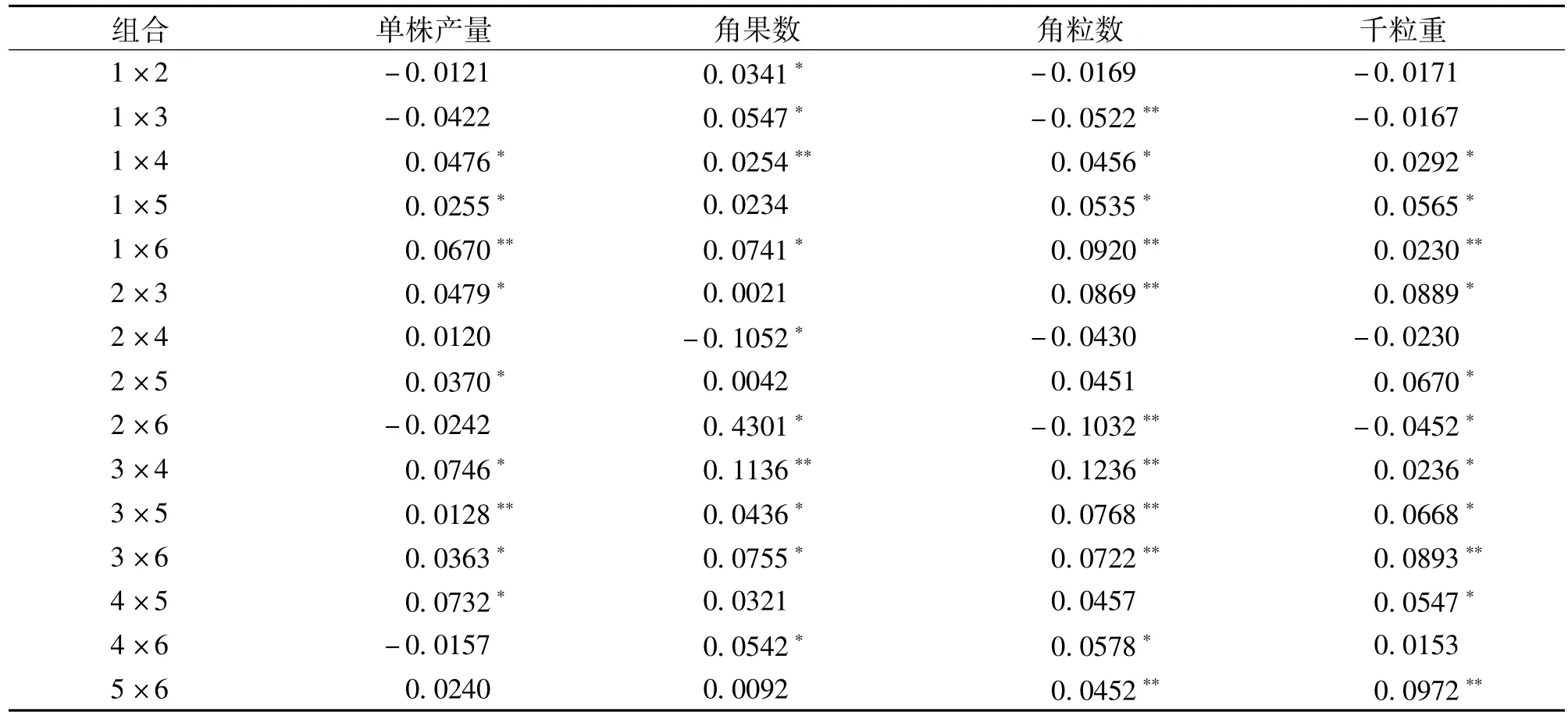
表4 杂交组合的显性遗传效应值(D)Table 4 Dominant genetic effect for yield traits of rapeseed parents
2.3产量性状的遗传方差分量分析
供试材料产量性状的各项遗传方差分量占表型方差的比值和机误方差占表型方差的比值见表5。从表5中可知,4个产量性状的加性方差比率和显性方差比率均达到了极显著水平;在与环境互作中,单株产量、角果数和千粒重的各遗传效应都与环境互作达到了显著性,说明这3个性状不仅受到遗传因素的影响,还与环境因素有很大的关系,而角粒数的遗传稳定与环境互作没有显著性。加性方差分量的比率反映了各性状的狭义遗传率,因此从表3中也可得知,角粒数的狭义遗传率最大,遗传改良过程中在早期世代进行选择最为有效。

表5 甘蓝型油菜产量性状的遗传方差分量比值估算Table 5 Estimated proportions of genetic variance components for yield traits of rapeseed
2.4在不同环境下F1和F2产量性状的基因型值预测
不同环境下产量性状在不同世代与环境互作效应差异较大。本试验中单株产量、角果数和角粒数的F1和F2世代,大荔试验点的基因型与环境的表现值高于张掖试验点基因型与环境的表现值,但千粒重性状却与之相反,表现为张掖试验点基因与环境的表现值高于大荔试验点(表6)。在单株产量、角果数和角粒数性状中,F1与F2相比较F2表型值低于F1,这也表明F2分离群体性状差异大,在相同环境中基因的变化对后代性状影响较大;但千粒重在F1和F2代中表现出较稳定的表型值,呈现出张掖试验点高于大荔试验点。

表6 不同环境下F1和F2产量性状的基因型值预测Table 6 Genotype values estimated of yield traits of F1and F2in different areas

(续表6)
3 小结与讨论
作物性状的遗传规律及亲本在杂交后代中的配合力表现一直是育种家最关心的,也是育种工作的核心。对甘蓝型油菜产量性状的遗传效应分析有利于清晰了解亲本材料的遗传行为,制定遗传改良方案或组合测配进行品种选育,从而提高育种的效率[16,17]。本研究采用加性—显性及其与环境互作遗传模型系统分析了亲本和杂种世代的遗传效应,在所研究的6个亲本材料、15个杂交组合中,4个产量性状杂种F1的表型值的均值均高于其亲本的表型值,说明甘蓝型油菜杂种F1的产量性状表现出了一定的正向杂种优势,同时环境条件对各性状的表型值影响又不同,如单株产量、角果数、角粒数大荔高于张掖,千粒重性状表现型值张掖高于大荔,说明存在着基因型与环境的互作现象,产量性状存在着基因型与环境的互作效应。在亲本加性效应分析中,4号和5号亲本可作为后代改良的理想亲本,在早期世代选择更有效;不同环境下甘蓝型油菜产量性状在不同世代与环境互作效应差异较大,单株产量、角果数和角粒数的F1表型值高于F2,但千粒重性状却与之相反,说明环境条件对千粒重性状的影响较大,在后代分离群体中千粒重受到较高的加性遗传,具有一定的杂种优势,因此千粒重需在特定环境中进行早期选择,这与沈金雄等人在油菜后代杂种优势的研究结果一致[18,19]。
在组合测配或品种改良中,既要注重一般配合力大小又要考虑其特殊配合力大小,因此亲本材料的加性遗传效应与具体组合的显性遗传效应在测配组合时就是选择的依据[20,21]。不同环境条件下,基因的表达有差异,因此,对甘蓝型油菜产量性状的遗传分析需要在多个环境中进行,探索出最有利于基因表达的环境条件,为改良材料聚合优良基因提供理论[22]。根据性状的遗传表现及与环境的互作,可有根据的进行选择与淘汰,这样有利于加快育种的进程,同时可缩小试验的规模,减少人为的误差。
参考文献:
[1] 官春云.双低油菜核心竞争力的研究[J].作物研究,2004,18(2):88 -93.
[2] 涂金星,张冬晓,张 毅,等.我国油菜育种目标及品种审定问题的商榷[J].中国油料作物学报,2007,29 (3):350 -352.
[3] 陈兆波,余 健.我国油菜生产形势分析及科研对策研究[J].中国油料作物学报,2010,32(2):303 -308.
[4] 邓武明,阳小虎,文凤君,等.甘蓝型油菜产量性状的遗传及相关与通径分析[J].中国油料作物学报,2003,25(4):27 -30.
[5] 李少钦,王健胜,张文学,等.甘蓝型油菜优良亲本对杂种后代产量性状的遗传效应分析[J].中国油料作物学报,2011,33(6):545 -549.
[6] 王 瑞,李加纳,唐章林,等.甘蓝型黄籽油菜产量性状的遗传分析[J].中国农学通报,2004(5):37 -38.
[7] 李泽炳.杂交水稻的应用与实践[M].上海:上海科学技术出版社,1981.
[8] 戚存扣.甘蓝型油菜主要农艺品质性状遗传体系与杂种优势改良研究[D].南京:南京农业大学博士学位论文,2001.
[9] 王健胜,侯桂玲,李少钦,等.甘蓝型油菜育种亲本产量及品质性状遗传效应分析[J].江苏农业学报,2015,31(3):489 -493.
[10]朱 军.遗传模型分析方法[M].北京:中国农业出版社,1996.241.
[11]朱 军.估算遗传方差和协方差的混合模型方法[J].生物数学学报,1992,7(1):1 -11.
[12]朱 军.作物杂种后代基因型值和杂种优势的预测方法[J].生物数学学报,1993,8(1):32 -44.
[13]朱 军,季道藩,许馥华.作物品种间杂种优势遗传分析的新方法[J].遗传学报,1993,20(3):262 -271.
[14]朱 军.广义遗传模型与数量遗传分析新方法[J].浙江农业大学学报,1994,20(6):551 -559.
[15]Zhu J,Weir BS.Diallel analysis for sex - linked and maternal effects[J].Theoretical and Applied Genetics,1996,92(1):1 -9.
[16]师家勤.甘蓝型油菜产量性状及其杂种优势遗传基础的全基因组解析[D].武汉:华中农业大学博士学位论文,2009.
[17]蔡东芳.甘蓝型油菜产量和品种相关性状关联分析[D].武汉:华中农业大学博士学位论文,2013.
[18]沈金雄,傅廷栋,杨光圣,等.甘蓝型油菜杂种优势及产量性状的遗传改良[J].中国油料作物学报,2005,27(1):5 -9.
[19]张书芬,马朝芝,朱家成,等.甘蓝型油菜主要农艺和产量性状的杂种优势及其分离世代的分析[J].中国油料作物学报,2007,29(2):121 -125.
[20]邹晓芬,邹小云,宋来强,等.甘蓝型油菜产量及其相关性状的配合力及遗传分析[J].江西农业学报,2007,19(2):1 -4.
[21]刘绚霞,董振生,刘创社,等.甘蓝型优质杂交油菜主要农艺性状配合力与遗传力研究[J].中国油料作物学报,2001,23(3):2 -5.
[22]董育红,关周博,侯君利,等.春油菜区甘蓝型油菜含油量的杂种优势及配合力研究[J].西北农业学报,2014,23(4):40 -44.
Genetic Analysis for Yield Traits of Brassica napus L.
GUAN Zhoubo,DONG Yuhong,LI Shaoqin,ZHANG Yaowen,TIAN Jianhua,LI Dianrong
(Hybrid Rapeseed Research Center of Shaanxi Province/ Shaanxi Rapeseed Branch of National Oil Crops Genetic Improvement Center,Yangling,Shaanxi 712100,China)
Abstract:To explore genetic effect of rapeseed parents and hybrid varieties in different ecological area,providing the theory base for breeding.This study analyzed genetic regularity of yield traits of different ecological area parents and hybrid combinations by additive dominant and the environment interaction genetic model.Estimated additive genetic effect value of parents yield traits,dominance genetic effect variance value of parents in hybrid combinations,the ratio of the variance components and genotype and environment effect value of F1and F2.The results showed that yield traits were affected by the additive and dominant genetic;In genetic effect and environmental interactions,all the genetic effects of yield per plant and pod number per plant and thousand grain weight achieved significant with environmental interactions,but except pod grain number.Rapeseed yield traits showed bigger different of F1and F2generation in different environment,overall performance that genotype and environment interaction value Dali was higher than Zhangye,but thousand grain weight was different,which offspring phenotype was bigger affected by environmental conditions.
Keywords:Brassica napus L;yield trait;genetic analysis
中图分类号:S565.403.2
文献标识码:A
文章编号:1001-5280(2016)03-0266-05
DOI:10.16848/ j.cnki.issn.1001-5280.2016.03.08
收稿日期:2016- 02- 26
作者简介:关周博(1983 -),男,硕士,主要从事油菜育种研究,Email:guanzhoubo1983@163.com。
基金项目:杨凌示范区科技计划项目(2015NY -02,2015NY -17,2014NY -43);油菜遗传育种创新团队项目(2015KTC -21)。
