16个红掌品种的核型分析
2016-05-30杨光穗冷青云王呈丹徐世松张志群
杨光穗 冷青云 王呈丹 徐世松 张志群

摘 要 采用常规压片的方法制备16个红掌品种的染色体标本,并进行核型分析。结果表明:16个红掌品种的染色体数目比较恒定,均为2n=2x=30。染色体主要由m染色体(中部着丝点区染色体)和sm染色体(亚中部着丝点区染色体)组成;最长染色体与最短染色体之比为1.83~3.07,平均臂比为1.95~2.29,臂比大于2的比例为43%~83%,核型不对称系数(As.K)为64.04%~68.17%,从核型类型来看,主要集中在2B型和3B型,只有‘紫公主为1A型。从核型分类上看,这些种类都属于较对称的类型。
关键词 红掌;染色体数;核型
中图分类号 Q949.717.2 文献标识码 A
红掌(Anthurium andraeanum)又名花烛、安祖花,为天南星科(Araceae)安祖花属(Anthurium)多年生草本植物,原产于中美洲及南美洲地区,是世界名贵花卉之一。因其绚丽多彩的心型佛焰苞和艳丽的肉穗花序、飘逸的叶形和持久的花期,红掌成为国际上流行的高档切花和盆花,风糜全球。1940年以来,世界各国纷纷开展红掌的引种、选种和育种研究,通过红苞花烛(A. Andraeanum)种内和花烛属种间杂交,对后代进行人工选育,获得了大量在花色、株形、花形等观赏性状上有所变异的杂种,具有较好的商业推广价格。红掌于1970年前后传入中国,特别是进入21世纪以来,随着国际、国内对热带花卉需求量的不断增加,国内红掌栽培面积不断扩大,其中以广东、上海、海南、陕西、河北、云南等地为主。目前有关红掌方面的研究主要集中在种质资源生物学特性观察[1-2]、栽培技术[3-4]、组织培养[5-7]、种质创新[8-9]、染色体数目分析[10]等方面,而在核型分析方面研究较少。本研究对16个红掌品种进行核型分析, 并研究其体细胞染色体数及染色体形态特征,探讨核型参数在不同红掌品种中总体上的规律与差异,为以后研究红掌品种的遗传变异、种间亲缘关系鉴定及新品种选育等提供依据。
1 材料与方法
1.1 材料
本实验供试的16个红掌品种来自于中国热带农业科学院热带作物品种资源研究所花卉种质资源圃。具体见表1。
1.2 方法
体细胞染色体标本制备参照冷青云等[11]的方法。镜检并统计染色体数后选取染色体分散、形态好的细胞在Leica DM2500生物显微镜下观察并拍照(拍照相机为该显微镜自带数码成像系统),使用photoshop7.0软件处理后作核型分析。染色体数目确定及核型分析方法按李懋学等[12]的规定,统计30个以上细胞,以其中85%以上细胞具有的恒定一致的染色体数来作为该种的染色体数。每个类群取5个细胞的核型平均值,核型不对称系数(As.K)按Arano[13]的方法,比值越大越不对称;核型分类按Stebbins[14]的方法划分。
2 结果与分析
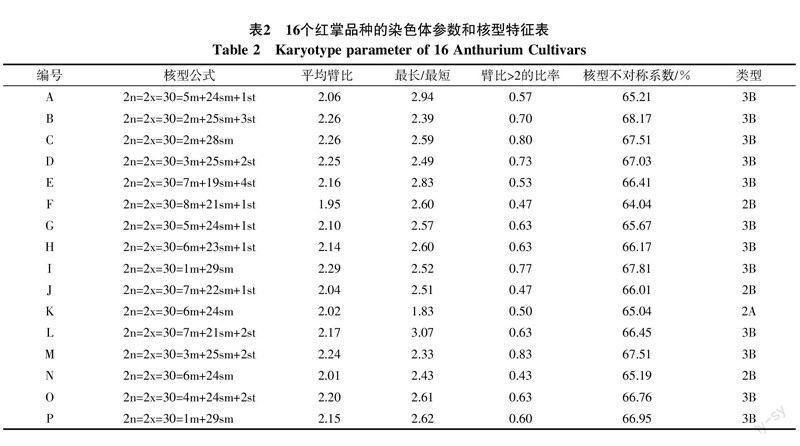
16个红掌品种的的染色体参数和核型特征见表2,染色体形态图、核型图分别见图1,图2。
16个红掌品种染色体数目均为2n=30,核型主要组成部分为中部着丝点区染色体(m)和亚中部着丝点区染色体(sm);此外,有些种具有数量不等的亚端部着丝点区染色体(st),平均臂比为1.95~2.29,其中臂比值最小的品种是欢呼,臂比均值为1.95;红塔的臂比均值最大,为2.29。最长染色体与最短染色体之比为1.83~3.07,其中紫公主最长染色体与最短染色体比值最小,为1.83;情迷最长染色体与最短染色体比值最大,为3.07。臂比大于2的比例占0.43%~0.83%,核不对称系数为64.04%~68.17%,从核型类型来看,主要集中在2B型和3B型,只有‘紫公主为1A型。从核型分类上看,这些种类都属于较对称的类型。
3 讨论
染色体数目及特征是重要的生物学数据,同一物种或品种的染色体数目比较恒定,对阐述植物进化程度和分类方面有重要意义。红掌栽培品种主要由红苞花烛(A. andraeanum)种内或与之亲缘关系较近的种间杂交选育而来,据Gaiser[15]、Sharma等[16]、Bliss等[17]报道,红苞花烛及其所在的Calomystrium组内其他近缘种A. amnicola、A. armeniense、A. crystallinum、A. antioquiense、A. esmeraldense、A.formousun等染色体数均为2n=30[15-17],本研究中16个红掌品种的染色体数目均为2n=2x=30,与肖建忠等[10]报道的3个品种‘亚美哥、‘瓦伦天奴、‘斯克斯染色体数目一致。与栽培菊花、蝴蝶兰[18-20]等花卉品种常出现非整倍体相比,红掌在染色体数目上比较恒定,这可能与红掌选育代数较少,且主要是由红苞花烛种内杂交选育而成有很大关系。而菊花是由原产于中国的几个野生种天然杂交,再经过人工选育而成的栽培杂交复合体,其染色体数目具有多样性;蝴蝶兰栽培品种是由多代的属间或种间杂交选育而成,由于双亲的染色体缺乏同源性,导致在减数分裂时配对紊乱,后代染色体分配不均衡,从而导致染色体数目不恒定。本研究尚未发现B型染色体的存在,但Carlsen[21]发现可与红苞花烛杂交的Cardiolonchium组内原生种存在B型染色体,这可能是由于本研究所用的材料不含有这些原生种的血统,也可能是由于在不断的杂交选育过程中B染色体丢失,或者由于本次试验条件有限,未观察到B染色体的存在。
核型分類用于区分核型的对称或不对称程度[12]。本研究中,16个红掌品种出现了3种核型类型:2A、2B、3B型,除‘紫公主为2A型,‘欢呼、‘热情、‘神秘3个品种为2B型外,其他12个品种均为3B型。2A型相对于2B型和3B型来说,是极对称的核型,表明红掌的染色体核型类型存在多样性,也说明‘紫公主与其他品种遗传背景相差较远,这与王呈丹[22]用SRAP分子标记对红掌品种进行遗传多样性和亲缘关系分析的结果相符,即‘紫公主与其他品种差异较大。从外部形态上看也可以验证这一点,‘紫公主株型松散,叶片墨绿,呈窄卵型,佛焰苞为郁金香型;其他品种株型紧凑,叶片多为卵型和阔卵型,佛焰苞为心型。‘紫公主可能由Calomystrium组内或者Cardiolonchium组内、Porphyrochitonium组内具有郁金香型佛焰苞的原生种与来源于红苞花烛的红掌品种杂交选育而来[23],具心型佛焰苞的品种血统主要来源于红苞花烛,因为红苞花烛是花烛属中唯一具有佛焰苞的原生种[17]。综合以上因素可将‘紫公主与其他品种区分开来,将其作为特异种质保存及加以利用,为红掌的进一步遗传改良提供材料基础。
参考文献
[1] 徐世松, 王呈丹, 黄素荣, 等. 红掌种质资源形态学标记与遗传多样性分析[J]. 热带作物学报, 2014, 35(10): 1 890-1 896.
[2] 常娟霞, 牛俊海, 黄少华, 等. 红掌种子结实性、 发育周期及杂交后代发芽力的变异分析[J]. 基因组学与应用生物学, 2014, 33(2): 392-397.
[3] 单芹丽, 赵 辉, 邓君浪, 等. 红掌的栽培与管理技术[J]. 北方园艺, 2003(2): 38-39.
[4] 陈春满, 郑贵朝, 张善信, 等. 不同栽培基质对红掌组培苗移栽成活及生长发育的影响[J]. 广东农业科学, 2008(2): 28-29.
[5] 兰芹英, 仇玉萍, 张运祥, 等. 不同红掌品种的叶片、 叶柄和茎段愈伤组织的诱导及植株再生[J]. 西北植物学报, 2003, 23(6): 1 006-1 009.
[6] 姜 雷, 梁彩虹, 张志胜, 等. 影响红掌愈伤组织诱导、 增殖和芽分化的因素[J]. 种子, 2006, 25(11): 26-30.
[7] 陈彦霖. 红掌叶片愈伤组织诱导及植株再生的优化[J]. 湖北农业科学, 2016, 55(6): 1 572-1 574.
[8] 张志胜, 黎扬辉, 姜 蕾, 等. 红掌四倍体的离体诱导及其鉴定[J]. 园艺学报, 2007, 34(3): 729-734.
[9] 徐洪伟. 利用红掌2n花粉创造三倍体新种质的研究[D]. 海口: 海南大学, 2015.
[10] 肖建忠, 尚爱芹, 陈段芬, 等. 花烛属植物的核型分析[J]. 河北农业大学学报, 2003, 26(2): 33-36.
[11] 冷青云, 莫 饶, 彭 彬, 等. 六种兰属植物的核型分析[J]. 园艺学报, 2009, 36(2): 291-296.
[12] 李懋学, 陈瑞阳. 关于核型分析的标准化问题[J]. 武汉植物学研究, 1985, 3(4): 297-302.
[13] Arano H. Cytological studies in subfamily Carduoideae(Compositae)of Japan. IX. The karyotype analysis and phylogenetic considerations on Pertya and Ainsliaea[J]. Botanic Magazine(Tokyo), 1963, 76: 32-39.
[14] Stebbins G L. Chromosome Evolution in Higher Plants[M]. London: Edward Arnold, 1971: 88.
[15] Gaiser, L O. Chromosome number and species characters in Anthurium[J]. Proc Trans R Soc Can, 1927, 21: 1-137.
[16] Sharma A K, Bhattacharyya U C. Structure and behavior of chromosomes in species of Anthurium with special reference to the accessory chromosomes[J]. Proceedings of the National Institute of India, Part B, Biological Science, 1961, 27: 317-328.
[17] Bliss B J, Suzuki Y J. Genome size in Anthurium evaluated in the context of karyotypes and phenotypes[J]. AoB Plants, 2012: 1-14.
[18] 李 畅, 陈发棣, 赵宏波, 等. 栽培小菊17个品种的核型分析[J]. 园艺学报, 2007, 35(l): 71-80.
[19] 张 迪, 朱根发, 叶庆生, 等. 50份蝴蝶兰种质的染色体数目与倍性分析[J]. 热带作物学报, 2013, 34(10): 1 871-1 876.
[20] 朱明丽. 大菊品种的细胞分类学研究[D]. 北京: 北京林業大学, 2010.
[21] Carlsen M. Understanding the origin and rspid diversification of the genus Anthurium Schott(Araceae), integrating molecular phylogenetics, morphology and fossils[M]. PHD Dissertdtion, university of Missouri, st Louis, MO, USA, 2011.
[22] 王呈丹. 红掌品种亲缘关系分析与细胞学鉴定[D]. 海口: 海南大学, 2015.
[23] Kamemoto H, Kuehnle A R. Breeding Anthuriums in Hawaii[M]. Honolulu: University of Hawaii Press, 1996.
