龙南钾矿区常见蕨类植物可培养内生真菌的多样性
2016-06-23甘会云杜勇涛樊有赋
钱 旭, 甘会云, 杜勇涛, 张 鲜, 陈 晔, 樊有赋
( 九江学院 药学与生命科学学院, 江西 九江 332000 )
龙南钾矿区常见蕨类植物可培养内生真菌的多样性
钱旭, 甘会云, 杜勇涛, 张鲜, 陈晔*, 樊有赋
( 九江学院 药学与生命科学学院, 江西 九江 332000 )
摘要:植物内生真菌作为一类特殊的微生物资源,与宿主在长期的生态系统演化过程中形成了互惠共生关系,通过多样化途径来增强植物体的营养生理和抗性机能,对宿主植物产生多种有益生物学作用,在植物演替过程中具有重要的生态学意义。这种特殊微生物资源近年来倍受关注,而利用植物-真菌共生体强化植物在矿区逆境中生长,提高矿区生物修复效率是一个新的研究热点。为探明钾矿区不同蕨类植物内生真菌的物种多样性、群落组成以及生态分布规律,该研究以芒萁、狗脊、禾秆蹄盖蕨、海金沙、华中铁角蕨、井栏边草和乌蕨等植物为材料,采用组织分离、形态学鉴定等方法,对其内生真菌多样性进行分析。结果表明:从7种植物中共分离获得377株内生真菌,总分离率在2.50%~4.52%之间。经鉴定377个菌株隶属于链格孢属、曲霉属、枝孢菌属和轮枝孢属等25个分类单元,其中链格孢属、曲霉属、枝孢菌属和轮枝孢属等在所有被调查蕨类植物中都有分布,为优势属,共计为185株,占总株数的49.07%,但它们在每种植物的分布存在明显差异;7种蕨类植物内生真菌总定植率为叶高于根状茎 (P<0.05),多样性指数在0.502~0.867之间,但每种植物及其不同组织部位的内生真菌定植率和多样性指数存在一定的差异;从相似性分析来看,同一个钾矿区不同蕨类植物内生真菌菌群之间的相似性程度较低,相似性系数在0.189~0.587之间。该研究结果不仅丰富了植物内生真菌种质资源,而且为进一步开展植物内生真菌强化宿主植物在钾矿区生长适应机制的研究奠定了基础。
关键词:钾矿区, 蕨类植物内生真菌, 优势菌群, 多样性指数, 相似性系数
钾是植物生长发育必需元素之一,土壤中的钾可分成矿物钾(占90%左右)、缓效钾(2%~8%)和速效钾(0.1%~0.2%)。矿物钾是植物难以利用的钾,如长石和白云母中的钾。各种形态钾的含量及其相对活性,决定了钾对植物的有效性。如何将矿物钾中营养元素溶出,形成有利于植物吸收的活性物质倍受关注(蒋梅茵,1982;徐晓燕和马毅杰,2001; 潘大伟等,2005)。植物内生真菌作为一类特殊的微生物资源,具有分布广、种类多等特点。自20世纪80年代以来,国内外学者对温带、亚热带、热带、盐碱地等不同生境植物内生真菌多样性进行了调查研究(Tejesvi,2011;Jyoti et al,2012;Kristin et al,2012;刘爱荣等,2010)。研究表明,植物内含有较丰富的内生真菌资源,受组织、环境等因素的影响体现出一定的组织、宿主差异性(郭良栋,2000)。植物内生真菌与宿主在长期的生态系统演化过程中形成了互惠共生关系,对宿主植物产生多种有益生物学作用(郭良栋,2000;Atsatt & Whiteside,2014)。应用植物—微生物共生体强化植物在矿区逆境中生长已成为新的研究热点(牛之欣等,2009)。
目前,内生真菌的宿主植物研究主要集中在种子植物,而有关蕨类等孢子植物内生真菌多样性及其生态功能活性研究的报道却相对很少(张君诚等,2010;邹文欣等,2001)。蕨类植物是一类较原始的高等植物,也是较早的陆生植物。现存的蕨类植物约有12 000种,世界各地均有分布,具有丰富的物种多样性。蕨类植物具有抗逆境能力强、生长迅速、繁殖能力强等特点,是较理想的植物修复资源(褚磊等,2007)。具有内生真菌定植的蕨类植物有Pteridiumaquilinu,Equisetumhyemale,Botrychiumternatum,Huperziaserrata,Huperziaserratavar.longipetiolata,Huperziaappressa,Huperziacrispata,Phlegmariuruscryptomerianus,Schizaeapusilla,Nodulisporiumsp.,Nephrolepisexaltata等(Gabel et al,1996;Petrini et al,1992;Sati et al,2006,2009;石玮等,2005;鞠錾等,2009;樊有赋等,2008;巫婷玉等,2012;詹寿发等,2007,2012;Riyaz-UI-Hassan et al,2013; Thangavelu & Kandasamy,2012),但对蕨类植物内生真菌的种类、形态结构、生态分布等尚缺乏全面系统地研究。
相对于丰富的蕨类植物资源,还有大量蕨类植物的内生真菌亟待研究,特别是矿区环境蕨类植物内生真菌的研究。本研究对采自江西省龙南县钾矿区7种常见蕨类植物进行内生真菌的分离、纯化和形态学鉴定,以探索钾矿区蕨类植物内生真菌的物种多样性、群落组成以及生态分布规律,为进一步探究植物内生真菌对宿主植物在矿区适应影响机制提供了理论依据。
1材料与方法
1.1 样地概况
龙南县安基山钾矿区位于江西省最南端龙南县程龙镇(114°23′~114°59′ E、24°29′~25°1′ N),占地面积15 000 m2,年产钾矿0.9 万t,开采多年,矿区内已堆积大量废弃裸露矿渣(矿渣由钾长石、白云母、黑云母等矿物组成,按照NY/T 889-2004标准测定矿渣速效钾为13.714 mg/mL,占全钾0.015%),生长在矿渣上的植物只有少数蕨类等草本植物,无木本植物生长。
1.2 样品采集处理
2013年7月上旬于江西省龙南县程龙镇安基山钾矿区采集芒萁(Dicranopterislinearis)、狗脊(Woodwardiajaponica)、禾秆蹄盖蕨(Athyriumyokoscense)、乌蕨(Stenolomachusanum)、海金沙(Lygodiumjaponicum)、华中铁角蕨(Aspleniumsarelii)、井栏边草(Pterismultifida)等7种常见蕨类植物样本。采集方法:在随机20 个10 m2样地内采集健康植株(每样地间距200 m),7 种植物各采集20 株健康植株。去除枯叶,地上叶部剪成10 cm带叶柄段片,立即放入无菌塑料袋中,地下根状茎连同土壤一起放入无菌塑料袋中,分别贴上标签放入低温采集箱内带回实验室,4 ℃冰箱保藏,叶在2 d内处理完毕,根状茎在7 d内处理完成。
1.3 内生真菌分离纯化
1.3.1 培养基改良的马铃薯葡萄糖琼脂(PDA,2%)培养基为分离、纯化培养基,配比:马铃薯200 g,葡萄糖20 g,琼脂20 g,10%蕨叶(蕨叶为每种植物自身叶片,如分离芒萁内生真菌采用芒萁叶)水提液100 mL,加蒸馏水定容至1 000 mL,自然pH(使用前在培养基中加80 U/mL庆大霉素1 mL)。
促孢培养基:KH2PO41.0 g,KNO31.0 g,MgSO4·7H2O 0.5 g,KCl 0.5 g,淀粉0.2 g,葡萄糖0.2 g,蔗糖0.2 g,琼脂20 g,蒸馏水1 000 mL,自然pH。 1.3.2 分离纯化按常规无菌操作方法(樊有赋等,2008;巫婷玉等, 2012;詹寿发等,2012),叶部从每种植物样品中随机选取20段带有叶片的小段片,自来水冲洗干净;按下列程序进行表面消毒:体积分数75%乙醇漂洗2~3 min、无菌水冲洗3次,25% NaClO溶液漂洗5 min、无菌水冲洗3~4次,再用75%乙醇冲洗1 min、无菌水冲洗5次;将样品置于无菌滤纸上吸干水分,剪成0.5 cm × 0.5 cm的片段,从每种植物样品中随机选取80个叶柄和80个叶片接到改良的PDA平板上,置于28 ℃恒温培养箱中黑暗培养3~10 d,每天观察,若发现组织块周围有菌落长出,则将菌转接入斜面,经纯化后得到内生真菌菌株。根状茎在用25%NaClO溶液漂洗5 min、无菌水冲洗3~4次后,用0.1%升汞溶液浸泡5~8 s,无菌水冲洗4~5次,再用75%乙醇冲洗1 min、无菌水冲洗5次,其它步骤同地上部分。分离纯化获得的菌株保存于九江学院药学与生命科学学院真菌研究室(JJTU)。
1.4 内生真菌的鉴定
依据菌落形态、产孢方式、孢子形态特征和产孢结构对菌株进行鉴定。将不产孢菌株在促孢培养基上进行促孢培养,并检查其产孢情况,产孢后按上述方法进行鉴定。经促孢培养后仍不产孢的菌株则根据菌落表面特征、菌丝颜色、菌丝生长速率等分为无孢类群的不同组。真菌属的鉴定主要依据Barnett & Hunter(1998);Keith et al(2011)等相关文献。
1.5 数据统计分析
定植率(Colonization rates,CR)是指样本中受内生真菌侵染的组织块数占样本全部组织块数的百分数;分离率(Isolation rate,IR):分离到的某一指定类型内生真菌的菌株数占样品组织块总数比值;分离频率(Isolation frequency,IF):某一指定类型内生真菌的菌株数占获得总内生真菌菌株数比值。
采用Shannon指数(H)进行内生真菌种群的多样性水平分析。H=-∑[ni/Nln(ni/N)]。式中,ni为第i个内生真菌菌株数目,N为全部内生真菌菌株数。
采用Jaccard相似性指数(Cj)对不同植物间内生真菌组成的相似程度进行比较和分析。Cj=c/(a+b-c)。式中,a是一种植物内生真菌的种类数,b是另一种植物内生真菌的种类数,c是两种植物共同具有的内生真菌种类数,比较两种植物之间内生真菌种类组成的相似程度。
2结果与分析
2.1 内生真菌分离率和定植率
从龙南钾矿区7种常见蕨类植物的叶片、叶柄和根状茎的1 680个组织块中共分离获得377个菌株。从表1可见,7种蕨类植物内生真菌的总分离率在2.50%~4.52%之间,其中乌蕨的内生真菌总分离率最高(4.52%),其它6种蕨类植物总分离率依次为禾秆蹄盖蕨(3.63%)>海金沙(3.27%)>华中铁角蕨(3.21%)>狗脊(2.74%)>井栏边草(2.56%)>芒萁(2.50%)。在7种蕨类植物中乌蕨的内生真菌总定植率最高(22.92%),其余依次为禾秆蹄盖蕨(17.08%)>华中铁角蕨(16.25%)>海金沙(15.83%)>狗脊(15.42%)>井栏边草(15.00%)>芒萁(14.17%)。从不同组织部位来看,7种蕨类植物内生真菌总分离率和定植率的趋势为叶高于根状茎。这说明蕨类植物内生真菌存在最主要部位是叶,而不是茎。

表 1 钾矿区7种蕨类植物内生真菌分离率和定植率
注: *为占总株数的百分数,P<0.05。
Note: * indicates the percentage of total strains number,P<0.05.
每种植物不同部位的内生真菌分离率和定植率存在差异,其中狗脊、禾秆蹄盖蕨、海金沙、乌蕨、芒萁等5种植物内生真菌的定植率和分离率依次为叶柄>叶片>根状茎,华中铁角蕨、井栏边草内生真菌的定植率和分离率依次为叶片>根状茎>叶柄。
2.2 内生真菌类群组成
从钾矿区7种常见蕨类植物中共分离获得内生真菌377个菌株,经鉴定属于链格孢属(Alternaria)、曲霉属(Aspergillus)、枝孢菌属(Cladosporium)和轮枝孢属(Verticillium)等25个分类单元(表2)。根据分离频率判断曲霉属、枝孢菌属、轮枝孢属、链格孢属和Exophiala等属为7种植物内生真菌的优势属,其分离频率分别为14.58%、13.26%、7.69%、7.16%和6.37%,5个属共获得菌株为185株,占总株数的49.07%。这5个分类单元从每种植物中分离获得的菌株数存在明显差异,链格孢属在芒萁中最多(6株),曲霉属在狗脊中最多(12株),枝孢菌属在乌蕨中(16株),Exophiala在禾秆蹄盖蕨中最多(6株),轮枝孢属在井栏边草中较多(7株)。其它20个分类单元在7种植物中分类获得菌株数也存在一些差异,其中Penicillium除芒萁外均可从其它6种植物中分离获得,并在井栏边草中最多(7株),Phomachrysanthemicola,Sclerococcum和Scytalidiym等3个分类单元获得的菌株数最少,主要分布在禾秆蹄盖蕨和乌蕨中。
2.3 不同植物的内生真菌分布
表1研究结果表明,同一矿区不同植物中内生真菌存在明显差异,其中从乌蕨植物中获得76个菌株,占总株数的20.16%。其中禾秆蹄盖蕨占15.92%、海金沙占14.59%、华中铁角蕨占14.32%、狗脊占12.20%、井栏边草占11.41%、芒萁占11.14%。
表2结果表明,每种植物内生真菌优势属不同。其中,狗脊的内生真菌优势属为Aspergillus、Penicillium、Cladosporium和Monilia,共计25个菌株,占其总株数的54.35%;禾秆蹄盖蕨的内生真菌优势属为Cladosporium、Aspergillus、Exophiala和Fusarium,共计33个菌株,占其总株数的54.10%;乌蕨的内生真菌优势属为Cladosporium、Monilia、Fusarium、Oidiodendron、Aspergillus和Alternaria,共计46个菌株,占其总株数的60.53%;海金沙内生真菌的优势属为Aspergillus、Cladosporium、Phomopsis和Penicillium,共计21个菌株,占其总株数的38.18%;华中铁角蕨内生真菌优势属为Aspergillus、Verticillium、Alternaria、Exophiala和Phomopsis,共计29个菌株,占其总株数的53.70%; 井栏边草的内生真菌优势属为Penicilli-um、Verticillium、Aspergillus和Cladosporium,共计25个菌株,占其总株数的58.14%;芒萁内生真菌的优势属为Alternaria、Cephalosporium、Cladosporium、Verticillium和Aspergillus,共计29个菌株,占其总株数的69.05%。
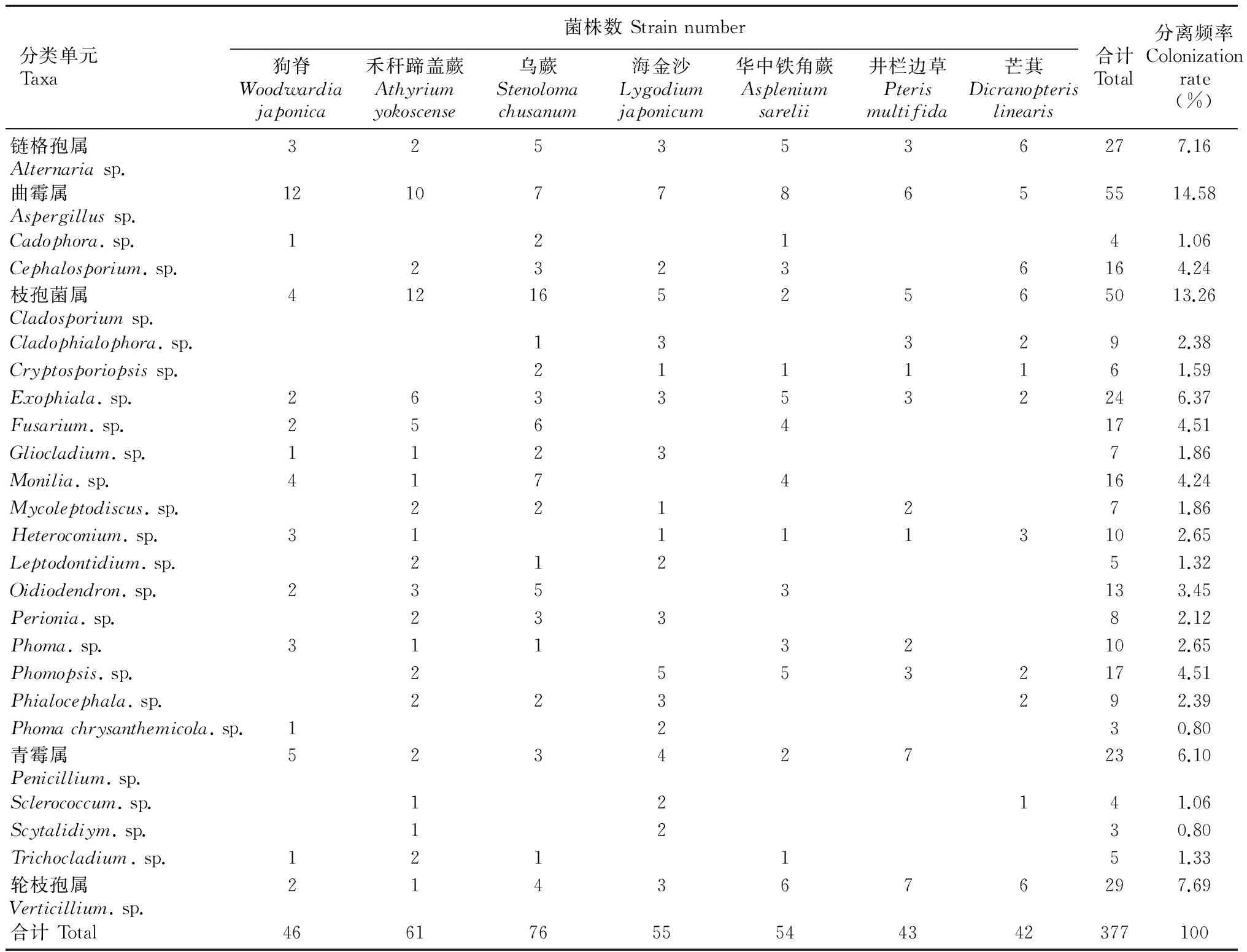
表 2 钾矿区7种蕨类植物内生真菌种群组成
2.4 蕨类植物的内生真菌多样性与相似性
由表3可知,每种蕨类植物内生真菌的多样性指数不同,从高到低的顺序为乌蕨(0.867)>禾秆蹄盖蕨(0.728)>海金沙(0.692)>华中铁角蕨(0.650)>狗脊(0.553)>井栏边草(0.553)> 芒萁(0.553),其中乌蕨内生真菌多样性最为丰富,多样性指数为0.867,芒萁内生真菌多样性最低,多样性指数为0.5020。从组织部位来看,内生真菌的多样性指数顺序为叶片(1.69)>叶柄(1.374)>根状茎(1.105);每种植物不同组织部位的内生真菌多样性指数存在差异,其中狗脊、禾秆蹄盖蕨、海金沙和乌蕨等4种植物的多样性指数依次为叶柄>叶片>根状茎, 而华中铁角蕨和井栏边草的内生真菌多样性指数依次为叶片>根状茎>叶柄,芒萁不同组织部位内生真菌的多样性指数为叶柄=叶片>根状茎。

表 3 蕨类植物内生真菌的多样性指数
对钾矿区7种常见蕨类植物内生真菌相似性分析表明, 不同植物之间内生真菌种类组成的相似性
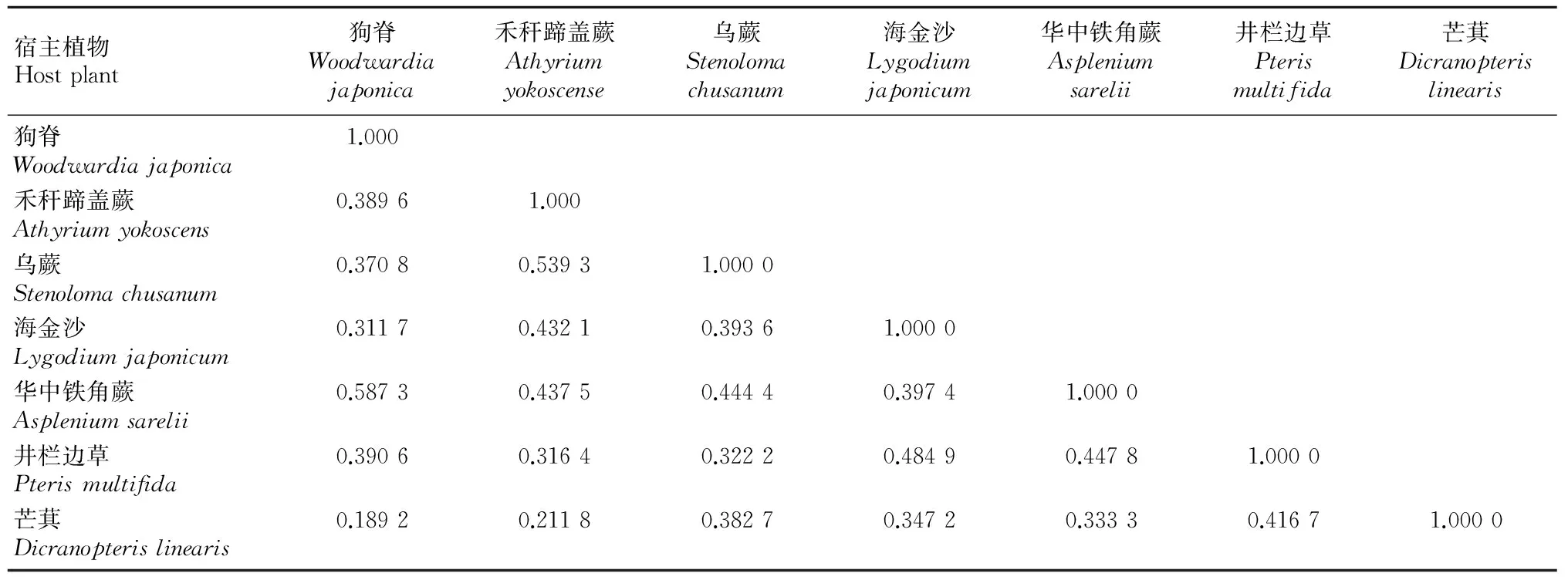
表 4 7种钾矿区常见蕨类植物内生真菌相似性系数
系数在0.189 2~0.587 3之间(表4),说明同一个钾矿区不同蕨类植物内生真菌菌群之间的相似性程度较低。其中,狗脊与华中铁角蕨的内生真菌为中等相似,相似性系数为0.5873;而狗脊与芒萁的内生真菌为极不相似,相似性系数为0.1892。芒萁的内生真菌与其它6种植物内生真菌相似性均较低。
3讨论
本研究从江西龙南钾矿区7种常见蕨类植物的1 680个组织块中共分离获得377个菌株,经鉴定属于Alternaria,Aspergillus,Cladosporium和Verticillium等25个分类单元。根据分离频率判断Aspergillus,Cladosporium,Verticillium,Alternaria和Exophiala等为7种蕨类植物内生真菌的优势属。
从分离率、定植率来看,7种蕨类植物内生真菌的总分离率在2.50%~4.52%之间,总定植率为14.17%~22.92%,说明在钾矿区蕨类植物中有一定数量的植物内生真菌定植。与非钾矿区其它蕨类植物内生真菌的分离率相比要低,如庐山石韦、狗脊、槲蕨等植物分离率在4.75%~5.67%(詹寿发等,2007,2012;巫婷玉等,2012;樊有赋等,2008);与铅锌矿废弃矿区相比钾矿区蕨类植物内生真菌的分离率偏低,如李东伟等(2012)报道的铅锌矿废弃矿区的硬毛南芥(Arabishirsuta)、鱼骨松(Acaciadecurrens)、白檀(Symplocospaniculata)、毛萼香茶菜(Rabbosiaeriocalyx)、蚤辍(Arenariaserpyllifolia)和倒挂刺(Rosalongicuspis)等种子植物内生真菌的分离率在0.42~0.93之间。
从不同组织部位来看,7种蕨类植物叶的内生真菌总分离率和定植率都明显高于根状茎的内生真菌分离率和定植率(P<0.05)(表1),说明蕨类植物内生真菌存在最主要部位是叶。可能与蕨类植物各部位组织结构有关,所研究7种蕨类植物为真蕨类,并且真蕨类植物叶在形态、结构上较发达,叶分为叶片和叶柄,叶片的内部结构为表皮、叶肉组织和叶脉,叶柄的内部结构为表皮、皮层和中柱,它们表皮均有气孔分布,内生真菌可能主要通过自然孔口(气孔)和叶表皮细胞间隙侵入植物体内,蕨类植物茎为根状茎,其内部结构为表皮、皮层和中柱,表皮无气孔,蕨类植物根状茎的内生真菌是通过茎表皮细胞间穿过皮层机械组织细胞间隙侵入到根状茎的皮层薄壁细胞(詹寿发等,2012);也可能与矿区土壤环境有关,因为矿区土壤贫瘠,有些真菌难以在这种环境中生存,故侵入蕨类植物根状茎内生真菌数量较少,而地上部分叶可以接受空气中大量真菌孢子,孢子通过不同途径侵入叶内定植。具体影响内生真菌在蕨类植物不同组织部位分布差异的因素需要进一步探究。
龙南钾矿区7种常见蕨类植物内生真菌多样性指数在0.502~0.867之间,可见钾矿区蕨类植物内生真菌较为丰富。从相似性分析结果来看,不同植物之间内生真菌种类组成的相似性系数在0.189 2~0.587 3之间,除了狗脊与华中铁角蕨、禾秆蹄盖蕨与乌蕨的内生真菌相似性为中等相似外,其它植物之间内生真菌的相似性都比较低,说明同一个钾矿区不同蕨类植物内生真菌菌群之间的相似性程度较低,这是否表明钾矿区蕨类植物内生真菌的宿主具有一定专一性还有待进一步研究。
致谢感谢中国科学院微生物研究所郭良栋研究员给予的帮助和指导。
参考文献:
ATSATT PR,WHITESIDE MD, 2014. Novel symbiotic protoplasts formed by endophytic fungi explain their hidden existence,lifestyle switching,and diversity within the plant kingdom [J]. PLoS One,9(4): e95266.doi:10.1 371/journal.pone. 095 266.
BARNETT HL,HUNTER BB, 1998. Illustrated Genera of Imperfect Fungi (4 Edition) [M]. St. Paul: APS Press: 1-218.BHAGAT J,KAUR A,SHARMA M,et al, 2012. Molecular and functional characterization of endophytic fungi from traditional medicinal plants [J] . World Microbiol Biotechnol,28:963-971.
CHU L,YU XL,LI Y,et al, 2007. Research advances on the heavy metal tolerance of pteridophyte in the mining wasterland [J] . J Anhui Norm Univ,30(5):587-592. [褚磊,于小丽,李影,等, 2007. 矿业废弃地中耐重金属蕨类植物的研究进展 [J]. 安徽师范大学学报,30(5):587-592.]
FAN YF,GAN JL,CHEN Y,et al, 2008. Prelininary study on the endophytic fungi in a medicinal Fera-Cibotiumbaromtz[J] . J Anhui Agri Sci,36(9):3 737,3 375. [樊有赋,甘金莲,陈晔,等, 2008. 药用蕨类植物狗脊内生真菌的初步研究 [J] . 安徽农业科学, 36(9):3 737,3 375.]
GABEL A,STUDT R,METZ S, 1996. Effect ofCryptomycinapteridisonPteridiumaquilinum[J]. Mycologia,88:635-641.
GUO LD, 2000. Advances of researches on endophytic fungi [J]. Mycosystema,20(1):148-152. [郭良栋, 2000. 内生真菌研究进展 [J]. 菌物系统,20(1):148-152.]
JIANG MY, 1982. Fix and release of potassium of soil potassium minerals [J]. Chin J Soil Sci,13(3):44-49;13(4):44-45. [蒋梅茵, 1982. 土壤含钾矿物中钾的固定与释放 [J]. 土壤通报,13(3):44-49;13(4):44-45.]
JU Z,WANG J,PAN SL, 2009. Isolation and preliminary identification of the endophytic fungi which produce Hupzine A from four species in Hupziaceae and determination of Huperzine A by HPLC [J]. Fudan Univ J Med Sci,36(4):445-449. [鞠錾, 王峻, 潘胜利 , 2009. 4种石杉科植物中产生石杉碱甲的内生真菌的分离及初步鉴定和HPLC测定石杉碱甲的含量 [J]. 复旦学报·医学版,36(4):445-449.]
KIRK PM,CANNON PF,MINTER DW,et al, 2008. Dictionary of The Fungi (10th) [M]. CABI Europe-UK: 1-782.
LI DW,XU HM,MEI T,et al, 2012. Diversity of endophytic fungi from six dominant plant species in a Pb-Zn mine wasteland in China [J]. Acta Ecol Sin,32(7):2 288-2 293. [李东伟,徐红梅,梅涛,等, 2012. 云南会泽铅锌矿废弃矿渣堆常见植物内生真菌多样性 [J]. 生态学报,32(7):2 288-2 293.]LIU AR,ZHANG Y,NIU LH,et al, 2010. Analysis of diversity of endophytic fungi in two plants in Hainan Province(Rhizophorastylosaandndeliacandel) [J] . Guihaia,30(5):657-660. [刘爱荣,张洋,牛丽红,等, 2010. 海南两种红树林植物内生真菌的多样性分析 [J] . 广西植物,30(5):657-660.]
MILLER KI,QING C,MAN-YUEN SZE D,et al, 2012. Culturable endophytes of medicinal plants and the genetic basis for their bioactivity [J]. Microb Ecol,64:431-449.
NIU ZX,SUN LN,SUN TH,2009. Plant-microorganism combined remediation of heavy metals-contaminated soils: Its research progress [J]. Chin J Ecol,28(11):2 366-2 373. [牛之欣,孙丽娜,孙铁珩, 2009. 重金属污染土壤的植物-微生物联合修复研究进展 [J]. 生态学杂志,28(11):2 366-2 373.]
PAN DW,LING CH,DU LY, 2005. Progress in research on potassium release from K-beared minerals in soil [J]. Chin J Soil Sci,36(2):253-258. [潘大伟,梁成华,杜立宇, 2005. 土壤含钾矿物的释钾研究进展 [J]. 土壤通报,36(2):253-258.]
PETRINI O,FISHER PJ,PETRINI LE, 1992. Fungal endophytes of bracken(Pteridiumaquilinun) with some reflections on their use in biological control [J]. Sydowia, 44:282-293.
RIYAZ-UI-HASSAN,SYED, STROBEL G,et al, 2013. An endophyticNodulisporiumsp. from Central America producing volatile organic compounds with both biological and fuel potential [J]. Microbiol Biotechnol,23(1):29-35.
SATI SC,BELWAL M, 2005. Aquatic Hyphomycetes as root endophytes of Riparian plant roots [J]. Mycologia,97(1):45-49.
SATI SC,PARGAEIN N,BELWA M, 2009. Diversity of aquatic hyphomycetes as root endophytes on pteridophytic plants in Kumaun Himalaya [J]. J Am Sci, 5(4):179-182.
SATI SC,PARGAEIN N,BELWAL M, 2006. Three species of aquatic hyphomycetes as new root endophytes of temperate forest plants [J]. Nat Acad Sci Ind, 29(2):9-10.
SEIFERT K,MORGAN-JONES G,GAMS W,et al, 2011. The Genera of Hyphomycetes [M]. CBS-KNAW Fungal Biodiversity Centre,P.O.,1-997.SHI W,LUO JP,TING ZH,et al, 2005. Isolation and identification of endophytic fungi ofHuperziaserrata[J]. Chin Trad Herb Drugs,36(2):281-283. [石玮,罗建平,丁振华,等, 2005. 千层塔内生真菌分离鉴定的初步研究 [J]. 中草药,36(2):281-283.]
SUN JQ,GUO LD,ZANG W,et al, 2008. Endophytic fungal diversity and ecological distribution of medicinal plants [J]. Sci Chin Life Sci,38(5):475-484.
TEJESVI MV,KAJULA M,MATTILA S,et al, 2011. Bioactivity and genetic diversity of endophytic fungi in Rhododendron to mentosum Harmaja [J]. Fung Divers,47(1):97-107.
THANGAVELU MUTHUKUMAR,KANDASAMY PRABHA, 2012. Fungal associations in gametophytes and young sporophytic roots of the fernNephrolepisexaltata[J] . Acta Bot Croat,71(1):139-146.
WU TY,YANG L,XU MM,et al, 2012. Diversity and correlation of endophytic fungi and rhizosphere fungi isolated fromDrynariaroosii[J]. Guangdong Agric Sci,39(17):157-159. [巫婷玉,杨丽,许明敏,等, 2012. Diversity and correlation of endophytic fungi and rhizosphere fungi isolated fromDrynariaroosii[J]. 广东农业科学,39(17):157-159.]XU XY, MA YJ, 2001. K-release from potassium-containing minerals in soil and it’s significance for plant nutrition [J]. Chin J Soil Sci,32(4):173-176. [徐晓燕,马毅杰, 2001. 土壤矿物钾的释放及其在植物营养中的意义 [J]. 土壤通报,32(4):173-176.]
ZHAN SF,FAN YF,GAN JL,et al, 2007. A study on endophytic fungi inPterismultifidaⅠ:Isolation and identification of endophytic fungi [J]. J Fung Res,5(4):195-197. [詹寿发,樊有赋,甘金莲,等, 2007. 凤尾蕨内生真菌的研究I-菌种分离及其分类鉴定 [J]. 菌物研究,5(4):195-197.]ZHAN SF,PENG Q,CHEN Y,et al, 2012. Diversity of endophytic fungi inPyrrosiasheareri(Bak.) Ching and the microstructure of its ground parts [J]. J Southern Agric,43(9):1 320-1 324. [詹寿发,彭琴,陈晔,等, 2012. 庐山石韦内生真菌多样性及地下部分显微结构观察 [J]. 南方农业学报,43(9):1 320-1 324.]ZHANG JC,WANG ZG,ZHANG HY,et al, 2010. The summarize about recent research process on fern endophyte [J]. Chin Agric Sci Bull,26(20):70-72. [张君诚,王铮敏,张杭颖,等, 2010. 蕨类植物内生菌研究进展 [J]. 中国农学通报,26(20):70-72.]ZOU WX,TAN RX, 2001. Recent advances on endophyte research [J]. Acta Bot Sin,43(9):881-892. [邹文欣,谭仁祥, 2001. 植物内生菌研究新进展 [J]. 植物学报,43(9):881-892.]
Diversity of culturable endophytic fungi separated from seven common ferns in potassium mine areas in Longnan, South Jiangxi Province
QIAN Xu, GAN Hui-Yun, DU Yong-Tao, ZHANG Xian, CHEN Ye*, FAN You-Fu
(SchoolofPharmacyandLifeSciences,JiujiangUniversity, Jiujiang 33200, China )
Abstract:Plant endophytic fungi is a special kind of microorganisms mutualistic ally symbiosed with host plants. The special microbial resources have attracted tremendous interests in recent years, because of their potential to address problems associated with enhanced plant growth in environmental stress of mines and to improve the efficiency of ecological restoration. With a long peoriod of evolution, endophytic fungi and their host plants have formed a closemutual symbiotic relationship. Many beneficial biological effects have been made on the host plants, and they have the important ecological significance in plant community succession process. It is widely attracted using plants-microorganisms to enhance plant growth in environmental stress of mines, and to improve biological restoration efficiency, and its significance to host plants has become a new hot issue. In order to make clear the species diversity, community composition and ecological distribution of endophytic fungi, we carried out our work among 7 species of common ferns in potassium mine areas in Longnan, South Jiangxi Province. They are Dicranopteris linearis, Woodwardia japonica, Athyrium yokoscense, Stenoloma chusanum, Lygodium japonicum, Asplenium sarelii and Pteris multifida. And we analyzed the diversity of endophytic fungi, using the method of tissue isolation, morphological identification. The results showed that 377 strains of endophytic fungi were isolated from 7 species of plants, and the separation rate ranged from 2.50% to 4.52%. All of 377 strains belong to 25 genera, and among them, Alternaria, Aspergillus, Cladosporium and Verticillium,which are found in all these 7 fern species, indicated that they were dominant, but there were obvious differences in the distribution in each fern species. The total separation rate and the colonization rate of entophytic fungi from leaves are generally greater than that from the rhizomes, but there are some differences in the total separation rate and the colonization rate from different parts of each fern species. The diversity indexes of endophytic fungi range from 0.502 to 0.867 in the seven species of ferns. It indicated that there was high diversity of endophytic fungi in potassium mines and the low similarity of endophytic fungi in different ferns in the same mine areas. The similarity coefficients of the endophytic fungi in the seven species of ferns were from 0.189 to 0.587. The results of this study would not only enrich germplasm resources of plant endophytic fungi, but also would offer basic data for further research on the mechanism that endophytic fungi could strengthen the adaptation of the host plant on mineral nutrition and stress physiology in potassium mine areas.
Key words:potassium mine areas, endophytic fungi in ferns, dominant endophytic fungi, diversity index, similarity coefficient
DOI:10.11931/guihaia.gxzw201408021
收稿日期:2014-12-14修回日期: 2015-01-14
基金项目:国家自然科学基金(31360064, 51264014)[Supported by the National Natural Science Foundation of China(31360064, 51264014)]。
作者简介:钱旭(1992-),男,河南郸城人,研究方向为真菌多样性,(E-mail)1321483416@qq.com。 *通讯作者: 陈晔,教授,研究方向为植物内生真菌多样性,(E-mail)chenyejjtc@126.com。
中图分类号:Q939.5
文献标识码:A
文章编号:1000-3142(2016)03-0342-07
钱旭,甘会云,杜勇涛,等. 龙南钾矿区常见蕨类植物可培养内生真菌的多样性 [J]. 广西植物, 2016, 36(3):342-348
QIAN X,GAN HY,DU YT,et al. Diversity of culturable endophytic fungi separated from seven common ferns in potassium mine areas in Longnan, South Jiangxi Province [J]. Guihaia, 2016, 36(3):342-348
