盐胁迫下苋菜品种有机酸变化对Cd累积和耐盐性的影响
2016-06-20陈柯罕李取生徐智敏郭世鸿余丹萍暨南大学环境学院广州市环境暴露与健康重点实验室广州510632广东省环境污染控制与修复材料中心广州510632暨南大学生命科学技术学院广州510632
胡 妮,陈柯罕,李取生*,徐智敏,3,郭世鸿,3,余丹萍,罗 涛(1.暨南大学环境学院,广州市环境暴露与健康重点实验室,广州510632;2.广东省环境污染控制与修复材料中心,广州510632;3.暨南大学生命科学技术学院,广州510632)
盐胁迫下苋菜品种有机酸变化对Cd累积和耐盐性的影响
胡妮1,2,陈柯罕1,2,李取生1,2*,徐智敏1,2,3,郭世鸿1,2,3,余丹萍1,2,罗涛1,2
(1.暨南大学环境学院,广州市环境暴露与健康重点实验室,广州510632;2.广东省环境污染控制与修复材料中心,广州510632;3.暨南大学生命科学技术学院,广州510632)
摘要:采用盆栽试验,研究0.3%NaCl胁迫下全红苋菜(低Cd累积品种,简称全红)和柳叶苋菜(高Cd累积品种,简称柳叶)的有机酸代谢差异及其与Cd累积和耐盐性的关系。结果表明:相比对照,盐胁迫后两个品种根部、地上部和根际土壤溶液中的Cd含量均呈增加趋势,且表现为全红显著低于柳叶(P<0.05),同时全红和柳叶的Cd转运系数分别提高2.84、5.58倍;盐胁迫促进了苋菜对Na+的积累且明显干扰K+的吸收,K+/Na+均显著降低(P<0.05),且全红根部K+/Na+显著高于柳叶(P<0.05)。盐胁迫后,两个品种根部、地上部及根际土壤溶液中均检测到多种有机酸,其中根际土壤溶液中苹果酸和草酸含量均显著升高(P<0.05),且与根际土壤溶液中的Cd含量显著正相关;柳叶地上部草酸及根部乙酸的含量均显著升高(P<0.05),且分别与地上部及根部Cd含量显著正相关。研究表明,盐分胁迫下,全红具有较低的Cd累积性且表现出更强的耐盐性,不同品种的有机酸积累和分泌的差异是导致这一结果的重要原因之一。
关键词:盐分胁迫;苋菜;有机酸;Cd累积;耐盐性
胡妮,陈柯罕,李取生,等.盐胁迫下苋菜品种有机酸变化对Cd累积和耐盐性的影响[J].农业环境科学学报, 2016, 35(5):858-864.
HU Ni, CHEN Ke-han, LI Qu-sheng, et al. Effects of salinity-inducted organic acid variation on Cd accumulation and salinity tolerance of edible amaranth[J]. Journal of Agro-Environment Science, 2016, 35(5):858-864.
土壤盐渍化已成为全球瞩目的问题。盐渍土壤占我国耕地总面积的近1/3,我国盐渍化的土壤区域主要集中在北方及沿海地区;据统计,我国的盐渍化耕地面积达到760万hm2,严重威胁到我国的可利用耕地面积[1]。土壤中过高的盐分会引起土壤的理化性质发生改变,抑制植物的生长,甚至引起死亡[2],严重影响现代农业的发展。沿海地区的滩涂资源分布广泛,且主要应用于农业。随着沿海地区经济的发展和城市化进程的加快,污染物的不合理排放导致滩涂中重金属严重超标。盐分和重金属的双重胁迫直接影响了植物对水分的利用和生物量,对农业生产产生严重影响[3]。我国海岸线漫长,且滩涂面积广泛,滩涂土壤的含盐量为0.15%~0.4%,滩涂成为沿海地区重要的土地后备资源,对其开发和利用已经成为农业发展的必然趋势和发展农业经济的基础[4-5]。
有机酸是一类具有一个或多个羧基的低分子化合物,是植物体内重要的代谢产物,对于植物应对各种胁迫起着重要作用[6]。逆境胁迫下,植物通过分泌和累积有机酸,调节根际环境和体内的代谢活动,这是植物对环境的适应性响应。有文献报道[7-8],盐分通过影响某些酶的活性,从而影响有机酸的分泌,以减轻盐分对植物的毒害作用。另一方面,在植物体内,有机酸在重金属的吸收、累积和解毒过程中起着重要作用[9-10]。植物体内的有机酸与重金属络合,降低了重金属的生物有效性和对植物酶系统的破坏,从而减轻重金属离子的毒害作用[11]。因此,探讨盐胁迫下植物分泌和积累的有机酸与重金属的关系,有助于了解植物对受重金属污染的盐渍土的耐受机制。
本试验所选用的苋菜具有较高的营养价值和一定的药用价值,在我国南方沿海滩涂围垦农田大量种植。本文基于课题组前期筛选的低Cd累积品种全红苋菜和高Cd累积品种柳叶苋菜[12],研究了两种苋菜有机酸代谢差异及其与耐盐性和Cd累积的关系。
1 材料与方法
1.1试验材料
本试验所用苋菜种子购买于市场。供试土壤采自广州郊区菜园,土壤基本理化性质:pH为6.3,总有机碳为35.4 g,阳离子交换量为20.86 cmol·kg-1,Cd含量2.28 mg·kg-1,K+含量19.1 g·kg-1,Na+含量169.6 g· kg-1。根据国家土壤环境质量标准(GB 15618—1995)二级标准,供试土壤Cd超标,属于重金属污染土壤。
1.2盆栽试验
设置对照(CK)和0.3%盐分处理(T1,每千克风干土壤中添加3 g NaCl)。土壤经晒干,研磨,混匀,准确称取500 g干土于300目尼龙根际袋中,根际袋直径约为10 cm,每个塑料花盆中均匀放置3个根际袋,以袋中的土作为植株的根际土,用1000 g干土填充花盆与根际袋的空隙,塑料花盆中干土总量为2500 g。种子经消毒,直接播种在根际袋中,待苋菜幼苗长至5片叶时间苗,每个根际袋中留苗3株。两个苋菜品种,对照和处理各3个平行,共12盆。盆栽试验时间为7—8月,环境温度为30~35℃,土壤持水量约为80%。
1.3样品处理及测定方法
苋菜种植50 d后收割,用去离子水洗净,后用滤纸吸干水分,分离地上部和根部。一部分鲜样加去离子水研磨呈匀浆,9000 r·min-1离心30 min,收集上清液经0.45 μm滤膜过滤后,使用戴安ICS900离子色谱仪测定有机酸[13]。一部分地上部样品放入烘箱,105℃杀青20 min,后置于60℃烘箱烘至恒重,样品干燥后磨碎,准确称取0.2 g干样,混合10 mL浓硝酸,电热板消解,消解过程中添加植物标准样品(GSV-1国家标准参比物质),保证测定元素在允许的误差范围内进行测试以达到质控要求。使用岛津AA-7000原子吸收分光光度计测定Na+、K+、Cd的含量;根部鲜样用(NH4)2EDTA溶液浸泡并超声处理后提取细胞质外体的金属[14],经(NH4)2EDTA提取后的根部置于烘箱中,60℃烘至恒重后磨碎并消解,用于测定Na+、K+、Cd的含量。用抖根法收集根际袋中的根际土,高速离心机中离心,获得苋菜根际土壤溶液,经0.45 μm滤膜过滤后测定Cd的含量。
1.4数据处理
采用SPSS软件对数据进行统计学分析,Origin软件作图。所有数据采用3次重复试验的平均值±标准差。
2 结果分析
2.1苋菜、根际土壤溶液Cd含量
两苋菜品种根部、地上部及根际土壤溶液Cd含量如表1所示,根向地上部的转运系数如图1所示。受到盐胁迫后,两苋菜品种的根部和地上部的Cd含量均增加,其中全红地上部的Cd含量显著增加,柳叶地上部和根部的Cd含量都显著增加,柳叶根部和地上部的Cd含量分别是全红的1.85、3.25倍。盐胁迫使得全红和柳叶的Cd转运系数分别从0.79和0.93提高到2.24和5.19,增加幅度分别为184.54%和458.06%。由表1可知,加盐处理后,两苋菜品种根际土壤溶液中Cd含量均显著升高,且柳叶根际土壤溶液的Cd含量是全红的1.90倍。
两苋菜品种根部、地上部和根际土壤溶液中Cd含量的相关性表明(表2),全红根际土壤溶液中Cd含量与地上部Cd含量呈极显著正相关,柳叶根际土壤溶液及地上部和根部Cd含量均呈极显著正相关。
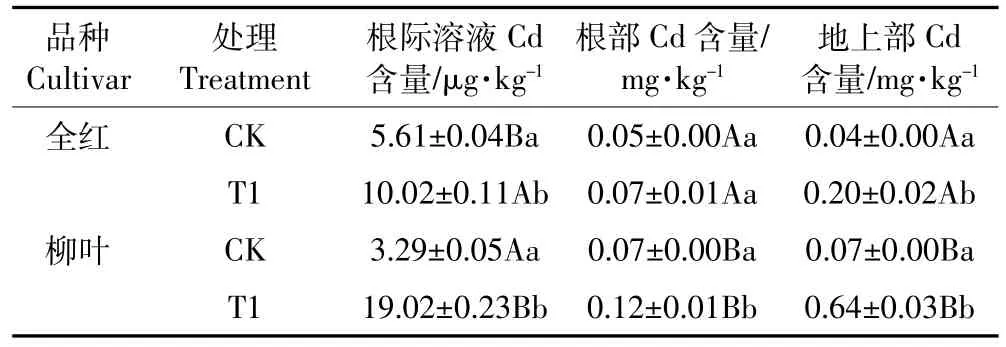
表1 苋菜及其根际土壤溶液中Cd含量Table 1 Content of Cd in amaranth and its rhizospheric soil solution

图1 苋菜根部向地上部Cd转运系数Figure 1 Translocation ratios of Cd from roots to shoots
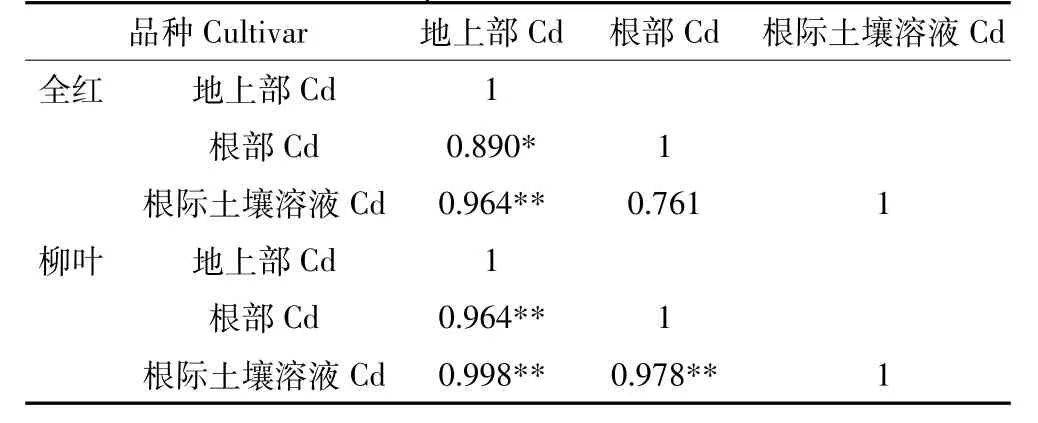
表2 苋菜植株与根际土壤溶液Cd含量相关性Table 2 Correlation coefficients between Cd in amaranth and in rhizospheric soil solution
2.2苋菜对K+、Na+的吸收
如图2所示,盐胁迫下两苋菜品种根部和地上部的Na+含量都显著性升高,且全红显著低于柳叶,全红和柳叶根部Na+含量分别升高了3.21、3.63倍,地上部Na+含量分别升高了1.34、2.64倍。另外,盐胁迫下两苋菜品种根部截留Na+的能力增强,使根部Na+含量均高于地上部。全红地上部的K+含量显著下降,而柳叶地上部K+含量虽下降但变化不显著;两苋菜品种根部的K+含量均显著性升高,全红和柳叶K+含量分别上升了2.73、2.20倍(图3)。盐胁迫下两苋菜品种根部和地上部的K+/Na+值均显著下降(图4),其中全红根部和地上部的下降幅度分别为15.17%和56.23%,柳叶根部和地上部的下降幅度分别为40.26%和62.76%,全红根部的K+/Na+显著高于柳叶,为柳叶的2.44倍。
2.3盐胁迫下苋菜有机酸的积累和分泌
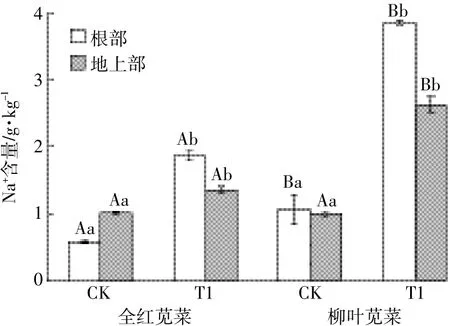
图2 苋菜体内Na+含量Figure 2 Na+content in amaranth
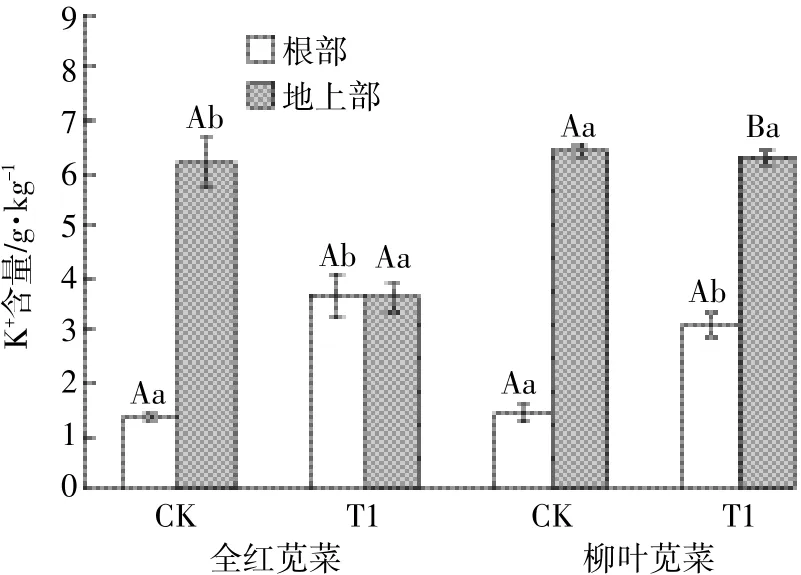
图3 苋菜体内K+含量Figure 3 K+content in amaranth
如表3所示,两苋菜品种的根际土壤溶液中检测出乳酸、乙酸、甲酸、苹果酸、草酸。盐胁迫下,全红根际土壤溶液中的苹果酸和草酸含量显著增加,而乳酸含量显著降低。柳叶根际土壤溶液的乳酸、苹果酸含量显著增加。盐胁迫下全红根际土壤溶液的乳酸、甲酸含量显著高于柳叶,分别是柳叶的1.13、2.67倍。而柳叶根际土壤溶液的乙酸、草酸显著高于全红,分别是全红的1.31、2.10倍。除了根际土壤溶液中检测出来的五种低分子有机酸外,两苋菜品种的根部和地上部均检测到酒石酸。盐胁迫下全红根部甲酸、苹果酸含量显著下降,而草酸含量显著升高,地上部乳酸、乙酸、苹果酸含量均显著下降。柳叶根部甲酸、苹果酸、酒石酸含量显著下降,而乙酸含量显著上升,地上部乳酸、甲酸和酒石酸显著降低,苹果酸和草酸显著升高。盐胁迫下,柳叶根部乳酸、乙酸、草酸含量显著高于全红,分别是全红的5.07、2.23、1.12倍,而酒石酸含量显著低于全红。柳叶地上部乙酸、苹果酸、草酸含量分别是全红的2.50、4.93、1.18倍。
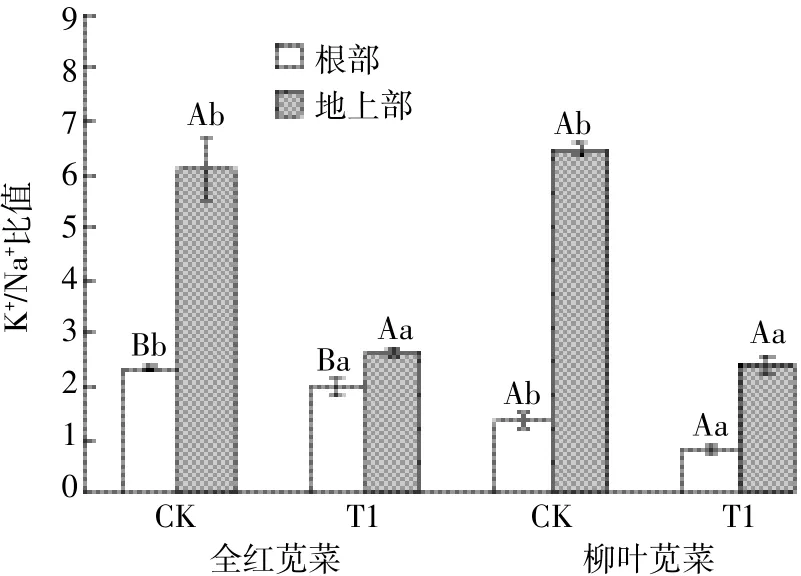
图4 苋菜体内K+/Na+比值Figure 4 K+/Na+ratios in amaranth
盐胁迫下,苋菜地上部、根部和根际土壤溶液中Cd含量与有机酸的相关性如表4所示。全红根际土壤溶液中的乳酸与Cd含量极显著负相关,苹果酸、草酸含量与Cd含量极显著正相关;地上部乳酸含量与Cd含量极显著负相关,乙酸、苹果酸、草酸含量与Cd含量显著负相关。柳叶根际土壤溶液中乳酸、苹果酸、草酸与Cd含量极显著正相关,甲酸含量与Cd含量极显著负相关;根部乙酸含量与Cd含量极显著正相关,甲酸、苹果酸、酒石酸含量与Cd含量极显著负相关;地上部乳酸、甲酸含量与Cd含量显著负相关,酒石酸含量与Cd含量极显著负相关,草酸含量与Cd含量极显著正相关。可见,盐分确实导致了苋菜某些有机酸分泌和积累的显著性变化,且不同苋菜品种的变化存在差异,并且有机酸分泌和积累对Cd的吸收和累积产生一定的影响。
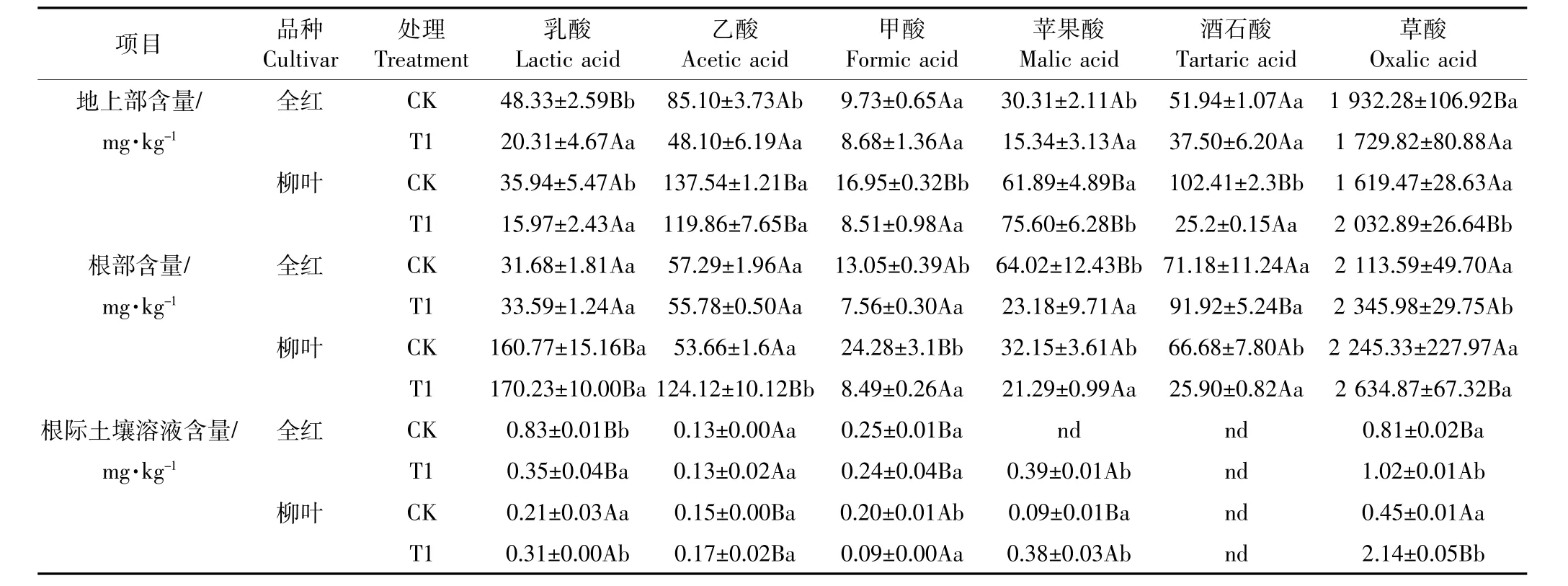
表3 苋菜及其根际土壤溶液中有机酸含量Table 3 Content of organic acids in amaranth and rhizospheric soil solution
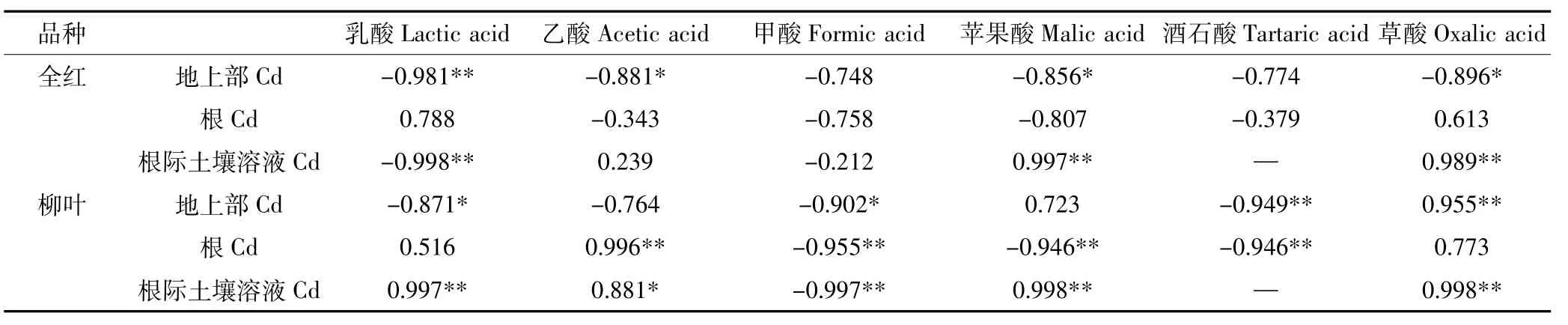
表4 苋菜、根际土壤溶液中Cd含量与有机酸含量相关性Table 4 Correlation coefficients between Cd content and organic acid content in amaranth and rhizospheric soil solution
3 讨论
本试验测定结果表明,两苋菜品种根部、地上部及根际土壤溶液中的Cd含量均增加。同时两个品种的Cd转运系数都升高,全红和柳叶的Cd转运系数分别提高2.84倍和5.58倍。转运系数反应植物根部向地上部转运重金属的能力,转运系数越大,说明植物转运重金属的能力越强。这说明盐分促进了两苋菜品种对Cd的吸收、累积及转运。徐智敏等[15]研究发现,随着土壤盐分浓度的增加,苋菜品种根、叶中的Cd含量均显著增加。许多研究表明,土壤中的Na+与Cd争夺土壤吸附位点或Cl-通过复杂的化学反应与Cd形成络合物等方式,提高Cd的可迁移性和生物有效性,进而促进植物的吸收[16-17]。这也是本试验中苋菜对Cd吸收和累积量增加的重要原因。本试验中,盐胁迫下柳叶根部和地上部的Cd含量分别是全红的1.85、3.25倍,且柳叶根际土壤溶液的Cd含量是全红的1.90倍,说明在相同浓度的盐胁迫下,两苋菜品种对Cd的吸收和累积能力存在差异。有研究发现,植物可以通过自身分泌和累积有机物,以及对无机离子的吸收改变根细胞的结构和功能特性[18],盐胁迫下,不同苋菜品种的响应差异是造成其对Cd吸收和累积差异的重要原因。
本试验中,盐胁迫下两苋菜品种根际土壤溶液中的苹果酸和草酸含量升高,说明盐分促进了苋菜根系分泌苹果酸和草酸。植物根系分泌的有机酸酸化土壤根际环境,与重金属形成络合物,使土壤溶液中的重金属离子降低,破坏了土壤液相和固相中重金属离子的平衡,重金属离子从土壤粘粒表面解吸,提高了土壤中重金属的可溶性和移动性,促进了植物对重金属的吸收[19]。Nigam等[20]研究发现,随着向土壤中添加有机酸的量的增加,有机酸与土壤中的Cd形成络合物,玉米对镉的吸收量增加。柳叶根际土壤溶液中草酸含量高于全红,与盐胁迫下柳叶根部、地上部和根际土壤溶液中的Cd含量显著高于全红是一致的。
有机酸在植物对重金属的转运和解毒方面起着重要作用。土壤中的重金属以离子形式或者有机酸-重金属络合态的形式进入植物根系,贮存在植物根部或者运输到地上部。有研究发现,植物体内游离的重金属比络合态的重金属对植物的毒害作用更强[21]。从两种苋菜地上部和根部积累的有机酸与Cd的相关性可知,苋菜体内有机酸的含量和种类与重金属的累积和解毒有关。本试验中,柳叶地上部草酸含量和根部乙酸含量显著升高,且与Cd含量显著正相关,说明柳叶地上部草酸和根部乙酸在重金属转运和解毒方面起着一定的作用。Ma等[22]通过对木质部汁液中铝的形态进行分析发现,木质部中的铝主要是以铝-柠檬酸络合态的形式存在,说明铝在木质部主要是以络合态的形式存在。Roser等[23]发现Zn超累积植物T. Cae rulescens地上部分的苹果酸和草酸与Zn形成复杂的络合物,以降低重金属的毒性。不同苋菜品种积累的有机酸与Cd的相关性不同,可能是不同苋菜品种积累的有机酸存在差异,且不同种类的有机酸对Cd的解毒和转运能力不同造成的。
离子区隔化是植物耐盐及维持植物体内离子平衡的重要途径。由Na+、K+的含量测定可知,两种苋菜根部Na+含量均高于地上部,刘爱荣等[24]研究发现,彩叶草主要将Na+截留在根部,其次是地上部,以减少对叶片的毒害作用。植物将Na+截留在根部的液泡中,减少向地上部分的转移,减轻对植物的毒害作用。K+是植物生长发育必需的矿质元素,K+和Na+具有相似的原子结构,植物对Na+和K+的吸收有明显的拮抗作用,因此常以K+/Na+来衡量植物耐盐性[25]。本试验中,两苋菜品种受到盐分的胁迫,地上部和根部的K+/ Na+比值均下降,全红的根部K+/Na+显著高于柳叶根部。丁能飞等[26]研究了盐胁迫下两个品种大白菜体内的离子分布,发现两个品种大白菜的K+/Na+均下降,耐盐性较强的大白菜品种K+/Na+高。这说明全红比柳叶有较强的耐盐性。有机酸是植物体内重要的代谢物质和根系分泌物,影响着植物体内外的各种生理活动。在逆境胁迫下,植物通过一系列的生理变化,调节体内代谢及根系分泌有机酸的数量和种类做出响应。本试验中两个品种通过不同的有机酸积累机制应对盐胁迫,全红根部草酸含量显著增加,柳叶根部草酸含量无显著变化。郭立泉等[27]研究发现,在盐胁迫下,星星草体内的草酸含量显著增加以应对盐渍胁迫。因此,全红根部较高的K+/Na+与草酸含量的显著增加有关,全红叶部较低的Na+积累与根部的截留作用密切相关,而根部草酸含量的显著增加在一定程度上缓解了盐离子对细胞的毒害作用。
4 结论
(1)盐胁迫促使苋菜根系分泌苹果酸和草酸,且此二者含量与苋菜根等器官中的Cd含量极显著正相关(P<0.01)。
(2)盐胁迫下,全红根际土壤溶液中草酸含量显著低于柳叶,这是全红具有较低Cd累积性的重要原因。
(3)盐胁迫下,全红的根部K+/Na+显著高于柳叶,全红具有较强的耐盐性。
参考文献:
[1]王遵亲.中国盐渍土[M].北京:科学出版社, 1993:1-6. WANG Zun-qin. Saline soil of China[M]. Beijing:The Science Publishing Company, 1993:1-6.
[2]罗旋,陈小娇,李取生,等.盐分胁迫下番茄盐分离子和重金属的分布特征[J].农业环境科学学报, 2012, 31(4):654-660. LUO Xuan, CHEN Xiao-jiao, LI Qu-sheng, et al. Distribution characteristics of salt ions and heavy metal in tomato under salinity stress[J]. Journal of Agro-Environment Science, 2012, 31(4):654-660.
[3]Zhang F Q, Wang Y S, Lou Z P, et al. Effect of heavy metal stress on antioxidative enzymes and lipid peroxidation in leaves and roots of two mangrove plant seedlings(Kandelia candel and Bruguiera gymnorrhiza)[J]. Chemospher, 2007, 67(1):44-50.
[4]李取生,楚蓓,石雷,等.珠江口滩涂湿地土壤重金属分布及其对围垦的影响[J].农业环境科学学报, 2007, 26(4):1422-1426. LI Qu-sheng, CHU Bei, SHI Lei, et al. Heavy metal distribution in tidal wetland soils and its effect on reclamation in the Pearl River Estuary[J]. Journal of Agro-Environment Science, 2007, 26(4):1422-1426.
[5]刘芳文,颜文,王文质,等.珠江口沉积物重金属污染及其潜在生态危害评价[J].海洋环境科学, 2008, 21(3):31-38. LIU Fang-wen, YAN Wen, WANG Wen-zhi, et al. Pollution of heavy metals in the Pearl River Estuary and its assessment of potential ecological risk[J]. Marine Environmental Science, 2008, 21(3):31-38.
[6]Huang W B, Ma R, Yang D, et al. Organic acids secreted from plant roots under soil stress and their effects on ecological adaptability of plants[J]. Agricultural Science & Technology, 2014, 15(7):1167-1173.
[7]Watanabe T, Misawa S, Osaki M. Aluminum accumulation in the roots of Melastoma malabathricum, all aluminum-accumulating plant[J]. Canadian Journal of Botany, 2005, 83(11):1518-1522.
[8]Shen R F, Iwashita T, Ma J E. Form of Al changes with Al concentration in leaves of buckwheat[J]. Journal of Experimental Botany, 2004, 55 (394):131-136.
[9]Boominathan R, Doran P M. Organic acid complexation, heavy metal distribution and the effect of ATPase inhibition in hairy roots of hyper accumulator plant species[J]. Journal of Biotechnology, 2003, 101(2):131-146.
[10]Zhao F J, Lombi E, Breedon T, et al. Zinc hyperaccumulation and cellular distribution in Arabidopsis halleri[J]. Plant, Cell & Environment, 2000, 23(5):507-514.
[11]朱艳霞,魏幼璋,叶正钱,等.有机酸在超积累植物重金属解毒机制中的作用[J].西北农林科技大学学报(自然科学版), 2006, 34(7):121-126. ZHU Yan-xia, WEI You-zhang, YE Zheng-qian, et al. Construction of mathematic model for solar greenhouse cucumber balanced fertilization in the sandy land[J]. Journal of Northwest Sci-Tech University of Agriculture and Forestry(Natural Science Edition), 2006, 34(7):121-126.
[12]陈艳.不同苋菜品种对重金属累积差异和机制初探[D].广州:暨南大学, 2013:12-13. CHEN Yan. The differences in accumulation of heavy metals in amaranth cultivars and preliminary mechanisms research[D]. Guangzhou:Jinan University, 2013:12-13.
[13]沈宏,严小龙,郑少玲,等.菜豆有机酸和无机阴离子同时分析的离子色谱法[J].华南农业大学学报, 2001, 22(2):8-10. SHEN Hong, YAN Xiao-long, ZHENG Shao-ling, et al. Simultaneous analysis of organic acids and inorganic anions in common bean by ion chromatography[J]. Journal of South China Agricultural University, 2001, 22(2):8-10.
[14]Feng M H, Shan X Q, Zhang S Z, et al. A comparison of the rhizosphere-based method with DTPA, ECTE, CaCl2, and NaNO3extraction methods for prediction of metal in soil to barley[J]. Environmental Pollution, 2005, 137(2):231-240.
[15]徐智敏,何宝燕,李取生,等.盐分胁迫下两个苋菜品种对镉及主要渗透调节物质累积的差异[J].生态学杂志, 2015, 34(2):483-490. XU Zhi-min, HE Bao-yan, LI Qu-sheng, et al. Differences between two amaranth cultivars in accumulations of Cd and main osmotic adjustment substances under salt stress[J]. Chinese Journal of Ecology, 2015, 34(2):483-490.
[16]Acosta J A, Jansen B, Kalbitz K, et al. Salinity increases mobility of heavy metals in soils[J]. Chemosphere, 2011, 85(8):1318-1324.
[17]Fitzgerald E, Caffrey J, Nesaratnam S, et al. Copper and lead concentrations in salt marsh plants on the Suir Estuary, Ireland[J]. Environmental Pollution, 2003, 123(1):67-74.
[18]Stevens D P, McLaughlin M J, Heinrich T. Determining toxicity of lead and zinc runoff in soils:Salinity effects on metal partitioning and on phytotoxicity[J]. Environmental Toxicology and Chemistry, 2003, 22 (12):3017-3024.
[19]Inskeep W P, Comfort S D. Thermodynamic prediction for the effect of root exudates on metal speciation in the rhizosphere[J]. Plant, 1986, 9 (3-7):567-586.
[20]Nigam R, Srivastava S, Prakash S, et al. Cadmium mobilisation and plant availability: The impact of organic acids commonly exuded from roots[J]. Plant and Soil, 2001, 230(1):107-113.
[21]Aravind P, Prasad M N V. Cadmium-induced toxicity reversal by zincin Ceratophyllum demersum L.(a free floating aquatic macrophyte)together with exogenous supplements of amino -and organic acids[J]. Chemosphere, 2005, 61(11):1720-1733.
[22]Ma J F, Hiradate S. Form of aluminum for uptake and translocation in buckwheat(Fagopyrum esculentum Moench)[J]. Planta, 2000, 211 (3):355-360.
[23]Roser P T, Charlotte P, Juan B. Zinc hyperaccumulation in Thlaspi caerulescensⅡInfluence on organic acids[J]. Plant Nutrition, 1996, 19(12):1541-1550.
[24]刘爱荣,张远兵,钟泽华,等.盐胁迫对彩叶草生长和渗透调节物质积累的影响[J].草业学报, 2013, 22(3):211-218. LIU Ai-rong, ZHANG Yuan-bing, ZHONG Ze-hua, et al. Effects of salt stress on the growth and osmotic accumulation of Coleus blumei[J]. Acta Prataculturae Sinica, 2013, 22(3):211-218
[25]Weimberg R. Growth and solute accumulation in 3-week old seedlings of Agropyron elongatiunstressed with sodium and potassium salts[J]. Physiologia Plantarum, 1986, 67(2):129-135.
[26]丁能飞,傅庆林,刘琛,等.盐胁迫对两个大白菜品种抗氧化酶活性及离子吸收的影响[J].浙江农业学报, 2008, 20(5):322-327. DING Neng-fei, FU Qing-lin, LIU Chen, et al. Effect of salt stress on growth antioxidant enzyme activities, and in uptake of two Chinese cabbages cultivars[J]. Acta Agriculturae Zhejiangensis, 2008, 20(5):322-327.
[27]郭立泉,陈建欣,崔景军,等.盐、碱胁迫下星星草有机酸代谢调节的比较研究[J].东北师大学报(自然科学版), 2009, 41(4):123-128. GUO Li-quan, CHEN Jian-xin, CUI Jing-jun, et al. Comparative studies of metabolic regulation on organic acids in Puccinellia tenuif lora under salt and alkali stresses[J]. Journal of Northeast Normal University (Natural Science Edition), 2009, 41(4):123-128.
中图分类号:X171.5
文献标志码:A
文章编号:1672-2043(2016)05-0858-07
doi:10.11654/jaes.2016.05.006
收稿日期:2015-12-23
基金项目:国家自然科学基金项目(41371321);广东省产学研项目(2013B0906001)
作者简介:胡妮(1992—),女,四川广安人,硕士研究生,主要研究方向为环境修复技术与应用。E-mail:294080182@qq.com
*通信作者:李取生E-mail:liqusheng@21cn.com
Effects of salinity-inducted organic acid variation on Cd accumulation and salinity tolerance of edible amaranth
HU Ni1,2, CHEN Ke-han1,2, LI Qu-sheng1,2*, XU Zhi-min1,2,3, GUO Shi-hong1,2,3, YU Dan-ping1,2, LUO Tao1,2
(1.School of Environment, Jinan University, Key Laboratory of Environmental Exposure and Health of Guangzhou City, Guangzhou 510632, China; 2.Guangdong Provincial Research Center of Environmental Pollution Control and Remediation Material, Guangzhou 510632, China; 3.College of Life Science and Technology,Jinan University, Guangzhou 510632, China)
Abstract:Soil Cd pollution and soil salinization have severely threatened agricultural development and human health. It is important to study Cd accumulation and salinity tolerance of vegetables. Here a pot experiment was conducted to examine the influence of organic acid metabolism on Cd accumulation and salinity tolerance of two amaranth cultivars(Quanhong and Liuye)under 0.3%NaCl stress. Results showed that:Under salinity stress, the content of Cd in roots, shoots, and rhizospheric soil solution and the translocation ratios(from roots to shoots)of Cd in two cultivars increased compared with the control. Cadmium concentrations in Quanhong were obviously lower than those in Liuye. In addition, the content of Na+was significantly increased but K+absorption was obviously inhibited by salt stress; Under salinity stress, various organic acids were detected in roots, shoots and rhizospheric soil solution. Malic acid and oxalic acid were significantly increased, which were significantly positively correlated with Cd content in rhizospheric soil solution(P<0.05). In Liuye, the content of oxalic acid in shoots and acetic acid in roots was significantly increased, and were significantly positively correlated with the correspondling Cd content(P<0.05). These results suggest that Quanhong has lower Cd accumulation, stronger salinity tolerance, which was attributed to the variations of organic acids in the plants.
Keywords:salt stress; amaranth; organic acid; Cd accumulation; salinity tolerance
