台风对上层海洋初级生产力和营养盐输运的影响❋
2016-06-28潘珊珊高会旺颜秀花
潘珊珊, 史 洁❋❋, 高会旺, 徐 昭, 颜秀花
(1.中国海洋大学环境与生态教育部重点实验室,山东 青岛 266100; 2.中国海洋大学物理海洋教育部重点实验室,山东 青岛 266003; 3.国家海洋局第三海洋研究所海洋-大气化学与全球变化重点实验室,福建 厦门 361005)
台风对上层海洋初级生产力和营养盐输运的影响❋
潘珊珊1, 史洁1❋❋, 高会旺1, 徐昭2, 颜秀花3
(1.中国海洋大学环境与生态教育部重点实验室,山东 青岛 266100; 2.中国海洋大学物理海洋教育部重点实验室,山东 青岛 266003; 3.国家海洋局第三海洋研究所海洋-大气化学与全球变化重点实验室,福建 厦门 361005)
摘要:利用垂直一维物理-生物耦合模型模拟了台风“派比安”和超强台风“珍珠”对南海北部水温、营养盐和叶绿素垂直分布的影响,并估算了2次台风对初级生产力和营养盐垂直输送的贡献。结果表明,“派比安”引发50m以浅海水温度降低,表层降温2.0℃,50~130m海水温度升高,混合层加深30m,海表叶绿素浓度增加0.18mg·m-3,营养盐垂向输运对初级生产力的贡献为2.3×103mg C·m-2,约占全年的3%。“珍珠”引发55m以浅海水温度降低,表层降温超过5.0℃,55~150m海水温度升高,混合层加深85m,海表叶绿素浓度增加0.9mg·m-3,带来的营养盐垂向输运量约为全年的30%,对初级生产力的贡献为12.8×103mg C·m-2,约占全年的18%。可见,台风过程,特别是强台风过程对上层海洋的初级生产和生物地球化学过程具有显著影响。台风的强度和移动速度等自身特征是决定海洋环境要素对台风响应程度的核心要素,同时台风过境前的水体层化状态和营养盐水平也是不可忽视的因素。
关键词:南海北部; 台风; 浮游植物水华; 初级生产力; 物理-生物耦合模型
引用格式:潘珊珊, 史洁, 高会旺,等. 台风对上层海洋初级生产力和营养盐输运的影响[J]. 中国海洋大学学报(自然科学版), 2016, 46(6): 120-133.
PAN Shan-Shan, SHI Jie, GAO Hui-Wang, et al. Impacts of typhoon on ocean primary production and nutrients transport[J]. Periodical of Ocean University of China, 2016, 46(6): 120-133.
台风是发生在热带或副热带海洋上的一种具有暖心结构的强烈气旋性涡旋,是发生在海洋上的强烈天气系统,是海气相互作用的直观表现[1]。台风引起的水体强烈垂直混合对上层海洋与深层海水之间的热量、能量和物质交换有重大影响[2]。台风过境可以加深海洋上混合层深度,通过卷挟作用将次表层冷水带到表层,引起海表温度降低及次表层温度升高[3-6]。伴随台风产生的大风混合、波浪混合、波浪破碎搅拌和上升流等动力过程,能够使深海大洋下层的营养盐穿过跃层进入寡营养盐的上层海洋[2,7],从而促进上层海洋浮游植物生长,使得叶绿素浓度升高[8-10],提高海洋初级生产力水平[11-13]。
目前,有关海洋生态系统对台风响应的研究方法包括有定点浮标观测[3,14-16]、卫星遥感观测[17-21]和数值模拟方法[22-25],以及观测与数值模拟相结合的方法[11,13,16]。台风期间的直接船基观测存在很大困难,而基于卫星观测的研究主要揭示了台风过境引起的海水表层温度降低以及叶绿素浓度增加,无法刻画表层以下海水物理及生物要素的响应。Zheng和Tang[11]利用卫星观测资料,通过对比台风“达维”(2005年9月)过境前后南海表层水体的叶绿素浓度分布,发现台风在近岸和远海2个不同海域均引发了浮游植物水华,但是二者的产生机制有所不同。远海水华主要是与台风引起的垂向混合以及上升流有关,而近岸水华则可能是台风带来的降雨增加地表径流向海洋输入营养盐导致。Zhao等[17]利用卫星遥感数据,对比分析了“玲玲”(2001年11月)和“启德”(2005年10—11月)2次台风对南海北部生态系统的影响,指出台风过境引起海表面叶绿素浓度升高,伴随着海表温度下降,海表面高度异常,并且叶绿素最大值出现的时间滞后于风速最大值5~12 d。基于数值模型的研究多数集中在分析台风过境时海水温度等物理要素垂直结构的变化,侧重大气与海洋之间的热力与动力过程。崔红等[26]采用POM模式研究了台风“伊布都”(2003年7月)过境期间南海物理环境的响应,结果表明:台风过后,海表温度下降2~6℃,混合层加深10~60m,混合层以下附近水层处于增温状态。而少有数值模型研究涉及营养盐和浮游植物生物量等生物要素垂直结构对台风的响应。
中国南海是一个半封闭性海盆,面积约3.5×106km2,平均水深约1200m,海盆水深最深处超过5000m,是世界上最大的边缘海之一[27-28](见图1)。南海大部分海域属于热带海区,太阳辐射较强,海水层化现象常年存在,阻碍了营养盐从深层向真光层的补充,上层海洋全年均呈现寡营养的状态。叶绿素浓度垂直分布呈现典型的次表层最大值现象[29-31]。南海台风活动频繁,研究表明,每年约有10次热带气旋经过南海[32],且从20世纪90年代—21世纪比20世纪70—90年代初有明显的增加趋势[33]。因此,研究南海环境要素对于台风过程的响应,进而分析台风影响下的海洋初级生产力变化具有非常重要的科学意义。
本文选取南海北部长期观测站SEATS站(18°N,116°E)(见图1)为研究对象,首先利用垂直一维物理-生物耦合模型对该站海水温度、盐度、营养盐及叶绿素浓度垂直结构的季节变化进行了模拟,并用该站长期的船载观测资料进行验证。在此基础上,利用物理-生物耦合模型,并结合卫星资料,分析了2006年台风“派比安”和超强台风“珍珠”过境时南海SEATS站物理及生物要素的响应过程,定量估算了2次台风对SEATS站营养盐跨跃层输运和初级生产力的贡献。最后,讨论了两次台风产生不同影响程度的原因。
1模型及数据介绍
1.1 模型介绍
本研究采用的垂直一维物理-生物耦合模型由2个模块组成。其中,物理模块采用一维POM模型(Princeton Ocean Model)[34],生物模块采用简化的欧洲区域海洋生态模型ERSEM(European Regional Sea Ecosystem Model)[35,36]。物理模块为生物模块提供温度和湍混合系数,生物模块进行营养盐、叶绿素、浮游动物和浮游细菌的浓度等生态变量的计算。
由于POM模型采用的是传统的二阶Mellor-Yamada湍流闭合模型,很多研究发现其模拟出的上混合层较浅[37],前人研究中考虑了特征波参数化下的海浪混合作用[38-40]。胡浩国等[41]在模型垂直混合系数的计算中采用了同样的方案,较好地模拟了渤海、黄海和东海等中国近海混合层季节变化规律。本文物理模块部分采用该修改方案,把海浪引起的垂直湍粘性系数和垂直湍混合系数Kwm和Kwh叠加到Mellor-Yamada湍封闭模型计算的垂直混合系数Km和Kh上。Kwm和Kwh可表示为:
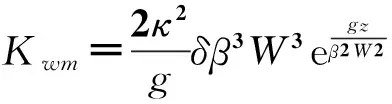
(1)
Kwh=PKwm。
(2)
其中:κ=0.4为Von Karman常数;g=9.8m·s-2为重力加速度;δ=0.1为波陡;β=0.65为波龄[42];W为海表风速(m·s-1);z为水深(m),以向上为正;P为与Richardson数有关的无量纲系数,表示热混合系数与动量混合系数(即湍黏性系数)的比值,参考文献中的取值[38-41],取P=0.1。混合系数的最小值取为1×10-5m2·s-1。
生物模块中包含9个状态变量(见图2):2种营养盐(溶解性无机氮(DIN:dissolved inorganic nitrogen)和溶解性无机磷(DIP:dissolved inorganic phosphate))、1种浮游植物、3种浮游动物(杂食中型浮游动物、微型浮游动物和异养鞭毛藻)、2种有机物(溶解性有机物(DOM:dissolved organic matter)和颗粒态有机物(POM:particulate organic matter))以及浮游细菌。浮游植物光合作用吸收营养盐,被杂食中型和微型浮游动物摄食;浮游细菌摄食有机物(DOM和POM),被异养鞭毛藻摄食;杂食中型浮游动物位于最高营养级,以浮游植物和微型浮游动物为食;浮游植物、浮游细菌和微型浮游动物排泄产生的DOM,被浮游细菌摄食;浮游植物和3种浮游动物排泄产生POM;DIN和DIP通过DOM和POM的矿化溶解实现再循环。模型参数的选取依据南海及邻近海域的研究结果和ERSEM模拟中经常使用的参数。具体参数取值见表1~2。
1.2 模型配置和数据来源
模拟水深取为南海平均水深1200m,代表常年层化的南海海盆区。垂向分为40层,由于重点关注上层海洋生态系统各要素的变化,故150m以浅采用上密下疏非均匀分为35层,而150m以深均匀分为5层。时间步长为216s。模拟实验包括气候态季节变化的模拟和2006年典型台风事件影响的模拟。气候态模拟的主要目的是再现南海SEATS站生态系统各要素的季节变化,对模型进行验证,并且为2006年的模拟提供初始场。
气候态模拟采用1981—2010年30年平均的月均海表强迫数据。海表风速取自NCEP月均海表10m风速数据。海表面净热通量包括太阳短波辐射、有效长波回辐射、蒸发潜热和感热。其中短波辐射和长波辐射数据来自NCEP再分析资料,潜热和感热通量采用块体动力学方法(COARE算法)根据气温、海表温度、相对湿度、海表气压和风速计算得到[45]。海表盐度强迫数据取自NCEP月均海表盐度数据。温度、盐度和营养盐初始场取自WOA2013的1月份的垂直剖面数据,浮游植物、浮游细菌、浮游动物和有机物(POM和DOM)的初始浓度均赋零。模型从1月1日开始在相同强迫下运行3年,选取第3年的模拟结果与观测资料进行对比验证,海水温度、盐度和叶绿素浓度的观测数据取自SEATS站船载观测资料[46],营养盐的观测数据取自WOA2013气候态平均的垂直剖面数据。
2006年的模拟采用的初始场为气候态模拟第3年1月1日的结果。风和热通量海表强迫取自2006年NCEP 6h 1次的海表数据。用来验证模型结果的海表温度数据取自TRMM Microwave Imager(TMI)卫星资料,海表叶绿素浓度数据取自Moderate Resolution Imaging Spectroradiometer(MODIS)和Sea-viewing Wide Field-of-view Sensor(SeaWiFS)Level 3产品,时间分辨率为1d 1次,空间分辨率为9km。卫星数据均为以SEATS站为中心2(°)×2(°)区域(17°N~19°N,115°N~117°E)的平均值。台风路径数据,即每6h1次的台风中心位置取自日本气象厅(Japan Meteorological Agency, JMA)的最佳路径数据。台风移动速度是根据每6h 1次的台风中心位置计算得到的。
1.3 2006年过境SEATS站的台风概况
2006年西太平洋共发生了23次热带气旋,过境南海的有9次,其中超强台风“珍珠”(0601)、台风“派比安”(0606)以及超强台风“西马仑”(0619)路径接近SEATS站(见图1,图中出现的时间均为世界时间,UTC)。但“西马仑”过境期间,NCEP反演SEATS站风速资料并未出现高值,而“珍珠”和“派比安”过境时SEATS站风速明显增大。故本研究选取“珍珠”和“派比安”两次台风影响下的模拟结果进行对比分析。
超强台风“珍珠”(0601,按照萨菲尔-辛普森(Saffir-Simpson)等级为4级台风)是1949年以来5月份登陆中国的最强台风之一。2006年5月9日06:00生成于菲律宾东南方向西太平洋洋面上,12:00加强为热带风暴,随后向西北偏西方向移动。5月13日00:00穿过菲律宾群岛后进入南海中南部,加强为台风并向西移动,5月15日00:00发展为强台风,台风中心在南海中部海面转向偏北方向移动,从SEATS站左侧穿过。5月17日00:00起移动路径转向东北,5月18日在广东省登陆,随后继续北偏东移动穿过福建省,最后在东海减弱消失。本次台风持续时间达10d(5月9—18日),平均移动速度为4.4m·s-1。台风“珍珠”过境期间SEATS站的海表风速5月9日为5m·s-1左右,随着台风不断前行,到5月15日缓慢增加至25m·s-1,随着台风继续北移,5月18日海表风速减弱至5m·s-1(见图3(b),图中灰色区域代表台风持续时间)。
台风“派比安”(0606,1级)于2006年7月31日00:00在菲律宾以东的太平洋洋面上生成,向西移动并穿过菲律宾北部。8月1日进入南海,并增强为热带风暴,继续向西北偏西移动,18:00经过SEATS站。8月3日转向西北移动,在广东西部登陆。持续时间为4d(7月31日—8月3日),平均移动速度为5.8m·s-1。台风“派比安”过境期间SEATS站的海表风速7月31日为5m·s-1左右,随着台风不断前行,8月2日迅速增加至22m·s-1,8月3日减弱至9m·s-1(见图3(c))。
2结果
2.1 物理及生物要素季节变化模拟结果与验证
模拟得到的SEATS站海水温度、盐度、营养盐和叶绿素浓度垂直剖面的季节变化与SEATS站长期观测资料和WOA2013数据对比可以看出,本模型较好地再现了SEATS站物理和生物要素气候态的季节变化规律(见图4)。
南海位于东亚季风区,每年6—9月盛行暖湿的西南季风(夏季风),平均风速6m·s-1,11月至次年3月盛行干冷的东北季风(冬季风),平均风速10m·s-1,春秋为季风转换期(见图3(a))。南海生态系统物理和生物要素季节变化受季风影响显著。本文定义温度比表层10m平均温度低1.0℃的深度为混合层深度[47]。模拟结果显示,冬季在强盛的冬季风影响下,混合层较深,可达80m左右。春季,随着太阳辐射不断增强,风速变小,海表面开始增温,上层海水出现层化现象,温跃层浅于20m。7月表层海水温度达到最大值,约为30.0℃。之后,由于太阳辐射减弱,表层海水温度降低,加之风速增大使得混合层逐渐加深(见图4(a))。
南海SEATS站营养盐浓度垂直结构无明显季节变化,上层50m以浅营养盐分布均匀,50m处为营养盐跃层的位置,50m以下浓度随深度迅速增加。除冬季以外的其他季节,上层50m以浅营养盐浓度很低(DIN<1.0mmol·m-3、DIP<0.1mmol·m-3),甚至低于检出限和浮游植物生长的最低阈值[48],限制了浮游植物生长。冬季风引起的较强垂向湍混合将深层富营养水体带至上层,使得营养盐浓度略微升高(见图4(c)~(d))。
(a.2006年和气候态平均风速;b.台风“珍珠”过境期间风速;c.台风“派比安”过境期间风速。a.Wind speed in 2006 and climatological wind speed; b.Wind speed during typhoon Chanchu; c.Wind speed during typhoon Prapiroon.)
图3风速随时间的变化
Fig.3Time series of wind speed
叶绿素浓度垂直分布的季节变化是温度、光和营养盐等要素共同作用的结果(见图4(e))。真光层深度采用1%表面光强的深度,其季节变化很小,基本在
90m深度左右。除冬季以外的季节,由于真光层以上营养盐匮乏,而真光层以下又受到光的限值,叶绿素浓度在垂向上出现次表层最大值现象,位于约60m,其浓度约为0.40mg·m-3。此时,海表叶绿素浓度低于0.10mg·m-3。冬季,下层营养盐被带至寡营养的上层,支持了浮游植物生长,海表叶绿素浓度升高,达到全年最高值0.30mg·m-3,次表层叶绿素浓度最大值现象消失。
2.2 温度的垂直分布对台风事件的响应
通过2006年海表温度模拟结果可以看出,海表温度的季节变化呈现冬季低夏季高的基本特征,并且2次台风过境期间有明显的海表降温现象,与TMI卫星观测结果吻合(见图5)。台风“珍珠”过境之前(5月9日)海表温度超过29.0℃,5月15日降至24.0℃,海表最大降温超过5.0℃;台风“派比安”过境之前(7月31日)海表温度超过29.0℃,8月2日降至27.0℃,最大降温超过2.0℃。
台风过境引起的强列垂直湍混合作用能够破坏温度跃层结构。从温度的垂直结构变化可以看出(见图6),台风“珍珠”过境之前(5月9日)混合层深度为15m,5月12日风速开始增强,混合层深度也逐渐加深,到5月15日已达到100m。各层海水温度呈现出不同的变化特征(见图7),55m以浅海水温度降低,最大降温发生在海洋表层;55~150m温度升高,如台风过境后100m处海水温度较之前上升了2.0℃。台风过境之后,由于海表加热,海表温度和混合层深度逐渐恢复。台风“派比安”过境之前(7月31日)混合层深度为45m。8月1日起风速开始增强,混合层深度也开始加深,到8月2日达到75m。50m以浅海水温度降低,最大降温发生在表层;50~130m温度升高,如100m水深海水温度在台风过境之后上升了0.7℃。台风过境之后,海表温度和混合层深度开始恢复。台风“珍珠”引起的各层温度变化幅度和影响的深度均比“派比安”大。
2.3 营养盐浓度的垂直分布对台风事件的响应
台风期间,增强的垂直混合将下层富营养盐海水带到真光层以上,使海表营养盐浓度升高(见图8),同时也改变了营养盐浓度的垂直结构(见图9)。模拟结果显示,台风“珍珠”过境之前(5月9日)海表DIN和DIP的浓度分别为1.08和0.09mmol·m-3;5月15日浓度分别增加到3.75和0.22mmol·m-3,分别为台风过境之前的3.3和2.4倍。台风“派比安”过境之前(7月31日)海表DIN和DIP的浓度分别为1.71和0.08mmol·m-3;8月2日分别增加到2.45和0.13mmol·m-3,是过境之前的1.4和1.6倍。台风期间向真光层的营养盐补充为浮游植物水华提供了必要物质基础。台风过后,由于浮游植物生长消耗和下层补充减弱,营养盐浓度逐渐降低并恢复到台风过境之前的水平。
2.4 叶绿素浓度的垂直分布对台风事件的响应
图10为2006年海表叶绿素浓度的模拟结果和卫星资料对比,结果表明,海表叶绿素浓度季节变化呈现冬季高夏季低的基本特征。在台风“珍珠”和“派比安”过境之后模拟结果显示有明显的海表叶绿素浓度增加现象,其增加幅度与MODIS和SeaWiFS卫星叶绿素数据吻合较好。“珍珠”过境前(5月9日),海表叶绿素浓度仅为0.12mg·m-3;过境期间(见图10:灰色区域),其值逐渐升高;直至台风过境,海表风速开始下降时,海表叶绿素浓度才达到最大值1.02mg·m-3,约为台风过境前的10倍;之后,其值逐渐降低至台风前的水平。“派比安”过境之前(7月31日),海表叶绿素浓度为0.13mg·m-3,与“珍珠”过境之前相当;随着台风影响的发展,海表风速逐渐增大,海表叶绿素浓度持续升高;直至台风过境之后,海表叶绿素浓度达到其最大值0.31mg·m-3,约为台风过境之前的3倍;之后逐渐降低至台风前的水平。
台风过境不仅引起了海表叶绿素浓度的升高,也改变了叶绿素浓度的垂直分布(见图11)。台风“珍珠”过境前(5月9日),垂向叶绿素浓度最大值为0.40mg·m-3,位于60m水层,呈现出南海春季次表层叶绿素浓度最大值的典型垂直分布特征[29-30,43]。随着风速的增大,强烈的垂直混合将位于次表层的浮游植物卷挟至表层和真光层以下。5月15日,风速达到最大,海表叶绿素浓度增加到0.25mg·m-3;真光层以下叶绿素浓度也有所升高,如100m深度处台风过境之前叶绿素浓度几乎为零,此时增加到0.15 mg·m-3,上层水体
中叶绿素浓度达到均匀,次表层叶绿素浓度最大值现象消失。之后,虽然海表风速逐渐减小,但真光层内叶绿素浓度继续增加,5月22日海表叶绿素浓度达到其最大值,滞后于风速最大值7d,且整个60m以浅的水层都出现了明显的浮游植物水华。5月23日40m水深叶绿素浓度达到其最大值1.23mg·m-3,是台风过境前的8倍多,达到最大值的时间比表层滞后1d。6月2日60m水深叶绿素浓度达到其最大值0.63mg·m-3,比表层滞后约10d。而被带至真光层以下的那部分浮游植物由于缺乏光照,逐渐死亡,叶绿素浓度持续降低。台风过后,上层营养盐被大量消耗,风速的降低也导致垂直混合变弱,营养盐跃层出现,次表层叶绿素浓度最大值现象再次形成。台风“派比安”过境前(7月31日),同样存在次表层叶绿素浓度最大值现象,位于60m水层,叶绿素浓度为0.38mg·m-3。8月2日,风速达到最大,上层水体中叶绿素被混合均匀,次表层叶绿素浓度最大值现象消失。海表叶绿素浓度增加为0.22mg·m-3;真光层以下叶绿素浓度也有所升高,如100m深度处台风过境之前叶绿素浓度几乎为零,此时增加到0.10mg·m-3。之后,虽然海表风速逐渐减
小,但真光层内叶绿素浓度继续增加,海表叶绿素浓度在8月8日达到其最大值0.31mg·m-3,滞后于风速最大值6d,且整个60m以浅的水层都出现了浮游植物水华。而被带至真光层以下的那部分浮游植物逐渐死亡,叶绿素浓度降低。台风过后,次表层叶绿素浓度最大值现象再次形成。与“珍珠”相比,“派比安”引起的叶绿素浓度变化幅度较小。
3讨论
3.1 台风事件对营养盐和初级生产力贡献的估算
基于模型结果,本文估算了由垂直混合作用引起的向真光层补充营养盐的量。结果表明,2006年全年DIN和DIP垂向输入真光层的量分别为692.74和38.21mmol·m-2。台风“珍珠”过境期间(5月9—18日)该量为192和9.9mmol·m-2,分别占全年总量的28%和26%。台风“派比安”过境期间(7月31日—8月3日)则为19.27和1.05mmol·m-2,约占全年2.8%和2.7%。在南海,还未有研究对台风引起的营养盐向真光层的补充量进行过计算,但在东海Hung等[7]通过船载观测数据估算得出2009年8月过境东海南部的台风“莫拉克”所贡献的DIN和DIP约占整个夏季的86%和87%。可见,台风过境对寡营养海区营养盐供给具有重要意义。
海洋初级生产力为海洋浮游植物通过光合作用或化学合成制造有机物的速率,通常以每天或每年单位面积上产生的碳量表示。本文将真光层内浮游植物净生长的积分值代表初级生产力(Integrated Primary Production(IPP),单位:mg C·m-2d-1或mg C·m-2a-1)。模拟结果显示,2006年SEATS站年初级生产力为71.9×103mg C·m-2a-1,平均初级生产力为200mg C·m-2d-1。季节变化上来看,最小值128mg C·m-2d-1出现在夏季,最大值280mg C·m-2d-1出现在冬季。Tseng等[49]估算南海SEATS站初级生产力在冬季为300mg C·m-2d-1,其他季节约保持在110mg C·m-2d-1的水平。Liu和Chai[50]用模型计算南海初级生产力年际变化不大,其范围在150~220mg C·m-2d-1之间。模型结果与以往研究大致相当。
台风“珍珠”过境之前5月9日初级生产力仅为130mg C·m-2d-1,5月12号开始迅速增大,到5月22日增大到1240mg C·m-2d-1,约为过境之前的10倍,6月10日前后初级生产力回到台风过境之前的水平。这期间(5月9日—6月10日)初级生产力总量为16.6×103mg C·m-2,而气候态模拟结果在这期间的积分结果仅为3.8×103mg C·m-2,故可以看出台风“珍珠”过境引起的浮游植物水华可贡献初级生产力12.8×103mg C·m-2,约为全年初级生产力的18%。台风“派比安”过境之前初级生产力为132mg C·m-2d-1,台风过境之后初级生产力最大可达289mg C·m-2d-1,8月底返回到台风过境前的水平,台风影响期间(7月31日—8月30日)初级生产力积分值为6.0×103mg C·m-2,气候态结果在这期间的积分结果为3.7×103mg C·m-2,可以看出台风“派比安”过境引起的浮游植物水华约贡献初级生产力2.3×103mg C·m-2,约为全年的3%。2次台风引起的浮游植物水华可贡献全年初级生产力的21%。在东海,台风“米雷”(2004年9月)的过境引起初级生产力增加了3倍,贡献全年新生产力的3.8%[16]。Naik等[51]估算在阿拉伯海过境的一次热带气旋贡献其初级生产力5%。Lin等[12]指出每年热带气旋经过南海,大约贡献南海新生产力的30%。2007年第24号台风“海贝思”在南海中部引起了强烈的浮游植物水华,海表叶绿素浓度增加了20多倍,最高可达5.0mg·m-3,可贡献年初级生产力的30%[52]。可见,本研究中台风事件对海洋初级生产力贡献的估算结果与南海及其他海域的以往研究具有可比性,说明在南海等热带海洋中台风对提高海洋初级生产力具有重要意义。
3.2 南海北部环境要素对台风响应程度的影响因素
由以上结果分析可以看出台风“珍珠”和“派比安”对SEATS站物理及生物要素的影响程度是不同的。表3给出了2次台风各自的特征以及引起的SEATS站物理和生物要素的变化情况对比。从表中可以明显看出,台风“珍珠”的影响要远大于“派比安”,这是由2次台风的强度、移动速度等自身特征及台风发生的季节或者说过境之前水体条件(如混合层深度和营养盐水平等)的不同所导致。为了定量研究南海北部环境要素对台风响应程度的2个影响因素(台风的自身特点和发生的季节),本研究设计了一个模型实验,将8月份台风“派比安”过境期间的海表强迫换为台风“珍珠”过境期间的海表强迫进行模拟,将模拟结果与采用2006年实际海表强迫模拟得到的结果进行对比分析,以定量化研究其影响机制。模拟得到的海表温度和海表叶绿素浓度如图12和13所示,引起的SEATS站物理和生物要素的变化情况见表3,可以看到,若“珍珠”发生在与“派比安”同样的时间,可引起海表温度降低大约4℃,混合层加深58m,海表叶绿素浓度增加0.62mg·m-3,贡献初级生产力9.0×103mg C·m-2。
不同强速和移动速度的台风对海洋生态系统各要素的影响是不同的。台风“珍珠”移动速度较慢,其等级也高于“派比安”,因此无论是其强度还是持续时间均比“派比安”强,在相同的前期水体条件下,对生态系统的影响程度必然要大于“派比安”。比较“派比安”模拟和数值实验的结果可以看出,若台风“珍珠”发生在和“派比安”相同的时间,即当台风过境之前的水体条件相同而台风的特点不同(见表3),可以看出强度大和移动速度慢的台风引起的各要素的响应程度以及对营养盐垂直输运和初级生产力的贡献均大于强度小和移动速度快的台风。Bender等[53]的研究表明,移动速度较慢或者较强的台风所引起的海表降温幅度比移动速度较慢和较弱的台风更大,这是因为移动速度慢的台风持续时间更长。因此不同强速和移动速度的台风对浮游植物生物量和初级生产力的影响也是不同的。Zhao等[17]用卫星数据对比研究了相同季节过境南海的两次不同强度和移动速度的台风对海洋生态系统的影响,移动速度较慢的台风“启德”(2005年10—11月)对海洋的影响时间较长,引起的浮游植物水华更强,而强度较强的台风“玲玲”(2001年11月)引起的海表降温和浮游植物水华区域的面积更大。
2个台风发生的季节不同也导致了其对海洋的影响不同。“珍珠”发生在5月(春季),并且在此之前没有台风经过SEATS站,混合层深度比较浅,仅为15m;而“派比安”发生在8月份(夏季),此次台风过境之前,SEATS站有几次明显的大风事件,使得混合层深度比较深,为45m。所以,就台风前的水体条件来说,“珍珠”过境之前的条件比“派比安”更容易加深混合层深度、打破营养盐跃层和提高叶绿素浓度。加之“珍珠”强度远大于“派比安”,其对SEATS站环境要素的影响也较为明显。结果表明,“珍珠”过后,水体混合层深度加深至100m,加深了85m,海表营养盐浓度增大为台风前的2~4倍,海表叶绿素浓度和水柱初级生产力约增大为台风前的10倍;而“派比安”仅使混合层加深了30m,营养盐浓度约增大为台风前的1.5倍,海表叶绿素浓度和水柱初级生产力约为台风前的3倍。比较“珍珠”模拟和数值实验的结果,可以看出当同一个台风发生在不同季节或者说其过境之前的水体条件不同时(见表3),对海洋的影响也是不同的,发生在混合层深度较浅的春季时引起的各要素的响应程度均大于发生在混合层较深的夏季引起的响应程度。以往研究显示,超强台风“玲玲”(0123),为4级台风,平均移动速度4.3m·s-1,于2001年11月经过南海中部,由于当时强冬季风使得南海上层水体混合已较为均匀,此次台风使海表叶绿素浓度从0.14mg·m-3增加到0.45mg·m-3,仅提高了3.2倍[17]。强台风“达维”(0518),为2级台风,平均移动速度也小于“玲玲”,为3.9m·s-1,于2005年9月过境南海北部,过境前海表面风速较小,海水层结稳定,上混合层较浅,台风使海表叶绿素浓度从0.12mg·m-3增加到0.62mg·m-3,提高达6倍之多[11]。而台风“启德”(0004)仅为1级台风,平均移动速度4.0m·s-1,于2000年7月过境南海北部,由于过境之前混合层和营养盐跃层位置更浅,此次台风引起海表叶绿素浓度增加了30倍之多[12]。此外,台风“珍珠”过境后,虽然海表太阳辐射可以使海表温度回升到台风前水平,但是台风期间强垂直混合将表层暖水带至55m以下的次表层,其温度很难恢复到台风过境之前的水平(见图6),生物要素的垂直结构也无法完全回到台风发生前的状态,这也将影响生态系统对其后台风的响应。所以,“珍珠”对SEATS站的影响将会削弱其后发生的“派比安”的作用。Yang等[6]研究了2005年过境南海北部的3次台风对上层海洋物理环境的影响,结果表明,由于发生在7月份的台风“天鹰”加深了混合层并且破坏了水体层结,使得海洋对发生在9月份的台风“达维”的响应明显弱于“天鹰”。可见,由台风发生的季节或者台风过境前物理和生物要素的垂直分布情况会影响上层海洋对台风的响应程度。
4结语
本文利用一维物理-生物耦合模型对南海北部SEATS站环境要素气候态的季节变化进行模拟,并用观测数据对模型结果进行了对比验证。在此基础上,结合卫星观测数据,研究了2006年过境南海SEATS站的台风“派比安”和超强台风“珍珠”对该站物理和生物要素的影响,对比分析了海水表层温度、营养盐和叶绿素浓度及其垂直结构的响应特征,估算了2次台风对营养盐垂向输运和初级生产力的贡献,并结合数值实验以及文献资料定量分析了南海北部环境要素对台风响应程度的影响因素。
台风“珍珠”过境引起强烈的湍混合,低温高营养盐的深层水被带到上层,造成55m以浅海水温度降低,表层降温超过5.0℃,55~150m海水温度升高,混合层深度从15m加深到100m;表层营养盐浓度可以达到过境前的2~4倍,叶绿素浓度可以达到过境前的10倍。“珍珠”过境期间向真光层营养盐的垂向输运量为192~9.9mmol·m-2,约占全年的30%;贡献水柱初级生产力为12.8×103mg C·m-2,约占全年的18%。台风“派比安”过境使50m以浅海水温度降低,表层降温2.0℃,50~130m温度升高,混合层深度从45m加深到75m,叶绿素浓度可以达到过境之前的3倍。“派比安”过境期间向真光层营养盐的垂向输运量为19.27和1.05mmol m-2;贡献水柱初级生产力为2.3×103mg·m-2,约占全年的3%。生态系统各要素对台风“珍珠”和“派比安”的不同响应程度取决于台风的自身特征、发生季节以及过境之前的水体条件等,且一次台风的过境加深了混合层和营养盐跃层位置,这也会削弱其后发生的台风对生态系统的影响。
参考文献:
[1]陈联寿, 丁一汇. 西太平洋台风概论[M]. 北京: 科学出版社, 1979.
Chen L S, Ding Y H. Introduction to the typhoon in the western pacific[M]. Beijing: Science Press, 1979.
[2]Tang D L, Sui G, Lavy G, et al. Typhoon impact and crisis management[M]. U.S.: Springer, 2014.
[3]Zenghong L, Jianping X U, Chaohui S, et al. An upper ocean response to Typhoon Bolaven analyzed with Argo profiling floats[J]. Acta Oceanologica Sinica, 2014(11): 90-101.
[4]Guan S, Zhao W, Huthnance J, et al. Observed upper ocean response to typhoon Megi (2010) in the northern South China Sea[J]. Journal of Geophysical Research: Oceans, 2014,119(5):3134-3157.
[5]Zhang Y, Tian J. Enhanced turbulent mixing induced by strong wind on the South China Sea shelf[J]. Ocean Dynamics, 2014, 64(6): 781-796.
[6]Yang B, Hou Y, Hu P, et al. Shallow ocean response to tropical cyclones observed on the continental shelf of the northwestern South China Sea[J]. Journal of Geophysical Research: Oceans, 2015,120(5): 3817-3836.
[7]Hung C, Chung C, Gong G, et al. Nutrient supply in the southern East China Sea after typhoon Morakot[J]. Journal of Marine Research, 2013, 71(1-2): 1-2.
[8]Zheng Z, Ho C, Zheng Q, et al. Satellite observation and model simulation of upper ocean biophysical response to Super Typhoon Nakri[J]. Continental Shelf Research, 2010, 30(13): 1450-1457.
[9]Liu X, Wang M, Shi W. A study of a Hurricane Katrina-induced phytoplankton bloom using satellite observations and model simulations[J]. Journal of Geophysical Research: Oceans (1978-2012), 2009, 114(C3):819-834.
[10]Shibano R, Yamanaka Y, Okada N, et al. Responses of marine ecosystem to typhoon passages in the western subtropical North Pacific[J]. Geophysical Research Letters, 2011, 38(18): 702-706.
[11]Zheng G M, Tang D. Offshore and nearshore chlorophyll increases induced by typhoon winds and subsequent terrestrial rainwater runoff[J]. Marine Ecology Progress Series, 2007, 333: 61-74.
[12]Lin I, Liu W T, Wu C C, et al. New evidence for enhanced ocean primary production triggered by tropical cyclone[J]. Geophysical Research Letters, 2003, 30(13): 51.
[13]Ye H J, Sui Y, Tang D L, et al. A subsurface chlorophyll a bloom induced by typhoon in the South China Sea[J]. Journal of Marine Systems, 2013, 128: 138-145.
[14]Shan H, Guan Y, Huang J. Investigating different bio-responses of the upper ocean to Typhoon Haitang using Argo and satellite data[J]. Chinese Science Bulletin, 2014, 59(8): 785-794.
[15]Liu Z, Xu J, Zhu B, et al. The upper ocean response to tropical cyclones in the northwestern Pacific analyzed with Argo data[J]. Chinese Journal of Oceanology and Limnology, 2007, 25: 123-131.
[16]Siswanto E, Ishizaka J, Morimoto A, et al. Ocean physical and biogeochemical responses to the passage of Typhoon Meari in the East China Sea observed from Argo float and multiplatform satellites[J]. Geophysical Research Letters, 2008, 35(15): 105-116.
[17]Zhao H, Tang D, Wang Y. Comparison of phytoplankton blooms triggered by two typhoons with different intensities and translation speeds in the South China Sea[J]. Mar Ecol Prog Ser, 2008, 365: 57-65.
[18]Babin S M, Carton J A, Dickey T D, et al. Satellite evidence of hurricane-induced phytoplankton blooms in an oceanic desert[J]. Journal of Geophysical Research: Oceans (1978-2012), 2004, 109(C3): 325-347.
[19]Tang D L, Kawamura H, Doan Nhu H, et al. Remote sensing oceanography of a harmful algal bloom off the coast of southeastern Vietnam[J]. Journal of Geophysical Research: Oceans (1978-2012), 2004, 109(C2): 325-347.
[20]Walker N D, Leben R R, Balasubramanian S. Hurricane-forced upwelling and chlorophyll a enhancement within cold-core cyclones in the Gulf of Mexico[J]. Geophysical Research Letters, 2005, 32(18): 241-248.
[21]Zhao H, Han G, Zhang S, et al. Two phytoplankton blooms near Luzon Strait generated by lingering Typhoon Parma[J]. Journal of Geophysical Research: Biogeosciences, 2013, 118(2): 412-421.
[22]Diaz-Hernandez G, Mendez F J, Mínguez R. Numerical analysis and diagnosis of the hydrodynamic effects produced by hurricane Gordon on the coast of Spain[J]. Weather and Forecasting, 2014, 29(3): 666-683.
[23]Seo H, Xie S. Impact of ocean warm layer thickness on the intensity of hurricane Katrina in a regional coupled model[J]. Meteorology and Atmospheric Physics, 2013, 122(1-2): 19-32.
[24]Tsai Y, Chern C, Wang J. Numerical study of typhoon-induced ocean thermal content variations on the northern shelf of the South China Sea[J]. Continental Shelf Research, 2012, 42: 64-77.
[25]Wada A, Uehara T, Ishizaki S. Typhoon-induced sea surface cooling during the 2011 and 2012 typhoon seasons: observational evidence and numerical investigations of the sea surface cooling effect using typhoon simulations[J]. Progress in Earth and Planetary Science, 2014, 1(1): 1-25.
[26]崔红, 张书文, 王庆业. 南海对于台风伊布都响应的数值计算[J]. 物理学报, 2009(9): 6609-6615.
Cui H, Zhang S W, Wang Q Y. Numerical calculation of the response of the south China sea to typhoon imbudo[J]. Acta Physica Sinica, 2009(9): 6609-6615.
[27]Yang H, Liu Q, Liu Z, et al. A general circulation model study of the dynamics of the upper ocean circulation of the South China Sea[J]. Journal of Geophysical Research-Oceans, 2002, 107(C7): 21-22.
[28]Wang G, Ling Z, Wang C. Influence of tropical cyclones on seasonal ocean circulation in the South China Sea[J]. Journal of Geophysical Research Oceans, 2009, 114(C10): 157-165.
[29]宫响, 史洁, 高会旺. 海洋次表层叶绿素最大值的特征因子及其影响因素[J]. 地球科学进展. 2012, 27(5): 539-548.
Gong X, Shi J, Gao H W. Subsurface chlorophyll maximum in ocean: its characteristics and influencing factors[J]. Advances in Earth Science, 2012, 27(5): 539-548.
[30]Gong X, Shi J, Gao H W, et al. Steady-state solutions for subsurface chlorophyll maximum in stratified water columns with a bell-shape vertical profile of chlorophyll[J]. Biogeosciences Discussions, 2014, 11(6): 9511-9538.
[31]Liu K, Chao S, Shaw P, et al. Monsoon-forced chlorophyll distribution and primary production in the South China Sea: observations and a numerical study[J]. Deep Sea Research Part I: Oceanographic Research Papers, 2002, 49(8): 1387-1412.
[32]Wang G, Su J, Ding Y, et al. Tropical cyclone genesis over the South China Sea[J]. Journal of Marine Systems, 2007, 68(3): 318-326.
[33]Ha Y, Zhong Z, Sun Y, et al. Decadal Change of South China Sea Tropical Cyclone Activity in Mid-1990s and Its Possible Linkage with Intraseasonal Variability[J]. Journal of Geophysical Research: Atmospheres, 2014,119(9): 5331-5344.
[34]Blumberg A F, Mellor G L. A description of a three-dimensional coastal ocean circulation model[J]. Three-Dimensional Coastal ocean Models, 1987: 1-16.
[35]Baretta J W, Ebenhöh W, Ruardij P. The European regional seas ecosystem model, a complex marine ecosystem model[J]. Netherlands Journal of Sea Research, 1995, 33(3): 233-246.
[36]Vichi M, Baretta J W, Baretta-Bekker J G, et al. European regional seas ecosystem Model III: Review of the biogeochemical equations[J]. Documentation Release Version, 2004:1-88.
[37]Martin P J. Simulation of the mixed layer at OWS November and Papa with several models[J]. Journal of Geophysical Research: Oceans (1978-2012), 1985, 90(C1): 903-916.
[38]袁业立. 黄海冷水团环流——I. 冷水团中心部分的热结构和环流特征[J]. 海洋与湖沼, 1979, 10(3): 187-196.
Yuan Y L. The circulation of yellow sea cold water mass——I. Structure of the heat and characteristics of the circulation in the central part of the cold water mass[J]. Oceanologia Et Limnologia Sinica, 1979, 10(3): 187-196.
[39]袁业立, 李惠卿. 黄海冷水团环流结构及生成机制研究——I. 0 阶解及冷水团的环流结构[J]. 中国科学 B 辑, 1993, 23(01): 93-103.
Yuan Y L, Li H Q. Study on structure of the circulation and generation mechanism of the yellow sea cold water mass——I. The zero order solution and structure of the circulation in the cold water mass[J]. Science in China (Series B), 1993, 23(01): 93-103.
[40]张书文. 黄海冷水团夏季叶绿素垂向分布结构的影响机制[J]. 海洋与湖沼. 2003, 34(2): 179-186.
Wen Z S. Effect mechanisms on chlorophyll-a vertical distribution in the summer within Yellow Sea Cold Water Mass[J]. Oceanologia Et Limnologia Sinica, 2003, 34(2): 179-186.
[41]胡好国, 袁业立, 万振文. 海浪混合参数化的渤海、黄海、东海水动力环境数值模拟[J]. 海洋学报(中文版), 2004(4): 19-32.
Hu H G, Yuan Y L, Wan Z W. Study on hydrodynamic environment of the Bohai Sea, the Huanghai Sea and the East China Sea with wave-current coupled numerical model[J]. Acta Oceanol, Sin, 2004(4): 19-32.
[42]Huang C J, Qiao F. Wave-turbulence interaction and its induced mixing in the upper ocean[J]. Journal of Geophysical Research: Oceans (1978-2012), 2010, 115(C4): 1-12.
[43]Liu K, Chao S, Shaw P, et al. Monsoon-forced chlorophyll distribution and primary production in the South China Sea: observations and a numerical study[J]. Deep Sea Research Part I: Oceanographic Research Papers, 2002, 49(8): 1387-1412.
[44]Vichi M, Masina S. Skill assessment of the PELAGOS global ocean biogeochemistry model over the period 1980-2000[J]. Biogeosciences Discussions, 2009, 6(2): 3511-3562.
[45]Fairall C W, Bradley E F, Rogers D P, et al. Bulk parameterization of air-sea fluxes for Tropical Ocean-global atmosphere coupled-ocean atmosphere response experiment[J]. Journal of Geophysical Research: Oceans (1978-2012), 1996, 101(C2): 3747-3764.
[46]陳英傑. 南海初級生產力之估算與 SEATS 測站顆粒態有機質之觀測: 應用於南海物理-生地化耦合模式之驗證與改進[D]. 台湾: 国立台湾大学, 2004.
Chen Y J. Estimation of primary production in the South China Sea and observation of particulate organic matter at SEATS station: application in the validation and improvement of the coupled physical-biogeochemical model of South China Sea[D]. Taiwan: National Taiwan University, 2004.
[47]Wagner R G. Decadal-scale trends in mechanisms controlling meridional sea surface temperature gradients in the tropical Atlantic[J]. Oceanographic Literature Review, 1997, 101: 16683-16694.
[48]Fisher T R. Nutrient limitation of phytoplankton in Chesapeake Bay[J]. Marine Ecology Progress, 1992, 82(1): 51-63.
[49]Tseng C M, Wong G T, Lin I I, et al. A unique seasonal pattern in phytoplankton biomass in low-latitude waters in the South China Sea[J]. Geophysical Research Letters, 2005, 32(8): 487-500.
[50]Liu G, Chai F. Seasonal and interannual variability of primary and export production in the South China Sea: A three-dimensional physical-biogeochemical model study[J]. ICES Journal of Marine Science: Journal du Conseil, 2009, 66(2): 420-431.
[51]Naik H, Naqvi S, Suresh T, et al. Impact of a tropical cyclone on biogeochemistry of the central Arabian Sea[J]. Global Biogeochemical Cycles, 2008, 22(3): 428-451.
[52]Sun L, Yang Y, Xian T, et al. Strong enhancement of chlorophyll a concentration by a weak typhoon[J]. arXiv preprint arXiv: 1004. 2329. 2010,404(6): 39-50.
[53]Bender M A, Ginis I, Kurihara Y. Numerical simulations of tropical cyclone-ocean interaction with a high-resolution coupled model[J]. Journal of Geophysical Research Atmospheres, 1993, 98(D12): 23245-23263.
责任编辑庞旻
Impacts of Typhoon on Ocean Primary Production and Nutrients Transport
PAN Shan-Shan1, SHI Jie1, GAO Hui-Wang1, XU Zhao2, YAN Xiu-Hua2
(1.The Key Laboratory of Marine Environment and Ecology, Ministry of Education, Ocean University of China, Qingdao 266100, China; 2.The Key Laboratory of Global Change and Marine-Atmospheric Chemistry, Third Institute of Oceanography, State Oceanic Administration, Xiamen 361005, China)
Abstract:Typhoons are intense atmospheric events with strong effects on the physical and biological environment in oceans. In this study, a 1-D physical-biological coupled model was used to study the impacts of the typhoon Prapiroon and the supper typhoon Chanchu on the vertical structures of water temperature, nutrients and chlorophyll-a in the northern South China Sea. The model successfully reproduced the climatological seasonal variations of the physical and biological elements by comparing the model results and the observations at the SEATS station. Besides, the sea surface temperature (SST) and chlorophyll-a concentration (Chl-a) compared well with satellite data during the typhoon passages. The model results indicated that the typhoon Parpiroon resulted in cooling in the water above 50 m with the maximum decrease of SST by 2.0℃, and warming in the deep layer from 50 m to about 130 m. The mixed layer depth (MLD) deepened by 30 m. The maximum increase of sea surfaceChl-awas 0.18 mg m-3. The integrated primary production (IPP) yielded by typhoon Prapiroon was 2.3×103mg C·m-3, accounting for ~3% of the annual IPP. Typhoon Chanchu resulted in cooling in the water above 55 m with the maximum decrease of SST by 5.0℃, and warming in the deep layer from 55 m to about 150 m. The MLD deepened by 85 m. The maximum increase of Chl-aat surface was 0.90 mg·m-3. The upward amounts of nutrients pumped by typhoon Chanchu accounted for ~30% of annual value. The IPP yielded by typhoon Chanchu reached up to 12.8×103mg C·m-3, accounting for 18% of the annual IPP. In addition, the effects of typhoons on ocean ecosystem mainly depend on the intensity and translation speed of typhoons, and the pre-conditions of the ocean before typhoon passage such as water stratification and nutrients level should also be considered.
Key words:the Northern South China Sea; typhoon; phytoplankton bloom; primary production; physical-biological coupled model
基金项目:❋ 国家海洋局海洋-大气化学与全球变化重点实验室开放基金项目(GCMAC1403);中国博士后科学基金项目(2014M560575)资助
收稿日期:2015-08-24;
修订日期:2015-12-14
作者简介:潘珊珊(1989-),女,硕士。 ❋❋通讯作者:E-mail:shijie@ouc.edu.cn
中图法分类号:P732.7
文献标志码:A
文章编号:1672-5174(2016)06-120-14
DOI:10.16441/j.cnki.hdxb.20150297
Supported by the Key Laboratory of Global Change and Marine-Atmospheric Chemistry, SOA (GCMAC1403) ; China Postdoctoral Science Foundation (2014M560575)
