不同营养方式下小球藻生长与光合作用的变化*
2016-06-27孙春晓乔洪金王际英张利民
王 超,孙春晓,乔洪金**,丛 超,3,王际英,张利民
(1.青岛大学生命科学学院,山东青岛 266071;2.山东省海洋资源与环境研究院,山东省海洋生态修复重点实验室,山东烟台 264006;3.上海海洋大学水产与生命学院,上海 201306)
不同营养方式下小球藻生长与光合作用的变化*
王超1,孙春晓2,乔洪金2**,丛超2,3,王际英2,张利民2
(1.青岛大学生命科学学院,山东青岛266071;2.山东省海洋资源与环境研究院,山东省海洋生态修复重点实验室,山东烟台264006;3.上海海洋大学水产与生命学院,上海201306)
摘要:【目的】研究小球藻在自养条件下和以乙酸为碳源的异养、混养条件下生长以及光合作用的变化。【方法】以小球藻Chlorella sorokiniana为研究对象,通过测定OD550和光系统Ⅱ(PSⅡ)叶绿素荧光研究其生长情况和光合作用的变化。【结果】小球藻在初始接种浓度为5×106个/mL的条件下,异养和混养的生长速度显著快于自养,到达稳定期仅需1.5 d,而自养生长需要9 d;叶绿素荧光参数自养大于混养,而混养又大于异养,有效光量子产量混养比自养降低21.5%,异养比自养降低98.1%;混养的Rubisco酶基因mRNA表达量最高,分别是异养和自养的3.2倍和1.8倍。【结论】小球藻在混养条件下光系统仅受到微弱抑制,生长速度和最高细胞密度均高于其它营养方式,适合规模化培养。
关键词:小球藻自养异养混养光合作用
0引言
【研究意义】小球藻是一种可进行多种营养方式的单细胞绿藻,其营养方式主要分为3种:一是利用光能和CO2进行光合自养生长;二是在黑暗条件下利用有机碳源和O2进行异养生长,类似于微生物的发酵作用[1-2];三是在光照条件下利用有机碳源进行混养生长,光合作用和发酵作用同时进行[3-4]。由于营养方式多样,生长快,光合效率高,且富含蛋白、脂肪和多糖等物质,小球藻在光合作用机理研究[5-6],抗生素与抗肿瘤等药物的开发[7-8],食品和水产饲料[8-9],高密度规模化培养与生物柴油炼制[10-12],以及污水处理[13-14]等方面都有应用。【前人研究进展】乙酸是一种很多种类的微藻都可以利用的碳源,可显著促进微藻在异养和混养条件下快速生长。早在1960年,Pringsheim等[15]就报道乙酸能引起衣藻的不正常光合作用,并微弱地抑制小球藻的光合活性。接着,Eppley等[16]在衣藻中发现光合作用产生的ATP和还原力被直接用于同化乙酸。此后,许多学者以衣藻作为材料对乙酸的效应进行研究。一方面,乙酸抑制衣藻光反应及其相关过程:减少氧气释放[17],抑制光捕获蛋白的表达[18];另一方面,乙酸降低衣藻暗反应的相关活性:降低碳酸酐酶活性[19],抑制Rubisco酶的表达[20]和光合固碳[17]。【本研究切入点】尽管在20世纪60年代乙酸被成功地应用于研究小球藻的乙醛酸循环[21-23],但是之后有关此方面的研究报道很少。随着小球藻规模化生产的开展,需进一步研究小球藻在乙酸条件下的光合营养机制。【拟解决的关键问题】以小球藻为材料,研究其在添加乙酸的条件下,自养、异养和混养条件下的生长变化,以及光合作用光反应和暗反应的变化,旨在为小球藻的异养和混养规模化生产提供理论依据。
1材料与方法
1.1材料与培养条件
所用小球藻藻种(Chlorella sorokiniana)由中国科学院海洋研究所馈赠。细胞培养在光照恒温摇床中,转速150 r·min-1,温度30℃,由冷荧光灯管提供的光照强度为80 μmol ·m-2·s-1,光暗比12 h∶12 h。自养(AT)培养采用BBM培养基[24],异养(HT)和混养(MT)培养则采用BBM培养基加上30 mmol/L的乙酸钠,初始接种浓度为5×106个/mL,其中异养培养瓶以黑色塑料布包裹避光,每种处理均重复3次。
1.2细胞生长的测定
微藻的生长通过在紫外分光光度计上(Shimadzu UV-1800,日本)测量550 nm处吸光度的变化来反映。同时用血球计数板进行细胞计数,得到细胞个数与OD550的标准曲线,根据OD550值就可以转换出每毫升细胞数。自养培养每天取样一次,异养和混养培养每隔半天取样一次。
1.3光系统Ⅱ(PSⅡ)叶绿素荧光的测定
使用具有调制荧光技术的Imaging-PAM系统(Heinz Walz,德国)测量PSⅡ的原位叶绿素荧光,该系统连接到一台安装有视窗软件的PC获取数据。使用慢速动力学程序自动诱导叶绿素荧光。分别取自养、异养和混养条件下对数生长期中期的藻液,首先暗适应15 min,然后使用饱和脉冲激发藻液,立即测量最小(F0)和最大(Fm)荧光值,计算出PSⅡ的最大光化学量子产量(Fv/Fm)。然后,光化光被打开,藻液开始光合作用,光照样品的最大荧光值(Fm′)被记录。PSⅡ的有效光化学量子产量Y(Ⅱ)、相对电子传递速率rETR,非光化学淬灭NPQ按照下列公式计算:
Y(Ⅱ) = (Fm′-F)/Fm′,
(1)
rETR=Y(Ⅱ)·PAR·0.84·0.5,
(2)
NPQ=(Fm-Fm′)/Fm′。
(3)
其中,PAR是光合有效辐射,0.84是植物的经验性吸光系数,0.5是假设植物吸收的光能被两个光系统均分。
1.4Rubisco酶mRNA表达量的测定
分别取自养、异养和混养条件下对数生长期中期的藻液,离心收集后采用Trizol法提取总RNA,并反转录为cDNA。使用定量PCR,设计引物扩增Rubisco大亚基上的片段(正向引物:5′-CAGCGGTAGCGGCAGAATC-3′;反向引物:5′-CTTCACCAGGAACAGGCTCG-3′)和内参基因18S rRNA上的片段(正向引物:5′-CTGAGAAACGGCTACCACATC-3′;反向引物:5′-CCCCACCCGAAATCCAAC-3′),PCR体系包含:10 μmol·L-1正向和反向引物各0.5 μL、无菌双蒸水10.5 μL和PCR mix (SYBR Green Realtime PCR Master Mix,大连宝生物公司) 12.5 μL,待测样品cDNA 1 μL,每个样品重复3次。反应条件:94℃预变性3 min;94℃变性10 s,60℃退火延伸1 min,35个循环,然后进行55~95℃的熔解曲线分析,荧光波长为490 nm,在icycler real-time quantity PCR仪(BIO-RAD公司,美国)上进行反应。数据处理采用△△Ct相对定量法。
1.5数据处理
采用SPSS 11.0(SPSS Inc.,美国)对数据进行单因素方差分析(one-way ANOVAs),结果用平均数±标准差(M±SD)表示。若存在显著差异,则应用Duncan多重比较法确定组间差异,当P < 0.05时,表示差异显著。
2结果与分析
2.1不同营养方式下小球藻的生长情况
经测定,小球藻藻液OD550值与细胞个数的换算公式为y=2×107x(R2=0.99,y是每毫升细胞个数,x是OD550值)。小球藻在自养条件下经历0.5 d的延滞期后进入对数生长期,9 d左右结束对数生长期,然后进入一个缓慢生长的稳定期;在异养和混养条件下,经历0.5 d的延滞期后迅速进入对数生长期,在1.5 d左右完成对数生长进入稳定期,混养生长基本维持细胞密度不变,而异养生长出现细胞密度缓慢下降的现象(图1)。混养生长最高细胞密度可达(3.0±0.2)×107个/mL,显著高于异养生长的最高细胞密度(2.7±0.4)×107个/mL(P<0.05),混养和异养生长的最高细胞密度又显著高于自养生长的最高细胞密度(2.3±0.2)×107个/mL(P<0.05)。
2.2不同营养方式下小球藻的叶绿素荧光参数
如表1所示,小球藻在自养条件下的各项叶绿素荧光参数均显著高于异养和混养条件(P<0.05),而混养条件的各项参数又显著高于异养条件(P<0.05)。在异养条件下,Fv/Fm、Y(Ⅱ)和rETR值均不到自养和混养条件下的10%,NPQ值分别是自养和混养条件下的21.7%和36.0%。混养条件下Fv/Fm、Y(Ⅱ)、rETR和NPQ分别比自养条件下下降13.2%、21.5%、21.9%和40.0%。
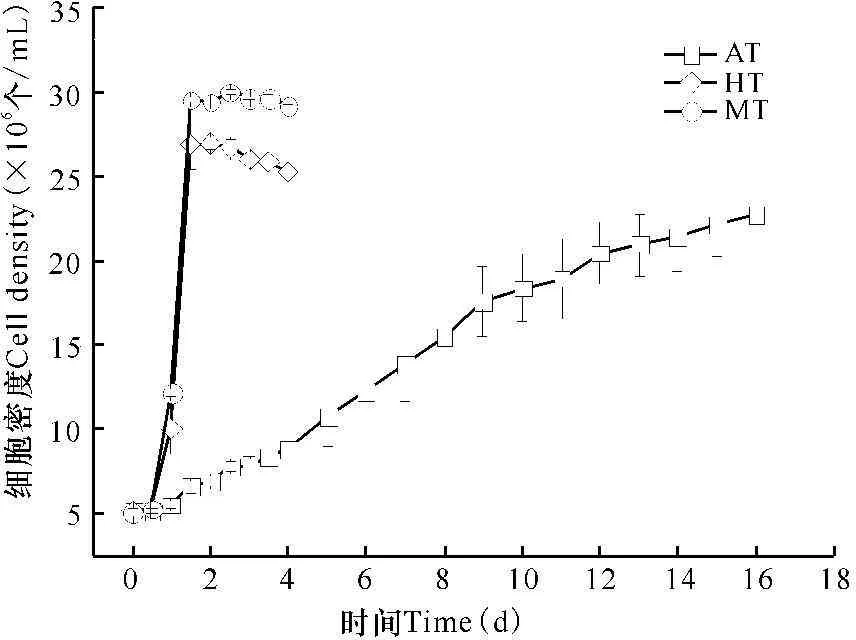
图1不同营养方式下小球藻的生长曲线
Fig.1Growth curves of Chlorella sorokiniana under different trophic modes
2.3不同营养方式下小球藻Rubisco酶的mRNA表达量
如图2所示,以Rubisco酶大亚基基因(rbcL)mRNA的表达量代表Rubisco酶的表达量,小球藻在混养条件下Rubisco酶的表达量最高,分别是异养和自养条件下的3.2倍和1.8倍,而自养条件下的表达量是异养条件的1.8倍,三者之间的差异显著(P<0.05)。
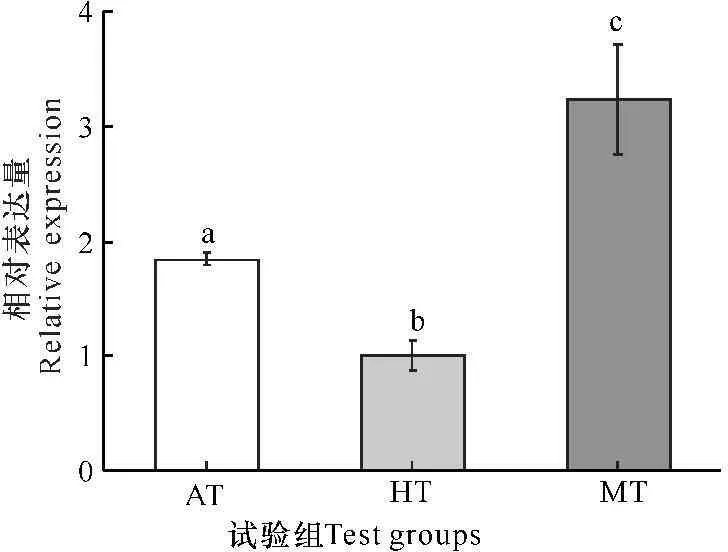
以HT组为参照进行归一化处理,标注不同英文字母的组具有显著性差异(P<0.05)
Transcript level of rbcL in AT and MT is normalized to that in HT.Values with different letters are significantly different (P<0.05)
图2不同营养方式下小球藻rbcL基因mRNA的相对表达量
Fig.2Messenger RNA expression of rbcL gene under different trophic modes in Chlorella sorokiniana
表1不同营养方式下小球藻叶绿素荧光参数的比较
Table 1Comparison of chlorophyll fluorescence parameters under different trophic modes in Chlorella sorokiniana

样品SampleFv/FmY(Ⅱ)rETRNPQAT0.713±0.003a0.538±0.009a18.000±0.265a0.060±0.012aHT0.061±0.010b0.010±0.004b0.250±0.212b0.013±0.004bMT0.619±0.005c0.422±0.008c14.067±0.252c0.036±0.003c
注:同一列中上标英文字母不同的组与同组数据相比差异显著,P<0.05
Note:Values within the same column with different letters are significantly different (P<0.05)
3讨论
小球藻在无光和有光条件下均可以利用乙酸进行生长,并且生长速度显著快于自养生长,对数生长期显著缩短,据报道,混养条件下的细胞生长速率可以大约看作是自养和异养生长速率之和[3,25-26],这与文章的结果基本一致(图1)。许多研究者证明乙酸主要是经过乙醛酸循环被利用[21,23,27],而乙醛酸循环是独立于光合作用的,因而乙酸可以在黑暗条件下被小球藻利用。
小球藻在异养条件下表现出极低的光合效率(表1,图2),表明其光系统已经大部分失去活性,暗反应的关键酶Rubisco酶也仅维持本底表达量,在此条件下小球藻完全依赖发酵作用产生的能量维持生长和分裂,表现出类似于真菌的营养特性。而在混养条件下,其光合效率与自养条件相比也受到一定的抑制(表1),其中代表潜在最大光合作用效率的Fv/Fm以及实际光合作用效率的Y(Ⅱ)均低于自养条件,这与Pringsheim等[15]、Heifetz等[17]、Kroymann等[28]的报道一致。然而,混养条件下的Rubisco酶的mRNA表达量要显著高于自养条件,表明至少在转录水平上Rubisco酶未受到抑制。但是mRNA表达量不能代表最终蛋白表达量,因此需要进一步研究Rubisco酶活性以确定暗反应是否也受到混养条件的抑制。
4结论
小球藻在乙酸为碳源的异养和混养条件下均能极快生长,仅1.5 d就可以到达稳定期,但异养条件下光系统已大部分失去活性,而混养生长光系统仅受到微弱抑制,既保留异养发酵生长的特性,又可以进行自养条件下的光合作用,因此生长速度和最终的生物量均高于其它营养方式,适合规模化放大培养。
参考文献:
[1]GRIFFITHS D A,GRIFFITHS D J.The fine structure of autotrophic and heterotrophic cells of Chlorella vulgaris (Emerson strain)[J].Plant Cell and Physiology,1969,10(1):11-19.
[2]WALTER J,RICKERT M,AACH H.The role of glucose on the enzymes involved in the release of mature spores of Chlorella fusca[J].Physiologia Plantarum,1987,71(2):219-223.
[3]ENDO H,SANSAWA H,NAKAJIMA K.Studies on
Chlorella regularis,heterotrophic fast-growing strain Ⅱ.Mixotrophic growth in relation to light-intensity and acetate concentration[J].Plant and Cell Physiology,1977,18(1):199-205.
[4]吴庆余,匡梅,GRANT N G.小球藻两个品系在自养与异养条件下的生长、能荷与色素差异[J].植物生理学报,1992,18(3): 293-299.
WU Q Y,KUANG M,GRANT N G.Growth,AEC and pigments of Chlorella protothecoides in two strains under the autotrohic and heterotrophic conditions[J].Acta Phytophysiologica Sinica,1992,18(3):293-299.
[5]BASSHAM J A,CALVIN M.The Path of Carbon in
Photosynthesis [M].Berkeley:Interscience Publishers,1960:1-66.
[6]LEY A,MAUZERALL D.Absolute absorption crossse-
ctions for photosystem Ⅱ and the minimum quantum requirement for photosynthesis in Chlorella vulgaris[J].BBA-Bioenergetics,1982,680(1):95-106.
[7]江红霞,郑怡.微藻的药用、保健价值及研究开发现状[J].亚热带植物科学,2003,32(1):68-72.
JIANG H X,ZHENG Y.A review of pharmaceutical and health care value of microalgae and their current status of research and development [J].Subtropical Plant Science,2003,32(1):68-72.
[8]BOROWITZKA M A.Microalgae source of pharmaceuticals and other biologically active compounds[J].Journal of Applied Phycology,1995,7(1):3-15.
[9]PULZ O,GROSS W.Valuable products from biotechnology of microalgae[J].Applied Microbiology and Biotechnology,2004,65(6):635-648.
[10]贺立燕,韩笑天,俞志明.基于熵权的产生物柴油微藻开发潜力评价[J].海洋与湖沼,2015,46(2):305-310.
HE L Y,HAN X T,YU Z M.Evaluation of microalgae potential for biodiesel production in entropy weight theory[J].Oceanologia et Limnologia Sinica,2015,46(2):305-310.
[11]RODOLFI L,ZITTELLI G C,BASSI N,et al.Microalgae for oil:Strain selection,induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor[J].Biotechnology and Bioengineering,2009,102(1):100-112.
[12]尹继龙,唐小红,郑洪立,等.不同光质对小球藻光自养培养积累油脂的影响[J].生物加工过程,2014,12(05):62-68.DOI:10.3969/j.issn.1672-3678.2014.05.011.
YIN J L,TANG X H,ZHENG H L,et al.Effect of light wavelengths on lipid accumulation of Chlorella vulgaris in photoautotrophic culture[J].Chinese Journal of Bioprocess Engineering,2014,12(05):62-68.DOI:10.3969/j.issn.1672-3678.2014.05.011.
[13]胡沅胜,刘斌,郝晓地,等.微藻处理污水中的絮凝分离/采收研究现状与展望[J].环境科学学报,2015,35(1):12-29.
HU Y S,LIU B,HAO X D,et al.Current status and outlook of microalgae flocculation in wastewater treatment[J].Acta Scientiae Circumstantiae,2015,35(1):12-29.
[14]ASLAN S,KAPDAN I K.Batch kinetics of nitrogen
and phosphorus removal from synthetic wastewater by algae[J].Ecological Engineering,2006,28(1):64-70.
[15]PRINGSHEIM E G,WIESSENER W.Photo-assimilation of acetate by green organisms [J].Nature,1960,188:919-921.
[16]EPPLEY R W,MACIAS F M.Role of the alga Chlam-
ydomonas mundana in anaerobic waste stabilization lagoons [J].Limnology and Oceanography,1963,8(4):411-416.
[17]HEIFETZ P B,FORSTER B,OSMOND C B,et al.
Effects of acetate on facultative autotrophy in Chlamydomonas reinhardtii assessed by photosynthetic measurements and stable isotope analyses[J].Plant Physiology,2000,122(4):1439-1445.
[18]KINDLE K L.Expression of a gene for a light-harvesting chlorophyll-a-binding chlorophyll-b-binding protein in Chlamydomonas reinhardtii:Effect of light and acetate[J].Plant Molecular Biology,1987,9(6):547-563.
[19]FETT J P,COLEMAN J R.Regulation of periplasmic carbonic-anhydrase expression in Chlamydomonas reinhardtii by acetate and pH [J].Plant Physiology,1994,106(1):103-108.
[20]GOLDSCHMIDT-CLERMONT M.The two genes for the small subunit of RuBP Carboxylase/oxygenase are closely linked in Chlamydomonas reinhardtii[J].Plant Molecular Biology,1986,6(1):13-21.
[21]SYRETT P J,MERRETT M J,BOCKS S M.Enzymes of glyoxylate cycle in Chlorella vulgaris [J].Journal of Experimental Botany,1963,14(2):249-264.
[22]SYRETT P J.Kinetics of isocitrate lyase formation in Chlorella-Evidence for promotion of enzyme synthesis by photophosphorylation[J].Journal of Experimental Botany,1966,17(53):641-654.
[23]GOULDING K,MERRETT M.The role of glycollic
acid in the photoassimilation of acetate by Chlorella pyrenoidosa [J].Journal of Experimental Botany,1967,18(4):620-630.
[24]BISCHOFF H W,BOLD H C.Phycological Studies
Ⅳ.Some Soil Algae from Enchanted Rock and Related Algal Species[M].Austin:University of Texas Publication,1963.
[25]OGAWA T,AIBA S.Bioenergetic analysis of mixotrophic growth in Chlorella vulgaris and Scenedesmus acutus [J].Biotechnology and Bioengineering,1981,23 (5):1121-1132.
[26]MARQUEZ F J,SASAKI K,KAKIZONO T,et al.
Growth-characteristics of Spirulina platensis in mixotrophic and heterotrophic conditions [J].Journal of Fermentation and Bioengineering,1993,76(5):408-410.
[27]LALIBERTÉG,NOÜE J.Auto-,hetero- and mixotrophic growth of Chlamydomonas humicola (Chlorophyceae) on acetate [J].Journal of Phycology,1993,29(5):612-620.
[28]KROYMANN J,SCHNEIDER W,ZETSCHE K.Opposite regulation of the copy number and the expression of plastid and mitochondrial genes by light and acetate in the green flagellate Chlorogonium[J].Plant Physiology,1995,108(4):1641-1646.
(责任编辑:米慧芝)
Variation of Growth and Photosynthesis in Chlorella sorokiniana Under Different Trophic Modes
WANG Chao1,SUN Chunxiao2,QIAO Hongjin2,CONG Chao2,3,WANG Jiying2,ZHANG Limin2
(1.College of Life Science,Qingdao University,Qingdao,Shandong,266071,China;2.Key Laboratory of Marine Ecological Restoration,Shandong Marine Resource and Environment Research Institute,Yantai,Shandong,264006,China;3.College of Fisheries and Life Science,Shanghai Ocean University,Shanghai,201306,China)
Abstract:【Objective】The variation of growth and photosynthesis of microalgae was studied and compared under autotrophic condition and hetero- and mixotrophic conditions with acetate as carbon source.【Methods】With Chlorella sorokiniana as material,the changes of growth and photosynthesis were reflected by determining the OD550and the chlorophyll fluorescence from photosystem Ⅱ(PS Ⅱ).【Results】The growth rates of hetero- and mixotrophic conditions were significantly higher than that of autotrophic condition.It was only 1.5 d for cells reaching stationary phase under hetero- and mixotrophic conditions,but 9 d for cells under autotrophic condition.The parameters of chlorophyll fluorescence under autotrophic condition were significantly higher than that under hetero- and mixotrophic conditions,and that under mixotrophic condition were significantly higher than that under heterotrophic condition.The effective PS Ⅱ quantum yield under mixo- and heterotrophic condition was reduced 21.5% and 98.1%,respectively,compared with that of autotrophic condition.The mRNA expression of Rubisco gene under mixotrophic condition were highest,and 3.2-fold and 1.8-fold of that under hetero- and autotrophic conditions,respectively.【Conclusion】The photosystems of Chlorella under mixotrophic condition only slightly inhibited.In addition,the growth rate and maximal cell density under mixotrophic condition were higher than other conditions.Therefore,mixotrophic growth mode is suitable for large-scale cultivation.
Key words:Chlorella sorokiniana,autotrophic,heterotrophic,mixotrophic,photosynthesis
收稿日期:2016-02-21
作者简介:王超(1982-),女,讲师,主要从事藻类生物技术研究。
中图分类号:Q945.1
文献标识码:A
文章编号:1005-9164(2016)02-0115-05
修回日期:2016-04-05
*山东省优秀中青年科学家科研奖励基金项目(BS2013HZ018),国家自然科学基金青年基金项目(31201973),海洋生物产业——水生动物营养与饲料研发创新示范平台项目(201303003)和山东省科技发展计划项目(2014GHY115006)资助。
**通讯作者:乔洪金(1983-),男,副研究员,硕士生导师,主要从事藻类生物技术研究,E-mail:hongjinqiao@hotmail.com。
广西科学Guangxi Sciences 2016,23(2):115~119
网络优先数字出版时间:2016-05-12
网络优先数字出版地址:http://www.cnki.net/kcms/detail/45.1206.G3.20160512.0904.002.html
