30个马铃薯卷叶病毒CP基因序列分析
2016-06-22韩树鑫张俊华白艳菊张威高艳玲范国权张抒申宇
韩树鑫,张俊华*,白艳菊,*,张威,,高艳玲,,范国权,张抒,申宇
1.东北农业大学,黑龙江哈尔滨1500302.黑龙江省农业科学院植物脱毒苗木研究所,黑龙江哈尔滨150086
30个马铃薯卷叶病毒CP基因序列分析
韩树鑫1,张俊华1*,白艳菊1,2*,张威1,2,高艳玲1,2,范国权2,张抒2,申宇2
1.东北农业大学,黑龙江哈尔滨150030
2.黑龙江省农业科学院植物脱毒苗木研究所,黑龙江哈尔滨150086
摘要:通过对含有PLRV的不同样品RT-PCR扩增,成功地克隆了PLRV CP基因,经比对样品的CP基因序列,核酸一致率为99.55%,仅发现了来自于广东的样品与其它样品有5个碱基的差异。试验样品与国内已报道的PLRV CP基因进行进化分析,不同地区的PLRVCP基因可分为A,B组,且有区域性特征。来自广东、云南及贵州的样品在两组中均有分布,但来自于内蒙古的样品仅聚类在A组。而中国样品与其他国家PLRV CP基因相比较,经进化树分组后,主要分为S1和S2两组,其中S1组中,包含了所有来自于我国的样品和绝大多数其它样品,且无规律可循;而S2组中的样品仅只来自于埃及和波兰。总体上,PLRV CP序列的差异仍然较小,因此,目前PLRV病毒的种群基因较稳定。
关键词:马铃薯;卷叶病毒;CP基因;序列分析
马铃薯卷叶病毒(Potato leaf roll virus,PLRV),属于黄症病毒科(Luteoviridae)马铃薯卷叶病毒属(Polerovirus),主要侵染茄科植物,能够引起马铃薯卷叶、黄化、矮缩、僵化及块茎网状坏死等病症,导致产量和品质下降。PLRV是单分子正义ssRNA病毒,全基因组长610 Kb,其中第4个读码框(ORF3)是CP的编码序列,长627 bp,编码产物为23 kDa的多肽[1,2]。该病毒于1916年被发现,病毒主要集中于寄主体内的维管束中,且含量低[3]。PLRV在田间以蚜虫持久性传播,持毒时间久,传播距离远[4]。通常蚜虫在马铃薯田中的密度比临近作物高,在马铃薯生长期间蚜虫密度逐渐增大,PLRV病毒感染率也随之增加[5]。由于PLRV在中国马铃薯生产区普遍发生[6],而病毒病防治没有特效药剂,一旦感病则难以控制,所以病毒检测非常重要,通过检测剔除感病植株或切断传播途径防止扩散。
中国是一个幅员辽阔、地理气候等环境复杂的国家。目前,马铃薯从南到北,从东到西,全国范围内广泛分布,PLRV在各地都有发生,且新的PLRV也会随种子资源的引进而发生同步引进的风险。围绕PLRV的CP基因进行的研究比较普遍,但都是以地区划分,缺乏衔接和与国际上整体PLRV种群间的比对分析,而且,PLRV病毒是否随环境和气候的改变而有所不同尚无系统研究和阐述。PLRV的CP基因还是PLRV检测单克隆抗体的重要来源。本研究的开展有利于了解PLRV病毒的变异、重组,可以进一步了解我国PLRV的种群构成,有助于了解病毒的发生、流行特点,并为在生产中PLRV的检测鉴定、防治及其致病机理、抗病基因工程育种的研究提供理论支持和参考。
1 材料与方法
1.1试验材料
试验使用的马铃薯材料来自于黑龙江省农科院植物脱毒苗木所,于2012~2014年采集,经ELISA检测感染PLRV的马铃薯样品30份(表1)。
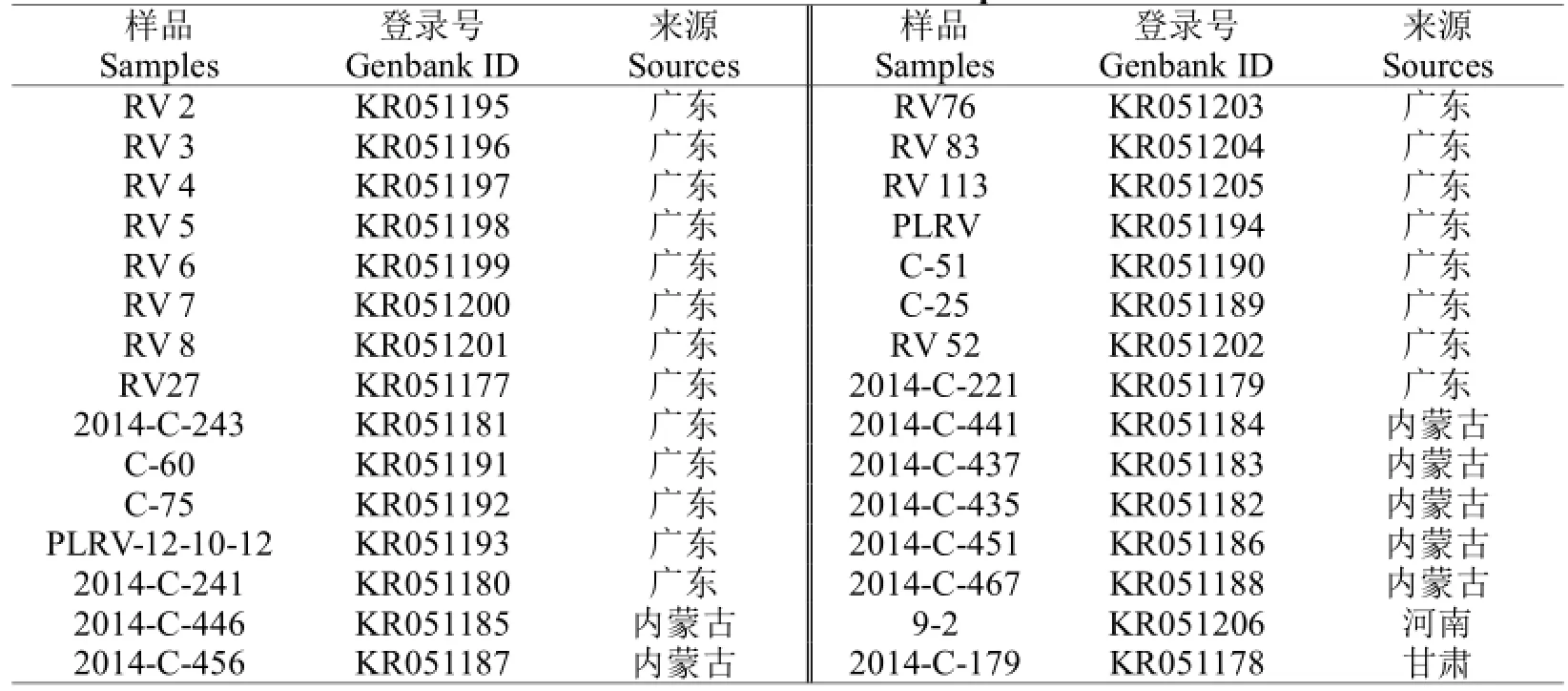
表1 样品来源Table 1 Sources of samples
1.2引物的设计与合成
根据已有的马铃薯卷叶病毒基因的保守序列,及常用于检测Total RNA质量的内参基因NAD5的保守序列,本试验共使用三对引物,其中PLRV-F/R用于鉴定马铃薯样品是否感染PLRV病毒,NAD5-F/R用于鉴定RNA是否符合试验要求。同时设计了引物CP-PLRV-F/R用于扩增PLRV的CP序列。引物由上海生工生物工程有限公司合成(表2)。
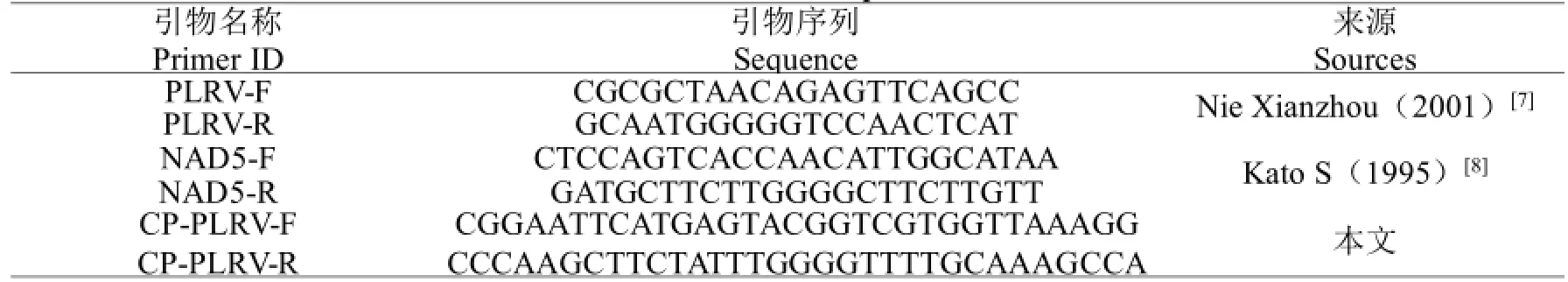
表2 引物列表Table 2 The list of primers
1.3总RNA的提取
植株总RNA的提取按照Trizol(Invitrogen)试剂说明书步骤进行。提取试验的所有离心操作均在4℃下完成,应用分光光度法测定RNA浓度及纯度,用电泳法检测RNA完整性。
1.4RT-PCR检测及PLRV的CP序列扩增
1.4.1cDNA第一链的合成以感染PLRV的马铃薯叶片总RNA为模板,反转录合成cDNA第一链,反应步骤为:在PCR管中加入模板RNA4 μL,Random primer 1 μL,用DEPC水补齐至10 μL。65℃加热5 min。冰上冷却2 min后像PCR管中加入5×Buffer 5 μL,dNTP 1.25 μL(10 mmol/L),RNasin 0.5 μL (40 U/uL),M-MLV1 μL(200 U/μL),用DEPC水补齐至25 μL。在PCR仪上按下列条件进行反转录反应:37℃1 h,70℃15 min(酶失活)后,冰上冷却,-20℃保存备用。
1.4.2RNA质量和马铃薯PLRV病毒的双重RT-PCR检测以cDNA第一链为模板,使用引物NAD5-F/R 和PLRV-F/R进行双重PCR扩增,使用1.5%的琼脂糖凝胶进行电泳检测。双重RT-PCR鉴定PLRV病毒及RNA质量的步骤为:在PCR管中加入制备的cDNA2 μL,dNTP 2 μL(2.5 mmol/L),rTaq 0.25 μL (5 U/μL),10×PCR Buffer 5 μL,上下游引物(10 mmol/L)各0.5 μL,用DEPC水定容到25 μL的反应体系。PCR扩增条件:94℃2 min预变性后进行35个循环(94℃变性20 s,55.5℃退火30 s,,72℃延伸30 s),72℃延伸5 min,4℃存放。
1.4.3CP基因的扩增PLRV的CP基因的扩增与马铃薯PLRV的RT-PCR检测除退火温度为60℃外,其余步骤相同,扩增完成后同样使用1.5%的琼脂糖凝胶进行电泳检测。
1.5PCR产物测序及序列分析
扩增产物经琼脂糖凝胶电泳检测后,交由华大基因进行测序,使用DNAMAN软件对测序结果进行拼接后对比分析,并使用MAGA6软件,通过最大相似法(Maximum Likeihood),在1000个重复下(Bootstrap trials)进行系统进化分析。
2 结果与分析
2.1双重RT-PCR检测结果
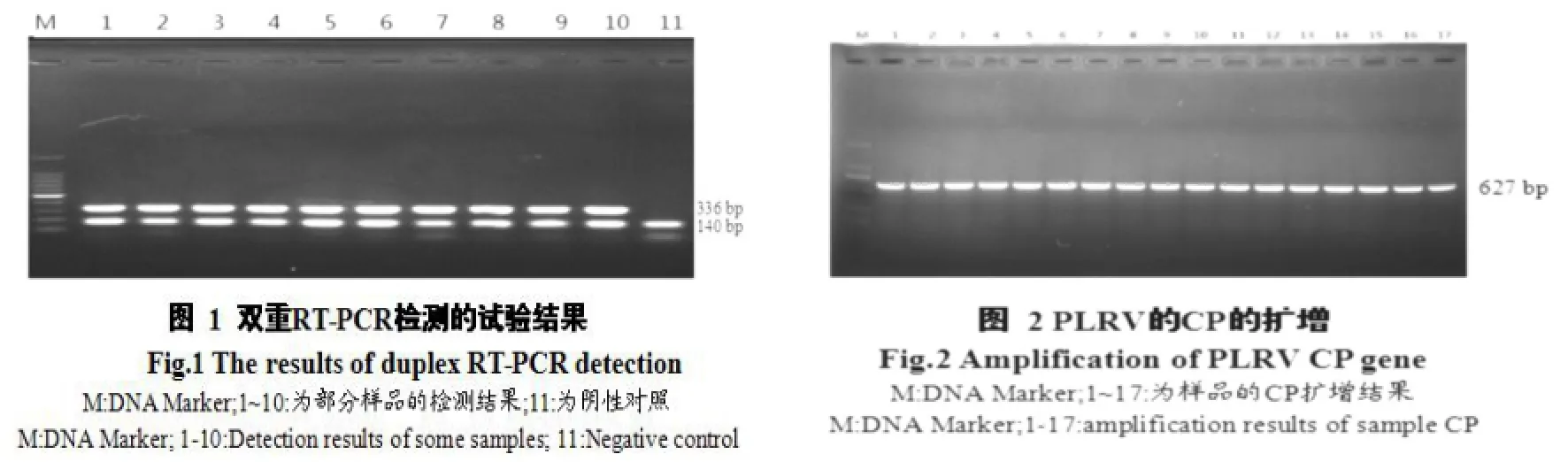
对经DAS-ELSIA的检测受PLRV病毒侵染的30份样品(表2)进行总RNA提取和双重RT-PCR扩增检测,结果显示(图1),可以扩增到2条不同的电泳条带,条带大小分别为336 bp及140 bp,片段大小与试验设计相符,表明样品中的总RNA含有PLRV病毒的RNA且RNA质量良好,进一步验证了样品感染了PLRV病毒。
2.2CP序列扩增结果
对筛选出的含PLRV病毒的样品进行CP基因序列经RT-PCR扩增,得到一条长约630 bp的扩增产物,大小与引物设计相一致,且条带单一,明亮。
2.3CP序列的测定与分析
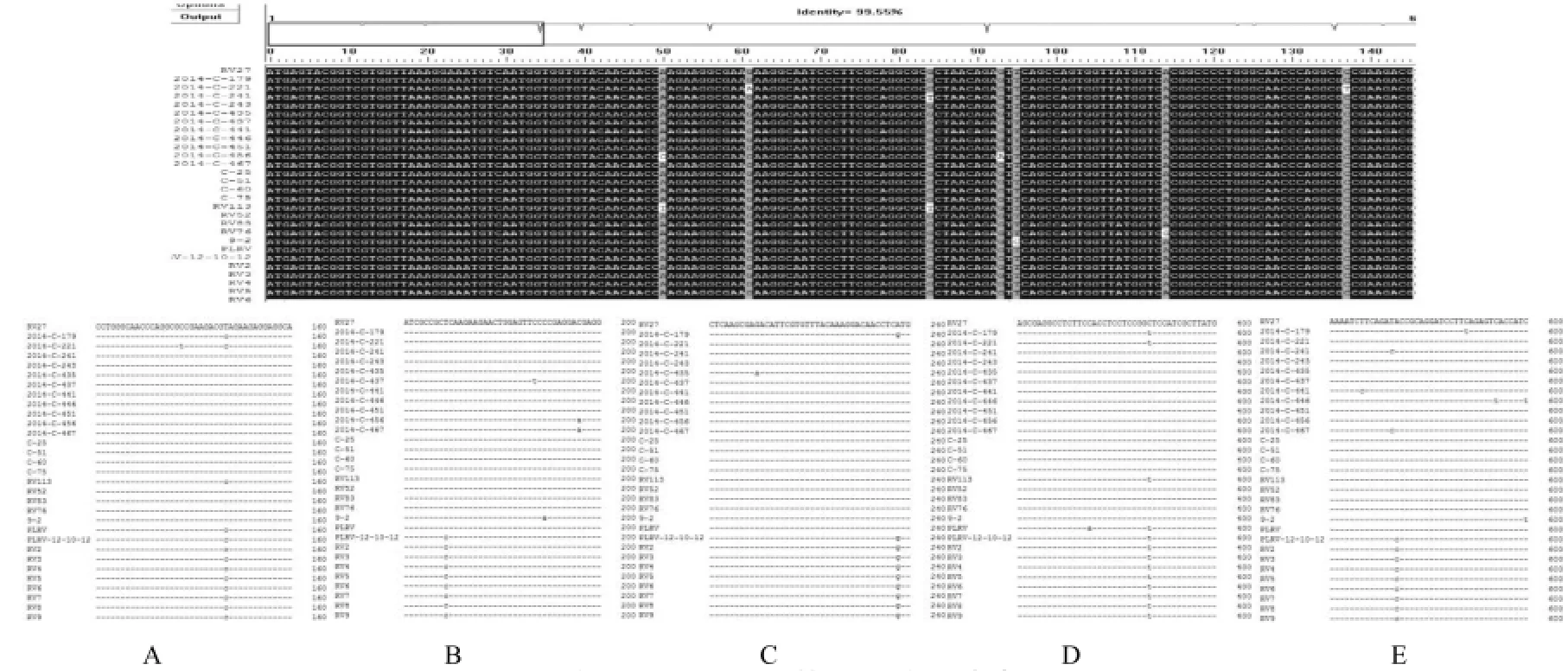
图3 检测样品的PLRV CP基因的核苷酸序列比对结果Fig.3 The contrast of PLRV CPgene alignment sequences of samples
PLRV病毒CP基因的PCR扩增产物经测序后,使用DNAMAN软件进行比对,结果表明,PLRV 的CP序列全长共627个碱基,样品序列间核酸一致率达到99.55%,说明此次进行序列扩增PLRV样品间的序列差异不大,基本没有发生的较多碱基参与重组及突变现象(图3)。另外,在序列比对中可以看到,样品间除了个别的碱基有不规律的改变,PLRV-12-10-12和RV2-RV8这些样品,在CP基因序列中的第143、169、238、387、574位处碱基发生了有规律的发生改变。PLRV-12-10-12和RV2-RV8这些样品的碱基与其它样品相比,第143位处,T变为了C、第169位处,T变为C、第238位,A变为G、第387为,C变为T、第574位,A变为C。虽然,这些突变在整个基因上的分布距离较远,并不能聚集成簇,且占整个基因序列的比例极小,但通过序列分析,我们还是能发现,这些样品与其它样品间,在序列上的差异是有规律可循的。第三,从样品采集的区域来看,发生碱基突变的PLRV-12-10-12和RV2-RV8这些样品的采集区域集中在广东地区。而其它的样品则来源于内蒙古、河南、甘肃等地。这也说明我国的PLRV的分布不但有其遗传因素的影响,来自于地域的自然环境的影响也有,但很小。
2.4中国PLRV病毒CP基因系统进化分析
为进一步明确我国不同地区间PLRV病毒的种群状态,使用MAGA 6.0软件并选取本试验样品中有代表性的样品与其它已报到的我国不同地区的CP序列(表3)进行系统进化比较分析(图4)。试验样品与我国不同地区的CP序列遗传进化分析表明,所有的序列可分为2组,我们将他们分别编为A组,B组(图4),从这两组样品的来源看,虽然同源性很高,却仍有区域性特征,来自广东、云南及贵州的样品在各组中均有分布,但来自于内蒙古自治区的样品则未在B组中被发现。另外,A组中,所有样品聚类为一个群组,内蒙古自治区的样品占大多数,而B组中,聚类成两个亚组,样品主要来自于广东、贵州、云南,地域间差异微小。
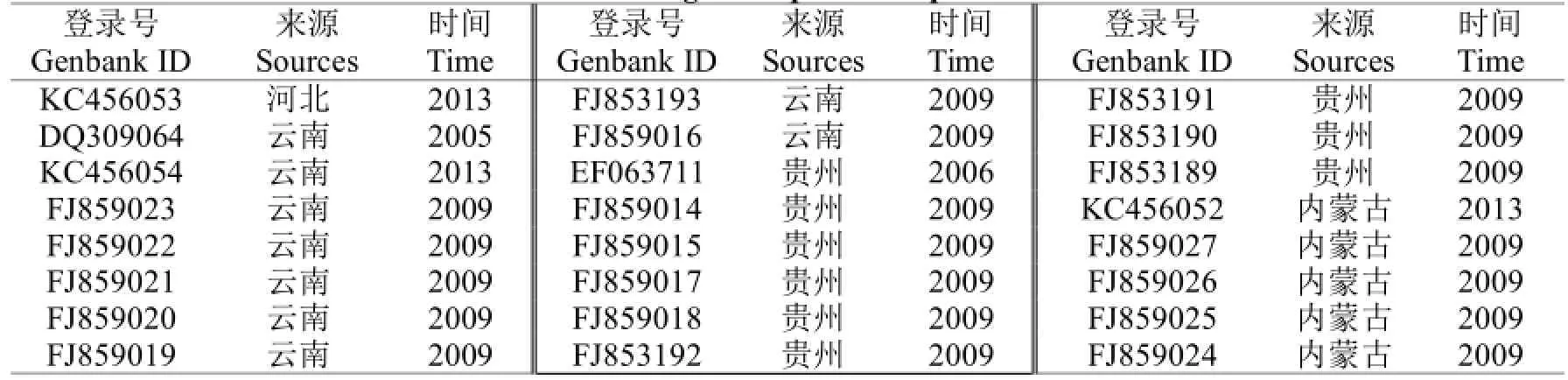
表3 国内已报道PLRV CP基因序列Table 3 PLRV CP gene sequences reported in China
2.5中国PLRV病毒CP基因序列与国外的比较
从国内样品的系统进化分析出来的两个组群中,以遗传关系的远近为标准选取其中有代表性的样品17个与世界其它地区的PLRV的CP基因序列进行系统进化分析(图5),在Gene Bank共查得国外序列49个(表4),分析结果表明,已报到的世界范围的PLRV,其CP序列的整体同源性同样较高,但仍有一定差异。经进化树分组后,可主要分为S1和S2两组,其中S1组中,包含了所有来自于我国的样品和绝大多数其它样品,且无规律可循,而S2组中的样品则只来自于埃及和波兰,由此,可说明目前世界范围内PLRV的种群基因较稳定。
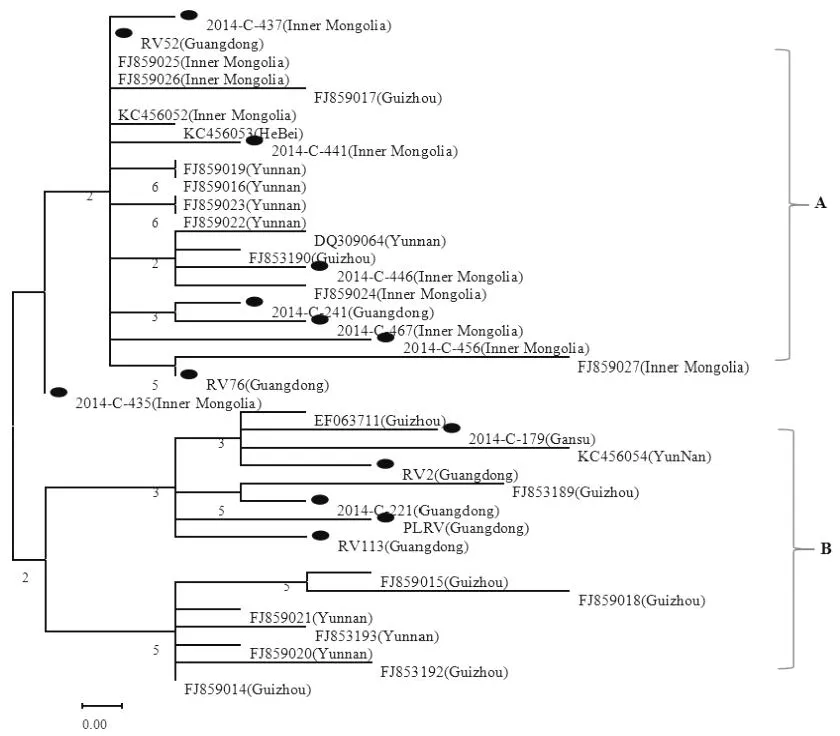
图4 试验样品与国内样品的CP核苷酸序列构建进化树Fig.4Thephylogenetictreeof CPuncleotidesequences ofthe testsampleandothersin China
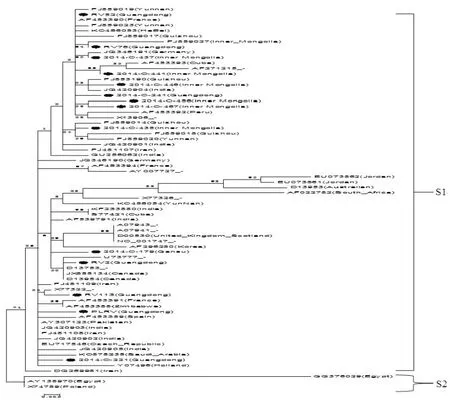
图5 试验样品与各地样品的PLRV核苷酸序列构建进化树Fig.5 The phylogenetic tree of PLRV uncleotide sequences from thetestsamplesandothers
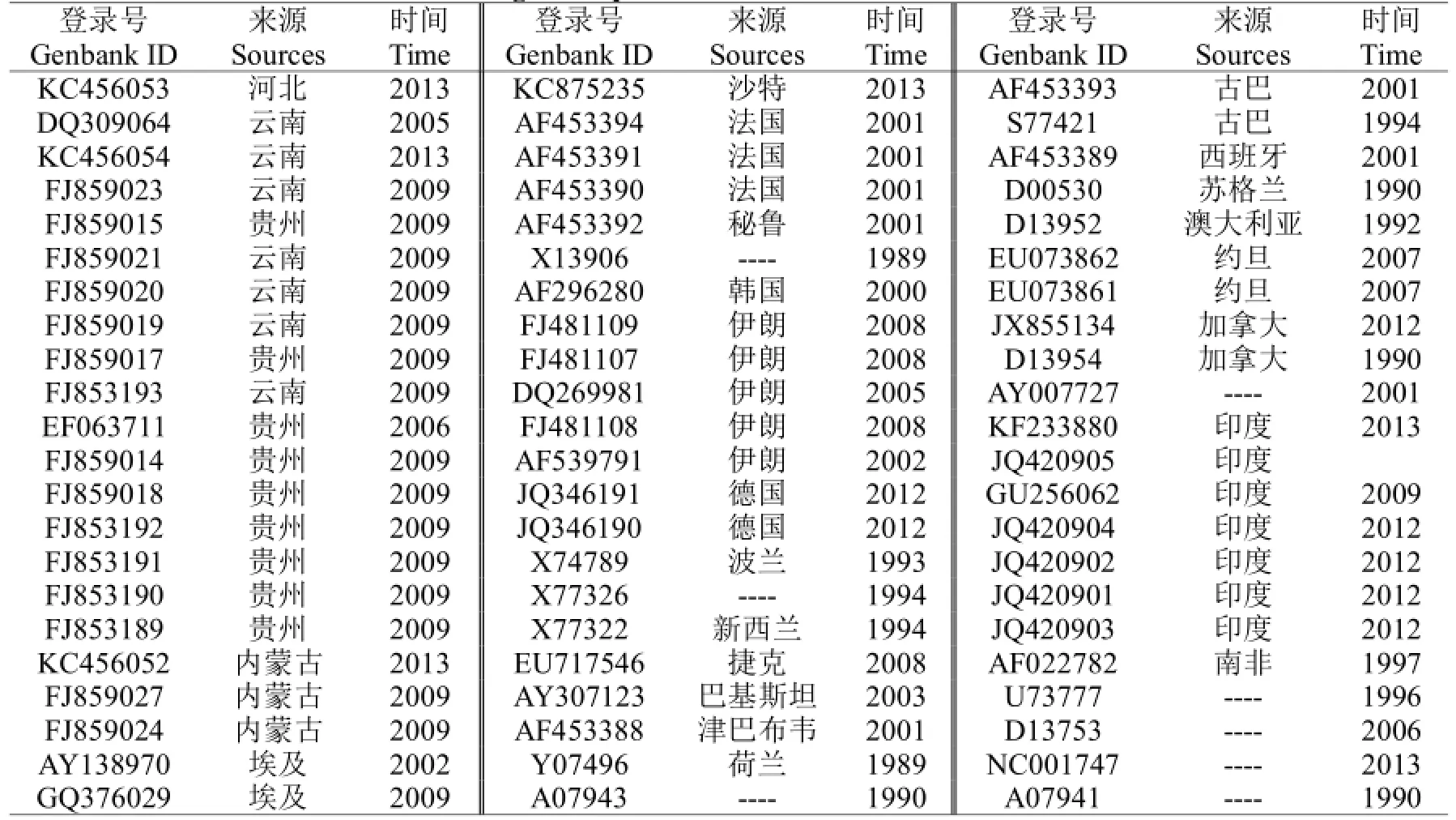
表4 各地的不同来源PLRV CP基因序列Table 4 PLRV CPgene sequences from different sources around world
3 讨论
目前,马铃薯卷叶病毒是造成马铃薯减产和品质下降的主要病害之一,各地发生普遍且不同的地区由于生产条件及自然条件不同,导致植株症状也不尽相同,从而对生产造成了严重的危害[9]。随着转基因技术的不断发展和完善,利用植物病毒保守区基因的siRNA进行转基因育种,从而使品种对相应植物病毒的抗性增强[10],由于马铃薯卷叶病毒CP序列的相对保守,所以人们对其的研究也随之增多[11-13]。另外,由于在生产中对PLRV检测的方法主要以血清学方法为主,而制备检测所用的单克隆抗体大多数是以PLRV的CP蛋白为识别目标,所以一旦有部分PLRV的CP基因编码发生变化,进而导致相应PLRV的CP蛋白发生变化,导致漏检,则会对我们下一步的工作造成影响[14,15]。
本试验对各地的受PLRV侵染的CP基因序列样品进行了RT-PCR扩增和测序。试验目的是检测我国不同地区的马铃薯卷叶病毒是否存在有CP基因序列测序出现重组或突变的个体及群体,并通过与世界其它地区PLRV的CP基因进行比对分析,进而对我国PLRV种群的现状进行分子生物学评价。结果表明本次扩增的样品的CP基因全长627 bp。利用DNAMAN软件对实验样品的CP基因序列进行分析,发现试验样品的CP序列间差异极小,核酸一致率达到99.55%,基因保守性高,这与大多数研究的结果相同[16]。但广东地区的样品与其它地区样品CP基因经比对后,有5个位置的碱基发生了有规律的变化。这说明,虽然CP基因保守性高,可有规律的突变在不同地区的PLRV CP基因中还是会发生的。
通过MAGA6.0对实验样品CP序列与Genbank中已知来自于我国的所有PLRVCP基因序列进行进化分析,发现虽然我国国内PLRV差异极小,但仍可分为A,B两组,其中A组中内蒙古自治区样品占绝大多数,而B组中不含来自于内蒙古自治区的CP基因,另外,广东、云南、贵州的样品则在两组中均有出现,这可能与该病毒受当地环境或气候长期影响有关,也与马铃薯种植特点有关,广东为马铃薯种薯输入省份,PLRV可随各地种薯到达此地,其基因型可能相对复杂;内蒙古、贵州、甘肃、云南为种薯输出省份,且云南、贵州的种薯通常不会北调,适宜这个地区的种子资源一般也不适宜北方,因此,这个区域的PLRV病毒则不会迁移北方,而北方的PLRV病毒会随种薯调运或种子资源到达南方,那么,B组中有一个亚组只有贵州和云南的样品的情况就很容易理解了。
挑选有代表性中国PLRVCP基因序列与Genbank中已上传的世界其它地区的基因进行进化分析,我国的PLRV与世界上大多数地区的样品被分在了同一大组S1组内,而另外的一组S2组中,仅有来自于埃及和波兰的样品,埃及地理位置和气候具有特殊性,其登录号为GQ376025的PLRV与其他分离物同源性略远也许反应出这一原因,但波兰的样品聚类到S2则没有合适的解释,尚需进一步研究。
本次比对的所有CP基因序列的时间跨度从1989年至今,虽然,时间跨度长达近30年,但作为极易发生突变的物种——病毒,却未发现PLRVCP基因序列发生较大突变或重组,这说明我国PLRV种群与国外的PLRV同属于一个来源,且在相当长的时间内,受自然环境和遗传因素的双重压力下,其遗传性状还是很稳定的。这进一步的表明,PLRV的多面体分子结构与其它病毒相比是稳定的。PLRV CP基因间差异,只是由于不同地区的自然环境导致单个核酸位点改变。这些结论与冯光惠等、吴兴全等、颜永杰等根据我国不同地区的PLRV CP基因与国内外其它PLRV CP基因进行分析后得出的结论是相同的,即不仅我国的PLRV CP基因具有高度的同源性及核酸一致率,且未发生改变其CP蛋白性状的突变[17-19]。目前仅突尼斯发现了PLRV的变异株[20],但变异部位在P0区,CP端无特殊变化。
4 结论
本研究通过测试样品与国内为样品CP序列的比对得到一系列对PLRV检测、防治和抗病育种有指导性意义的结论,但由于可供分析的国内外样品数量有限,不排除有PLRV的CP基因突变株的存在。
参考文献
[1]郭志华,孙毅,张效梅,等.马铃薯卷叶病毒(PLRV)内蒙古分离物基因间隔区(IS)的克隆与序列分析[J].华北农学报,2007,22(6):19-23
[2]隋炯明,何心凤,郭真,等.马铃薯卷叶病毒缺失突变CP基因的原核表达及抗血清的制备[J].园艺学报,2012,39(10):1949-1957
[3]张鹤龄.马铃薯卷叶病毒(PLRV)基因组研究进展[J].中国病毒学,1996,11(1):1-8
[4]Srinivasan R,Alvarez JM,Bosque-Pérez NA,et al. Effectof an Alternate Weed Host,Hairy Nightshade,Solanumsarrachoides,on the Biology of the Two Most Important Potato Leafroll Virus(Luteoviridae:Polerovirus)Vectors,Myzus persicae and Macrosiphumeuphorbiae(Aphididae:Homoptera)[J].Environmental Entomology,2008,37(2):592-600
[5]Srinivasan R,Alvarez JM,Cervantes F. The effect of an alternate weed host,hairy nightshade,Solanum sarrachoides (Sendtner)on green peach aphid distribution and Potato leafroll virus incidence in potato fields of the Pacific Northwest[J]. Crop Protection,2013,46(46):52-56
[6]陈兆贵,施招婉,陈静敏.惠州马铃薯卷叶病毒病原分子鉴定及序列分析[J].作物杂志,2012(4):45-48
[7]Nie X,Singh R. Anovel usage of random primers for multiplex RT-PCR detection of virus and viroid in aphids,leaves,and tubers[J]. J Virol Methods,2001,91(1):37-49
[8]Kato S,Shimamoto Y,Mikami T. The apple mitochondrial atp9 gene:RNA editing and cotranscription with exons a and b of the nad5 gene[J]. Physiologia Plantarum,1995(3):572-575
[9]张威,白艳菊,罗丽环,等.三种马铃薯病毒多重RT-PCR检测体系建立[J].东北农业大学学报,2014(5):13-19
[10]张陈明,胡宗利,陈国平.根癌农杆菌介导转化马铃薯与抗病毒基因工程[J].生物技术学通报,2008(6):30-35
[11]周云,杨永智.马铃薯卷叶病毒基因组保守序列片段的RT-PCR扩增[J].青海大学学报,2008,26(2):16-19
[12]Zhuo T,Li YY,Xiang HY,et al. Amino acid sequence motifs essential for P0-mediated suppression of RNAsilencing in an isolate of potato leafroll virus from Inner Mongolia[J]. Molecular Plant-Microbe Interactions,2014,27(6):515-527
[13]董代幸.乌鲁木齐地区马铃薯病毒和类病毒的分子鉴定及检测技术研究[D].乌鲁木齐:新疆农业大学,2010
[14]李楠楠,左玉玲,隋炯明,等.重组CP多克隆抗体在马铃薯卷叶病毒DAS-ELISA检测中的应用[J].华北农学报,2011,26(6):85-88
[15]Wróbel S. Modification of ELISA by Replacing Incubation of Microtiter Plates in an Incubator with Their Shaking in PVY,PVM and PLRVDetection[J]. American Journal of Potato Research,2014,91(4):354-362
[16]丁铭,方琦,李婷婷,等.马铃薯卷叶病毒云南分离物外壳蛋白基因的克隆与序列分析[J].植物病理学报,2006,36(5):473-476
[17]冯光惠,杜虎平,李夏隆.陕北地区马铃薯卷叶病毒的RT-PCR检测与序列分析[J].湖北农业科学,2014,53(19):4734-4737
[18]颜永杰,吴宽,谢海峰,等.陕西马铃薯卷叶病病原的分子生物学鉴定[J].西北农林科技大学学报:自然科学版,2010(5):87-92
[19]吴兴泉,谭晓荣,陈士华,等.马铃薯卷叶病毒福建分离物的基因克隆与序列分析[J].河南农业大学学报,2006,40(4):391-393
[20]Khouadja FD,Rouzé-Jouan J,Guyader S,et al. Possible correlations between the characteristics of Potato leafroll virus isolates occurring in different geographical regions in Tunisia[J]. Phytoparasitica,2014,42(2):259-267
Analysis on CPGene Sequences of Thirty Potato Leaf Roll Virus
HANShu-xin1,ZHANGJun-hua1*,BAIYan-ju1,2*,ZHANGWei1,2,GAOYan-ling1,2,FANGuo-quan2,ZHANGShu2,SHENYu2
1. Northeast Agricultural University,Harbin 150030,China
2. Heilongjiang Academy of Agricultural Sciences,Harbin 150086,China
Abstract:We used RT-PCR to amplify the different samples with PLRV and successfully cloned PLRV CP gene by means of comparing CP gene sequence of samples to discover 99.55%of the nucleic acid concordance rate,there was only five bases difference between the sample from Guangdong and others. We had analyzed the test samples and other recorded PLRV CP genes in China and divided them from different districts into Group A,Group B. The results showed that samples from Guangdong,Yunnan and Guizhou distributed in two groups but only the samples from Inner Mongolia cluster in Group A. The PLRV CP genes from Chinese samples compared with other countries to be divided into Group S1 and Group S2. In Group S1,there were unordered samples from China and most of other countries;in Group S2,there were only samples from Egypt and Poland. In one word,there were a little difference in PLRV CP sequences so that at present the gene of the PLRV population was stable.
Keywords:Potato;potato leaf roll virus(PLRV);CP gene;sequences analysis
中图法分类号:S4325.32;Q943.2
文献标识码:A
文章编号:1000-2324(2016)03-0353-06
收稿日期:2015-04-20修回日期:2015-08-20
基金项目:现代农业产业技术体系专项资金资助(CARS-10-P14)
作者简介:韩树鑫(1984-),男,在读博士研究生,主要从事植物病理方向的研究. E-mail:pluto789@163.com
*通讯作者:Author for correspondence. E-mail:Podozjh@163.com;Yanjubai@163.com
