U、Cd单一及复合污染对水芹生长和生理特性影响
2016-06-20姚天月李泽华西南科技大学生命科学与工程学院四川绵阳621000西南科技大学核废物与环境安全国防重点实验室四川绵阳621000
姚天月,王 丹*,李泽华(1.西南科技大学生命科学与工程学院,四川绵阳621000;2.西南科技大学核废物与环境安全国防重点实验室,四川绵阳621000)
U、Cd单一及复合污染对水芹生长和生理特性影响
姚天月1,2,王丹1,2*,李泽华1,2
(1.西南科技大学生命科学与工程学院,四川绵阳621000;2.西南科技大学核废物与环境安全国防重点实验室,四川绵阳621000)
摘要:通过土壤盆栽试验研究不同浓度的U、Cd单一及其复合污染对水芹的生长、叶绿素含量、根系活力、细胞膜透性和MDA含量的影响。结果表明:与对照相比,随U、Cd单一及其复合污染处理浓度的增加,水芹的生长受到显著抑制(P<0.05);水芹叶绿素含量逐渐减少,其中以Cd元素对水芹叶绿素b胁迫作用最显著;水芹根系活力在低浓度(15 mg·kg-1)时受到促进,而在高浓度(150 mg·kg-1)时受到抑制;叶片相对电导率处理组数据均显著高于对照组,U胁迫作用强于Cd;叶片丙二醛含量与对照相比随处理浓度的增加呈上升趋势。比较U、Cd单一及其复合处理对水芹的胁迫效应表明,高U高Cd(U150 mg·kg-1+Cd150 mg·kg-1)对水芹生长和生理影响最显著,水芹在低浓度U、Cd胁迫下具有较好的抗逆性,因此,其在低浓度U、Cd单一及其复合污染的土壤修复方面具有潜在应用价值。
关键词:水芹;铀;镉;生理特征
姚天月,王丹,李泽华. U、Cd单一及复合污染对水芹生长和生理特性影响,等.题目[J].农业环境科学学报, 2016, 35(5):871-877.
YAO Tian-yue, WANG Dan, LI Ze-hua, et al. Effects of single and combined pollution of uranium and cadmium on growth and physiological characteristics of Oenanthe javanica[J]. Journal of Agro-Environment Science, 2016, 35(5): 871-877.
核电事业的迅猛发展为我们的生活带来巨大的社会效益和经济效益,同时加剧了人们对铀矿开采的力度。铀是一种核素兼具辐射性和化学毒性[1],铀矿开采后的尾矿,废渣、废水的不合理排放和处理,可诱发严重的以铀元素为主还伴有镉、锌、铜等其他重金属元素的土壤和水体污染。因此,土壤重金属污染并不是单一的,往往是多种重金属并存的复合污染,并伴随元素间的相互作用和生态效应的综合响应[2]。镉是一种有毒的重金属,活性较强且易从土壤中向植物体转移[3],被植物吸收后便会沿着食物链转移,最终危害人体健康。治理重金属污染一直以来都是科研的热点问题,植物修复技术是一项具有前瞻性的修复技术,以植物忍耐和积累或是超量积累某种或某些化学元素的理论为基础,利用植物吸收环境中的污染物从而达到治理环境的目的,以绿色环保成本低廉等多种优势而被应用于治理土壤重金属污染。然而,该技术的成功应用还有赖于更多的科学实验、大田示范工程和技术体系的集成。
水芹属伞形科多年生草本植物,富含多种营养元素,是一种栽培广泛且常见的叶菜类蔬菜。水芹能分泌丰富的脂溶性根系分泌物,可有效去除土壤中有机物[4],对重金属元素具有一定富集能力,属于高富集蔬菜[5]。本研究以水芹为材料,以植物提取技术为主要研究手段,模拟土壤污染盆栽试验,研究U、Cd单一及其复合污染对水芹的生长和生理特性的影响,了解植物抗铀、镉胁迫机理,为植物修复铀镉污染的开展和应用提供理论支持。
1 材料与方法
1.1试验材料
供试植物为水芹[Oenanthe javanica(BI.)DC],种子购于绵阳市农资批发市场。
U的施加形式为UO2(CH3CO2)2·2H2O(分析纯,湖北楚盛威化工有限公司);Cd的施加形式为CdCl2· 2.5H2O(分析纯,四川生科力有限公司)。试验用土采自西南科技大学后花园表层土(0~20 cm),土壤类型为黄壤土,其pH6.98,有机质含量5.86 g·kg-1,碱解氮、有效磷、有效钾含量分别为51.67,7.21,21.21 mg· kg-1。土样经自然风干,剔除草根、碎石,压碎混匀后,过1 cm筛,静置备用。
1.2试验方法
1.2.1水芹苗的培育
选择均匀饱满无病虫害的水芹种子,进行常规大田育苗,严格控制水肥管理,待幼苗生长至约12 cm,选择长势一致且大小均匀的水芹苗备用。
1.2.2试验设计
试验采用土壤盆栽方式,在四川绵阳西南科技大学温室棚进行。首先,每次定量称取3 kg备用土壤移入塑料盆(口径20 cm,高20 cm,底部有孔,带托盘),并向其施浇清水至田间持水量的60%左右,平衡1周后,把相应浓度的UO2(CH3CO2)2·2H2O和CdCl2· 2.5H2O配成溶液,均匀浇灌在栽培盆中(利用托盘将渗出液回收浇灌),处理浓度以纯的U和Cd计,单位为mg·kg-1,以不添加U、Cd元素为对照(CK),其余处理保持一致。待沉降2周后,将各处理下的盆土充分搅拌至U、Cd离子与土壤均匀混合,然后将备用水芹苗(高12 cm)移栽至U、Cd处理的栽培盆中,保持含水量为田间持水量的60%~70%。根据《中国土壤元素背景值》中的数据,我国土壤(A层)铀元素背景值平均为3.03 mg·kg-1,有研究表明我国铀矿废石平均铀含量比土壤本底值高出4~10倍,铀尾矿及其污染地范围内的铀含量普遍在每千克中几十到几百毫克,因此试验浓度设置高(150 mg·kg-1)、低(15 mg·kg-1)两个浓度。U、Cd单一及复合处理浓度如表1所示(共计9个处理),每个处理重复3次,待植物生长90 d后收获。
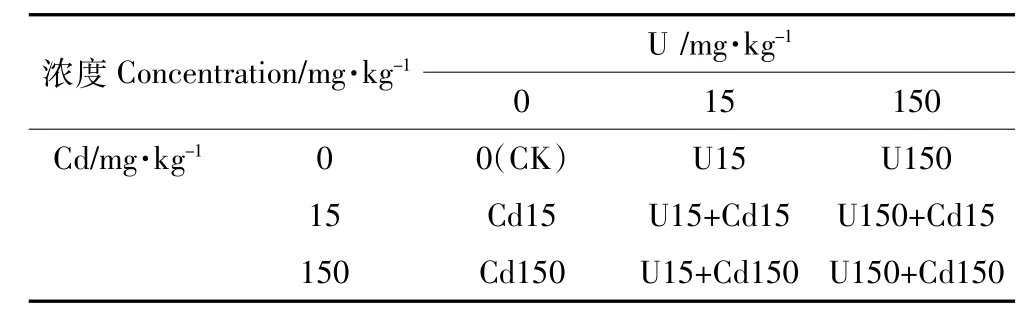
表1 试验处理因素和水平Table 1 Experimental factors and levels
1.3测定方法
1.3.1植物生物量
将植株从不同介质中连根移出,用去离子水充分清洗沥干,用毫米刻度尺对根长、株高、茎粗进行测量,将根与地上部分开,105℃杀青30 min后,75℃下于烘箱中烘干至恒重。
1.3.2植物生理指标
叶绿素总含量测定采用乙醇丙酮浸泡提取法[6],细胞膜透性利用电导仪测定[7],丙二醛(MDA)含量测定采用硫代巴比妥酸(TBA)法[8]。
1.4数据分析
采用Microsoft Excel 2013对试验数据进行平均值和标准差的计算,SPSS 22.0软件进行单因素方差分析和LSD多重比较检验处理间差异程度,采用Origin Pro 9.0绘图。
耐性指数(Tolerance index,TI,%)=实验组根长均值×100/对照组根长均值[9]
2 结果与分析
2.1U、Cd单一处理对水芹生长和生理特性的影响
2.1.1对水芹生长的影响
分析表2得知,U、Cd单一处理下水芹的株高和茎粗均随处理浓度的增加呈下降趋势,与对照CK相比,U、Cd单一处理浓度在150 mg·kg-1条件下,水芹株高分别减少了26.53%和17.01%,茎粗分别减少了44.87%和19.23%,由此可见U对水芹的株高和茎粗的抑制作用显著大于Cd。水芹根长在受到U、Cd单一胁迫时均呈现低浓度(15 mg·kg-1)促进高浓度(150 mg·kg-1)抑制的现象,U浓度15 mg·kg-1条件下水芹根长达最大值8.47 cm,是对照CK的1.28倍,比同浓度Cd处理高出12.39%,耐性指数也达到峰值127.63%,显著高于其他各处理。当处理浓度为150 mg·kg-1时,U、Cd胁迫水芹根长分别减少23.53%, 14.03%。即无论U、Cd处理浓度如何,其对水芹根长的影响作用为U>Cd,此结果与耐性指数反应效果一致。
2.1.2对水芹叶绿素含量和根系活力的影响
由表3可知,U、Cd单一处理会减少水芹叶片叶绿素含量,且抑制作用随处理浓度的增加而增大。水芹叶绿素a、叶绿素b含量与U、Cd浓度呈负相关关系,相关系数分别为-0.908(U对叶绿素a)、-0.996(U对叶绿素b)、-1.000*(Cd对叶绿素a)、-1.000*(Cd对叶绿素b)。可见Cd与水芹叶绿素含量相关性比U强。当U、Cd浓度为150 mg·kg-1时,水芹叶绿素a含量分别减少18.34%、21.97%,叶绿素b含量分别减少20.78%、29.02%。可见叶绿素含量减少作用为Cd>U。此外,叶绿素a/b的值在处理浓度低时降低,处理浓度高时升高,说明叶绿素b含量降幅大于叶绿素a,可见水芹叶绿素b对U、Cd胁迫变化更敏感,因而叶绿素b含量可以作为评价水芹耐受U、Cd胁迫能力大小的晴雨表。根是植物吸收水分和养分的主要器官,是植物营生的根本,根系活力的改变直接影响地上部茎叶的生长。分析表3可知,随U、Cd单一处理浓度的增加,水芹根系活力整体呈现先升高后降低的趋势,与水芹根长所受胁迫效应一致。在Cd浓度15 mg·kg-1处理下出现峰值2.79 mg·g-1·h-1,是对照的1.19倍,在U浓度150 mg·kg-1处理下出现最小值,且当处理浓度相同时U、Cd单一处理差异不显著,说明U、Cd对水芹根系活力的影响主要取决于浓度的大小。在低浓度胁迫下,根系能够保持较高的根系活力,保持较好的吸收和运输等能力。
2.1.3对水芹细胞膜透性和MDA含量的影响
由图1可知,U、Cd单一处理下水芹叶片的相对电导率随处理浓度的增加呈逐渐增大趋势,各处理组数据均显著高于对照。U浓度150 mg·kg-1时水芹叶片相对电导率达各处理峰值88.33%,是对照的4.48倍,是同浓度Cd处理的1.62倍,可见U、Cd对水芹细胞膜透性影响为U>Cd,U更能使水芹细胞膜过氧化,增大膜透性,使胞内电解液外渗增加。MDA是膜质过氧化主要产物之一,如图2所示,水芹叶片MDA含量随U、Cd单一处理浓度的增加呈上升趋势,U、Cd浓度在150 mg·kg-1条件下水芹MDA含量分别为15 mg·kg-1处理下的0.99倍和1.06倍,在P<0.05水平上差异显著,然而低浓度(15 mg·kg-1)处理与对照组CK差异不显著,可见水芹对低浓度U、Cd毒害作用具有一定耐受性。此外,在P<0.05水平上比较处理浓度相同的U、Cd元素对水芹叶片MDA含量的影响,差异不显著,说明两者对水芹叶片MDA含量影响无明显差异(P>0.05)。

表2 U、Cd单一处理对水芹生长的影响Table 2 Effects of U and Cd single treatments on growth of Oenanthe javanica
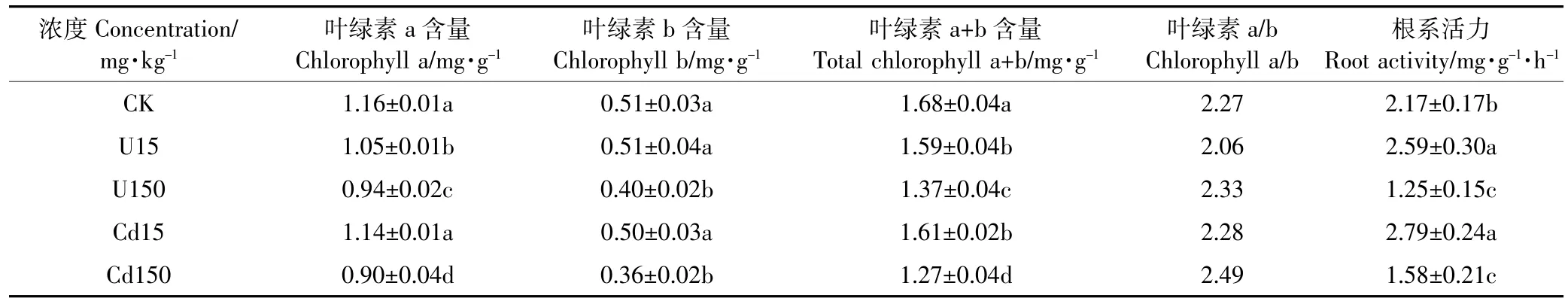
表3 U、Cd单一处理对水芹叶绿素含量和根系活力的影响Table 3 Effects of U and Cd single treatments on chlorophyll content and root activity of Oenanthe javanica
2.2U、Cd复合处理对水芹生长和生理特性的影响
2.2.1对水芹生长的影响
U、Cd复合处理下水芹的株高、茎粗和根长均受到抑制,处理浓度越高对水芹生长抑制作用越强。分析表4可知,水芹生长各指标数据在低U低Cd(U 15 mg·kg-1+Cd15 mg·kg-1)复合处理下最大,高U高Cd (U150 mg·kg-1+Cd150 mg·kg-1)复合处理下最小,前者是后者的1.67~2.55倍,差异显著(P<0.05)。水芹株高对U、Cd复合处理胁迫响应最显著(P<0.05),各处理差异达到显著水平(P<0.05)。水芹茎粗在U浓度一定时不随Cd浓度的增加表现显著差异(P<0.05)。与对照CK相比U、Cd复合处理下水芹根长在低浓度(U15 mg·kg-1+Cd15 mg·kg-1)处理下促进根伸长,是对照的1.11倍,当处理浓度高时显著抑制(P<0.05)。U、Cd单一和复合处理均可抑制水芹生长,且同元素条件下处理浓度越高其对水芹生长抑制作用越强,同浓度条件下不同元素复合胁迫效应大于单元素胁迫效应。
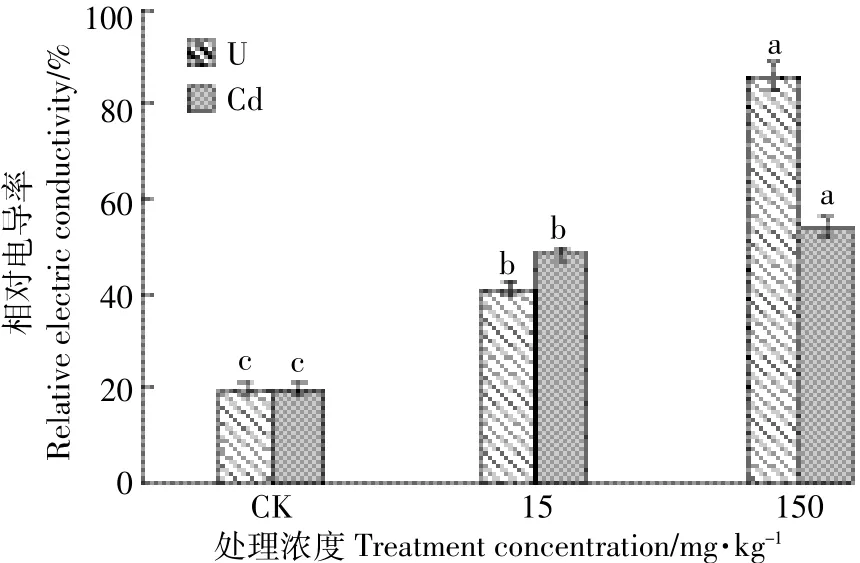
图1 U、Cd单一处理对水芹细胞膜透性的影响Figure 1 Effects of U and Cd single treatments on relative electrical conductivity of Oenanthe javanica
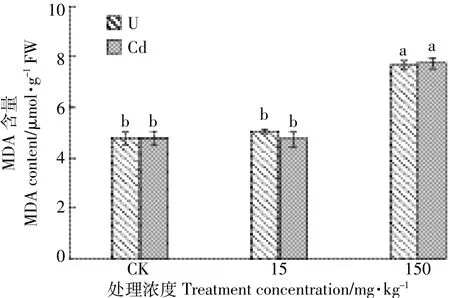
图2 U、Cd单一处理对水芹MDA含量的影响Figure 2 Effects of U and Cd single treatments on MDA content of Oenanthe javanica

表4 U、Cd复合处理对水芹生长的影响Table 4 Effects of U-Cd combinations on growth of Oenanthe javanica
2.2.2对水芹叶绿素含量和根系活力的影响
由表5分析得知,与对照CK相比,U、Cd复合处理下水芹叶绿素含量显著低于对照,可见复合处理对水芹叶绿素合成具有抑制作用,此结果与单元素处理效果一致。在低浓度U、Cd复合处理(U15 mg·kg-1+ Cd15 mg·kg-1,U15 mg·kg-1+Cd150 mg·kg-1)中,叶绿素b含量显著下降,且小于相应浓度下U、Cd单一处理,表现为拮抗效应,可见低浓度的U、Cd复合处理可显著抑制叶绿素b合成。在高浓度复合处理(U150 mg· kg-1+Cd15 mg·kg-1,U150 mg·kg-1+Cd150 mg·kg-1)中,U150 mg·kg-1+Cd15 mg·kg-1处理下叶绿素a、b含量略有升高,分别是单一U(U150 mg·kg-1)处理下的1.09 和1.16倍,可见一定浓度Cd元素的加入可缓解高浓度U对水芹叶绿素的破坏作用。水芹根系活力随复合处理浓度的增加呈下降趋势,低浓度复合处理(U15 mg·kg-1+Cd15 mg·kg-1)下水芹根系活力略高于对照CK,说明U、Cd复合处理浓度低时,水芹根尚可保持较好的根系活力,但小于同水平单元素处理下根系活力。在高浓度复合处理中(U150mg·kg-1+Cd15mg·kg-1,U150 mg·kg-1+Cd150 mg·kg-1),U、Cd复合胁迫的毒害效应小于同浓度下U、Cd单一胁迫且差异显著,说明在处理浓度高(150 mg·kg-1)时U、Cd复合处理对水芹根系活力影响比单一元素影响作用弱,主要原因可能是U、Cd复合污染对水芹根系活力表现拮抗作用。
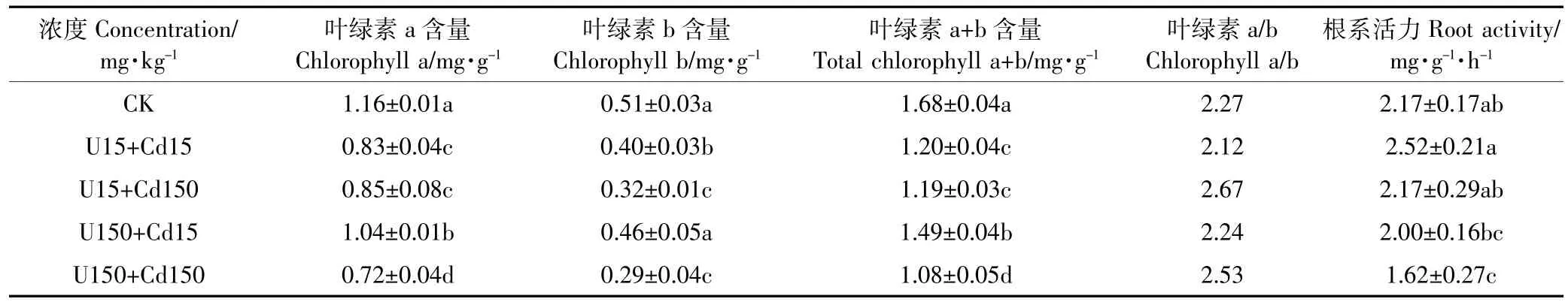
表5 U、Cd复合处理对水芹叶绿素含量和根系活力的影响Table 5 Effects of U-Cd combinations on chlorophyll content and root activity of Oenanthe javanica
2.2.3对水芹细胞膜透性和MDA含量的影响
比较U、Cd复合处理对水芹叶片相对电导率和MDA含量的影响,分析图3得知,复合处理浓度越高,水芹叶片相对电导率越大,细胞膜透性越强,但U、Cd复合处理组实验数据均低于相应浓度下的U、Cd单一处理。可以看出,水芹叶片细胞膜透性,U、Cd单一处理>U、Cd复合处理,更能使叶片膜脂过氧化,破坏膜系统,增加膜透性。U、Cd复合处理下水芹叶片MDA含量变化与水芹叶片相对电导率变化趋势一致(图4),U、Cd复合胁迫下水芹MDA含量介于单元素浓度15 mg·kg-1和150 mg·kg-1处理之间,说明水芹MDA含量在高浓度复合处理下,U、Cd间表现为拮抗作用而在低浓度复合处理下表现为协同作用。
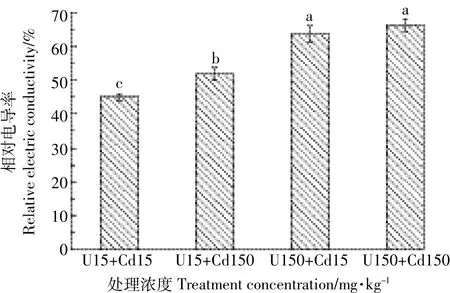
图3 U、Cd复合处理对水芹细胞膜透性的影响Figure 3 Effects of U-Cd combinations on relative electric conductivity of Oenanthe javanica
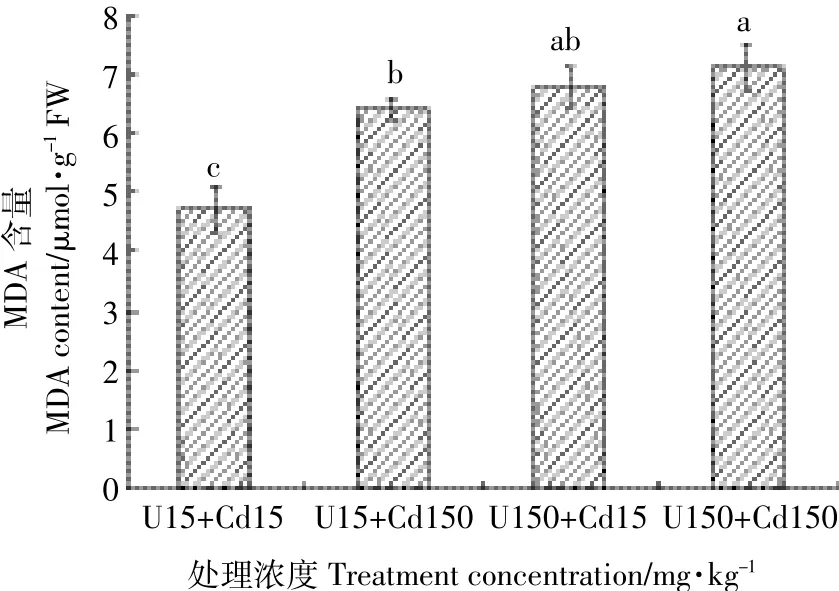
图4 U、Cd复合处理对水芹MDA含量的影响Figure 4 Effects of U-Cd combinations on MDA content of Oenanthe javanica
3 讨论
研究U、Cd单一及其复合污染处理下水芹生长状况,发现随U、Cd处理浓度的增加,水芹株高和茎粗均受到明显抑制作用,水芹的根长对U、Cd胁迫的响应表现为低浓度(15 mg·kg-1)促进高浓度(150 mg· kg-1)抑制现象。此现象在多数研究中都有报道[10-12]。其主要原因是低浓度的U、Cd可提高或加速根系生理生化反应从而促进根的伸长[13],然而当金属离子在植物根部过量积累时,会抑制根尖细胞伸长并加快细胞木质化进程,导致根长变短[14]。水芹对重金属胁迫的响应因重金属的种类、胁迫浓度、作用时间以及重金属复合处理下金属离子间相互作用效应而各有差异[15-16],试验中U元素对水芹生长胁迫效应大于Cd,且U、Cd复合处理下胁迫毒害效应大于相应浓度下单元素处理,复合处理中U、Cd元素间表现为协同效应。此结果与何芳芳等[17]研究结果一致。水芹生长状况对U、Cd胁迫响应差异的原因可能是Cd迁移性较强易于被植物吸收[18],而U分子量较大,不容易在植物体中转移,其单位面积积累浓度高,毒性强。
叶绿素是植物进行光合作用和机体正常代谢活动的重要保障,叶绿素含量的高低直接决定了光合作用的强弱及有机物合成的速率。试验中,水芹叶绿素含量随U、Cd单一及其复合处理浓度的增加而减小。张艳等[19]通过水培试验研究了不同浓度U、Cd复合胁迫对豌豆叶绿素含量的影响,结果表明豌豆对低浓度复合污染具有一定抗性,然而随U、Cd复合处理浓度的增加叶绿素含量表现下降趋势。根据Somashekaraish等[20]的研究,其可能原因是U、Cd在水芹体内大量积累,抑制了叶绿素酸酯还原酶和氨基-r-酮戊酸活性,使叶绿素的合成受阻,植物体叶绿素含量减少,光合速率降低,水芹有机质合成减少。这也印证了水芹在高浓度U、Cd胁迫时株高和茎粗降低的现象。试验中水芹叶绿素含量减少作用为Cd>U,有研究表明Cd可以抑制植物气孔开放,损害光合器官,特别是捕光色素蛋白复合体Ⅱ[21],抑制环式和非环式光合磷酸化[22],破坏叶绿体结构,影响植物光合作用[23]。
植物根系具有锚定植株,吸收和运输土壤中水分及养分,合成和贮藏营养物质等重要功能[24]。根系活力的强弱一定程度上反映植物对胁迫环境适应能力的大小。本研究中,水芹根系活力随处理浓度的增加,表现先升后降的趋势,在浓度15 mg·kg-1时根系活力最大,且U和Cd在同一处理浓度下水芹根系活力差异不显著(P<0.05),与U、Cd胁迫下水芹根长变化情况一致。有研究表明水芹能分泌丰富的脂溶性根系分泌物[4],因此在低浓度胁迫条件下水芹对U、Cd产生一定应激性,高浓度时水芹根系活力明显下降,可能原因是根部积累大量U、Cd元素直接影响了根系细胞的有丝分裂,使根系代谢紊乱,抑制了植物生长,降低根系活力。
正常情况下,植物细胞膜的选择透性对维持细胞正常代谢和微环境起着重要的作用。当植物受到逆境影响时,细胞膜的结构遭到破坏,膜透性增大,从而使细胞内的电解质外渗,导致植物细胞浸提液的电导率增大。试验结果表明U、Cd胁迫下水芹叶片相对电导率均高于对照,U、Cd可显著破坏细胞膜透性,并随处理浓度的增加而增强,与钟钼芝等研究的蚕豆结论一致[25]。U浓度150 mg·kg-1条件下水芹细胞膜透性最大,是同浓度Cd处理的1.62倍,可见U对细胞膜透性的影响显著强于Cd(P<0.05)。有研究表明铀对生物细胞具有很强的化学毒性[26],可导致植物体活性氧如氧氢自由基(HO2·)和超氧阴离子(O-2·)的增加[27],水芹抗氧化系统失衡,膜透性增大,细胞内电解质外渗液积累,水芹叶片相对电导率增大。
逆境胁迫条件下,植物体产生的大量活性氧,攻击细胞原生质膜中的不饱和脂肪酸双键而发生过氧化反应产生大量MDA,使植物细胞膜透性增大[28-29]。本研究中,U、Cd胁迫使水芹体内活性氧自由基积累,引发膜质过氧化作用产生大量MDA,MDA具细胞毒性,其含量在一定程度上反映植物的抗胁迫能力[30]。当U、Cd单一浓度为15 mg·kg-1时,水芹MDA含量与对照CK无显著差异(P<0.05),可见水芹对低浓度U、Cd胁迫具有一定抗性。随着处理浓度增加,水芹MDA含量不断增加,水芹抗胁迫能力逐渐降低,可能是水芹体内累积的铀、镉与酶蛋白中巯基(-SH)结合,抑制了抗氧化酶的合成,破坏了酶结构。
4 结论
U、Cd元素对水芹生长抑制效果一致,处理浓度越高胁迫效应越显著。但U、Cd元素对水芹生理胁迫效应因作用部位的不同而略有差异,两者对水芹根系活力的影响表现为拮抗作用,对MDA含量的影响表现为协同作用,Cd元素能显著抑制叶绿素b合成,U元素则对水芹叶片膜质过氧化作用更显著。此外,水芹在低浓度U、Cd单一及其复合污染处理下均能保持较好的抗逆性,因此水芹在修复低浓度铀镉污染土壤方面具有潜在应用价值。
参考文献:
[1]张斌,丁新民,段蕴铀.贫铀对机体健康影响的研究进展[J].中国预防医学杂志, 2011, 12(5):448-451. ZHANG Bin, DING Xin-min, DUAN Yun-you, et al. Research progress of the effects of depleted uranium on the health of the body[J]. Chin Prev Med, 2011, 12(5):448-451.
[2]曹心德,魏晓欣,代革联.土壤重金属复合污染及其化学钝化修复技术研究进展[J].环境工程学报, 2011, 5(7):1441-1453. CAO Xin-de, WEI Xiao-xin, DAI Ge-lian. Combined pollution of multiple heavy metals and their chemical immobilization in contaminated soils:A review[J]. Chinese Journal of Environmental Engineering, 2011, 5(7):1441-1453.
[3]Sun Y H, Li Z J, Guo B, et al. Arsenic mitigates cadmium toxicity in rice seedling[J]. Environmental and Botany, 2008, 64(3):264-270.
[4]吉敏,孟梁.芹菜对土壤中多环芳烃修复作用的初步研究[J].环境科技, 2014, 27(5):12-15. JI Min, MENG Liang. Remediation of PAHs-contaminated soil by watercress[J]. Environmental Science and Technology, 2014, 27(5):12-15.
[5]张迪,胡学玉,殷俊,等.城郊农业生态系统中茎菜类蔬菜对重金属镉的累积特征及品质表现[J].应用与环境生物学报, 2015, 21 (2):188-194. ZHANG Di, HU Xue-yu, YIN Jun, et al. Cadmium accumulation and vegetable quality of edible parts of stem vegetables growing in suburban farmlands[J]. Chin JAppl Environ Biol, 2015, 21(2):188-194.
[6]张志良,瞿伟菁,李小方.植物生理学实验指导[M].北京:高等教育出版社, 2009:54-55. ZHANG Zhi-liang, QU Wei-jing, LI Xiao-fang. Plant physiology experiment[M]. Beijing:Higher Education Press, 2009:54-55.
[7]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社, 2003:261-263. LI He-sheng. Experimental principle and technology of plant physiology and biochemistry[M]. Beijing:Higher Education Press, 2003:261-263.[8]王学奎.植物生理生化试验原理与技术[M].北京:高等教育出版社, 2006:169-171. WANG Xue-kui. The principle and technology of plant physiological and biochemical experiment[M]. Beijing:Higher Education Press, 2006:169-171.
[9]Brown M T, Wilkins D A. The effects of zinc on germination, survival and growth of Betula seed[J]. Ecological and Biological, 1986, 1(41):53-61.
[10]王亭亭,王中浩,熊方建,等.铀胁迫对油菜的生长及抗氧化酶的影响[J].四川大学学报(自然科学版), 2014, 51(1):171-176. WANG Ting-ting, WANG Zhong-hao, XIONG Fang-jian, et al. Uranium induced effects on development and antioxidase of Brassica napus L.[J]. Journal of Sichuan University(Natural Science Edition), 2014, 51(1):171-176.
[11]荣丽杉,杨金辉,凌辉,等.印度芥菜对铀的生理响应与积累特征[J].金属矿山, 2015, 469(7):155-158. RONG Li-shan, YANG Jin-hui, LING Hui, et al. Physiological response and accumulation character of Brassica juncea under uranium stress[J]. Metal Mine, 2015, 469(7):155-158.
[12]聂小琴,丁德馨,董发勤,等.水培和盆栽条件下碎米莎草对铀的耐受性及富集特征[J].环境科学研究, 2015, 28(5):831-838. NIE Xiao-qin, DING De-xin, DONG Fa-qin, et al. Effects of tolerance and enrichment characteristics of Cyperus iria L. to uranium under hydroponic and potted culture[J]. Research of Environmental Sciences, 2015, 28(5):831-838.
[13]蒋汉明,李书启,韩希凤,等.镉对植物生长的影响及植物耐镉机理研究进展[J].广东微量元素科学, 2012, 19(5):1-6. JIANG Han-ming, LI Shu-qi, HAN Xi-feng, et al. Progresses on the effect of cadmium for plant and their tolerant mechanism[J]. Guangdong Weiliang Yuansu Kexue, 2012, 19(5):1-6.
[14]Aline F T, Maria D F, Anderson R S, et al. Cadmium-induced lignification restricts soybean root growth[J]. Ecotoxicology and Environmental Safety, 2010, 73(8):1959-1964.
[15]Sarwar N, Ishaq W, Farid G, et al. Zinc-cadmium interactions:Impact on wheat physiology and mineral acquisition[J]. Ecotoxicology and Environmental Safety, 2015, 122(10):528-536.
[16]肖厚军,芶久兰,何佳芳,等.镉胁迫下芹菜产量和氮、磷、钾吸收及镉积累量的变化[J].环境科技, 2014, 27(4):10-13. XIAO Hou-jun, GOU Jiu-lan, HE Jia-fang, et al. Effect of cadmium stress on yield, nitrogen, phosphorus, potassium and cadmium uptake of Apium graveolens[J]. Environmental Science and Technology, 2014, 27(4):10-13.
[17]何芳芳,张德刚,陈雅顺,等.重金属复合污染对芹菜生长的影响[J].北方园艺, 2012, 20(20):5-7. HE Fang-fang, ZHANG De-gang, CHEN Ya-shun, et al. Effect of combined heavy metals pollution on the growth of celery[J]. Northern Horticulture, 2012, 20(20):5-7.
[18]曹霞,郭朝晖,肖细元,等.海桐(Pittosporum tobira)对污染土壤中镉的耐受和吸收特征[J].农业环境科学学报, 2015, 34(4):627-632. CAO Xia, GUO Zhao-hui, XIAO Xi-yuan, et al. Cadmium tolerance and uptake characteristics in Pittosporum tobira in contaminated soil [J]. Journal of Agro-Environment Science, 2015, 34(4):627-632.
[19]张艳,王丹,李黎,等.豌豆对U和Cd复合胁迫反应及其积累特征[J].辐射研究与辐射工艺学报, 2013, 31(1):55-59. ZHANG Yan, WANG Dan, LI Li, et al. Response of peas to U and Cd combined stress and their accumulation characteristics[J]. J Radiat Res Radiat Process, 2013, 31(1):55-59.
[20]Somashekaraiah B V, Padmaja K, Prasad A R K, et al. Phytoxicity of cadmium ions on germination seedling of mung bean:Involvement of lipid peroxides in chlorophyll degradation[J]. Plant Physiology, 1992, 85:85-89.
[21]Li X, Zhao M X, Guo L P, et al. Effect of cadmium on photosynthetic pigments, lipid peroxidation, antioxidants, and artemisinin in hydroponically grown Artemisiaannua[J]. Journal of Environmental Sciences, 2012, 24(8):1511-1518.
[22]Vanalakar S A, Mali S S, Pawar R C, et al. Synthesis of cadmium sulfide spongy balls with nanoconduits for effective light harvesting[J]. Electrochimica Acta, 2011, 56(6):2762-2768.
[23]杨菲,唐明凤,朱玉兴,等.水稻对镉的吸收和转运的分子机理[J].杂交水稻, 2015, 30(3):2-8. YANG Fei, TANG Ming-feng, ZHU Yu-xing, et al. Molecular mechanism of cadmium absorption and transport in rice[J]. Hybrid Rice, 2015, 30(3):2-8.
[24]马献发,宋凤,张继舟,等.根系对土壤环境胁迫响应的研究进展[J].中国农学通报, 2011, 27(5):44-48. MA Xian-fa, SONG Feng, ZHANG Ji-zhou, et al. Advances of research of roots responses to environmental stress on soil[J]. Chinese Agricultural Science Bulletin, 2011, 27(5):44-48.
[25]钟钼芝,王丹,徐长合,等.螯合剂对铀镉污染土壤中蚕豆幼苗生理特性影响[J].农业环境科学学报, 2011, 30(4):639-644. ZHONG Mu-zhi, WANG Dan, XU Chang-he, et al. The physiological characteristics of Vicia faba Linn. to extract uranium cadmium under chelate treatment[J]. Journal of Agro-Environment Science, 2011, 30 (4):639-644.
[26]Sheppard S C, Sheppard M I, Gallerand M O, et al. Derivation of ecotoxicity thresholds for uranium[J]. Journal of Environmental Radioactivity, 2005, 79(1):55-83.
[27]Srivastavas S, Bhainsa K C, D’Souza S F. Investigation of uranium accumulation potential and biochemical responses of an aquatic weed Hydrilla verticillata(L.f.)Royle[J]. Bioresource Technology, 2010, 101 (8):2573-2579.
[28]Wu L H, Yi H L, Li M. Assessment of arsenic toxicity using allium/vicia root tip micronucleus assays[J]. Journal of Hazardous Materials, 2010, 176(1):952-956.
[29]惠俊爱,党志,叶庆生.镉胁迫对玉米光合特性影响[J].农业环境科学学报, 2010, 29(2):205-210. HUI Jun-ai, DANG Zhi, YE Qing-sheng. Influence of cadmium stress on photosynthetic characteristics of maize[J]. Journal of Agro-Environment Science, 2010, 29(2):205-210.
[30]王贺正,马均,李旭毅,等.水分胁迫对水稻结实期一些生理性状的影响[J].作物学报,2006,32(12):1892-1897. Wang H Z, Ma J, Li X Y, et al. Effects of water stress on some physiological characteristics in rice during grain filling stage[J]. Acta Agronomica Sinica, 2006, 32(12):892-1897.
中图分类号:X591
文献标志码:A
文章编号:1672-2043(2016)05-0871-07
doi:10.11654/jaes.2016.05.008
收稿日期:2015-11-02
基金项目:国防科工局国家核设施退役及放射性废物治理科研重点项目(14ZG6101)
作者简介:姚天月(1990—),女,新疆奎屯人,硕士研究生,从事辐射生物效应及其生物修复研究。E-mail:531801450@qq.com
*通信作者:王丹E-mail:wangdan@swust.edu.cn
Effects of single and combined pollution of uranium and cadmium on growth and physiological characteristics of Oenanthe javanica
YAO Tian-yue1,2, WANG Dan1,2*, LI Ze-hua1,2
(1.Life Science and Engineering College, Southwest University of Science and Technology, Mianyang 621000, China; 2.State Defense Key Laboratory of the Nuclear Waste and Environmental Security, Southwest University of Science and Technology, Mianyang 621000, China)
Abstract:The growth, chlorophyll content, root activity, cell membrane permeability and MDA content of Oenanthe javanica(Blume)DC were studied under uranium(U)and cadmium(Cd)alone and in combination in pot experiment. Results showed that U and Cd stresses significantly inhibited the growth of O. javanica(P<0.05)when compared with the control. The inhibition increased with increasing concentrations of U and Cd. The chlorophyll content of O. javanica decreased remarkably under U and Cd stresses. Cadmium reduced the content of chlorophyll b of O. javanica more significantly than U did. The activity of O. javanica roots was enhanced by low concentrations(15 mg·kg-1)of U and Cd, but significantly inhibited at high concentrations(150 mg·kg-1). The relative electrical conductivities of leaves were higher under U and Cd stresses than under CK, with U showing greater effect than Cd. Leaf MDA content increased as U and Cd concentrations increased. Combined applications of U and Cd(U150 mg·kg-1+Cd 150 mg·kg-1)had greater influences on O. javanica’s growth and physiological characteristics than their single. The present findings showed that O javanica has high resistance to U and Cd, which has potential to remediate U and Cd single and combined pollution soils.
Keywords:Oenanthe javanica; uranium; cadmium; physiological characteristic
