植物气孔导度的环境响应模拟及其尺度扩展
2016-05-23高冠龙张小由常宗强鱼腾飞
高冠龙, 张小由, 常宗强, 鱼腾飞, 赵 虹
1 中国科学院寒区旱区环境与工程研究所, 兰州 730000 2 中国科学院大学, 北京 100049 3 浙江瑞启检测技术有限公司, 杭州 310000
植物气孔导度的环境响应模拟及其尺度扩展
高冠龙1,2, 张小由1,*, 常宗强1, 鱼腾飞1, 赵虹3
1 中国科学院寒区旱区环境与工程研究所, 兰州7300002 中国科学院大学, 北京1000493 浙江瑞启检测技术有限公司, 杭州310000
摘要:气孔导度是衡量植物和大气间水分、能量及CO2平衡和循环的重要指标,探讨气孔导度与环境因子的关系及其模拟,以及气孔导度在叶片、冠层及区域尺度间的尺度转换及累积效应,对更好地认识植被与大气间的水热运移过程,合理评价植被在陆面过程中的地位和作用都具有重要意义。从植物气孔导度与环境因子的关系、气孔导度模拟以及尺度扩展三个方面,对前人的研究成果进行了概括总结。从叶片和冠层两个尺度出发,归纳总结了前人对于不同植物气孔导度与环境因子关系的研究成果,发现由于不同植物的遗传特性、测定时的环境、时间尺度的不同,以及未考虑各个环境因子的相互作用对气孔导度的影响,由此得到的气孔导度与环境因子之间的关系也不尽一致。对各单一环境因子与气孔导度的关系,给出了生理学解释,从根本上说明了环境因子变化对气孔导度的影响,而研究环境因子对气孔导度的综合影响时,应对各环境因子进行系统控制与同步观测。模拟计算植物气孔导度的模型主要有Jarvis模型和BWB模型两类,这些模型的模拟能力随着研究对象、试验区域、环境条件的改变而存在一定的差异,在具体使用时应结合实际情况选择最优模型进行模拟。除上述常用模型外,还总结了其他学者分别从不同角度提出的新的模型,对现有气孔导度模型进行了全面的总结。从叶片-冠层、冠层-区域两个方面归纳总结了前人关于气孔导度尺度扩展的研究成果,发现叶片-冠层的尺度扩展研究较成熟而冠层-区域的尺度扩展在模拟精度的验证方面存在困难。针对以下几个方面提出了今后气孔导度的研究重点:(1)结合研究对象所在的区域及环境条件,选择最优模型进行模拟;(2)综合考虑环境因子之间的相互作用及其对气孔导度的累积影响;(3)BWB模型与光合模型的耦合;(4)提高大尺度范围内的气孔导度模拟精度。
关键词:气孔导度;环境响应模拟;环境因子;尺度扩展
气孔是植物与大气间进行碳水交换的通道[1],气孔行为及其模拟对于分析生态过程、陆面过程或水循环过程等都具有非常重要的意义[2]。气孔影响着蒸腾和光合等生理机能[3],随所处的环境状况而时刻发生着变化,在植物中起平衡调节作用[4]。气孔的运动状况一定程度就反应了植物体内的代谢情况,气孔的灵敏度也是植物的一个重要抗旱特征[5]。随着陆-气相互作用中植被的作用逐步被认识,关于气孔导度的研究成为一个新的热点。笔者归纳总结发现,目前国内外学者关于气孔导度的研究主要集中于气孔导度与环境因子的关系、气孔导度的模拟以及气孔导度的尺度扩展等方面。本文将重点从这几个方面展开讨论,在总结前人研究成果的同时分析相关研究中的不足之处,以期为气孔导度的进一步研究提供科学依据。
1气孔导度与环境因子的关系研究
1.1叶片气孔导度与环境因子的关系
气孔在植物中起着平衡调节的作用,它是调控土壤-植被-大气这个连续体之间物质和能量交换的关键环节,弄清叶片气孔导度与环境因子之间的相互关系是探讨植物的能量和水分交换动态的基础[6]。迄今为止,我国已有大量学者研究了植物叶片气孔导度与环境因子之间的关系。
1.1.1叶片气孔导度与单一环境因子的关系
关于大气CO2浓度对植物叶片气孔导度的影响,许多学者认为高浓度CO2可降低植物叶片气孔导度,但气孔导度对高浓度CO2的响应随植物种的不同和环境条件的不同而变化。郑凤英[7]等总结发现,在CO2倍增时,大豆气孔导度下降25%[8],白桦气孔导度下降21%[9],春小麦在550mg/kg时的气孔导度比370 mg/kg时下降36%[10],而海棠、刺叶栎、红槲斗栎几乎没有变化。银白槭在温度>33℃时才下降[11],红栎在土壤水分良好和亏缺时下降均明显[12],纸桦、美洲落叶松、黑云杉、美洲短叶松在温度范围为(日/夜)18/12—30/24℃时的降低幅度为10%—25%[13],但挪威云杉几乎无影响[14]。王建林[15]等针对9种植物的叶片气孔导度对CO2浓度变化响应进行了模拟,结果表明随着CO2浓度的升高,气孔导度会逐渐降低,且下降的幅度会随着CO2浓度的升高而逐渐减弱。
光照强度对叶片气孔导度的影响表现为正相关关系,即在一定范围内气孔导度随着光照强度的增加而增大。陶汉之[16]等针对茶树叶片的研究发现,气孔导度随光强度增加而增大,反之减小。气孔导度与光量子通量密度之间呈显著和极显著正相关。申双和[17]等对棉花阴、阳叶的研究发现,气孔导度因叶片受光强度的不同而不同,阴叶的气孔导度总是小于阳叶。
水分对叶片气孔导度的影响表现为水分的降低会对气孔导度产生抑制作用。柯世省[18]等研究了云锦杜鹃叶片气孔导度对水分的响应,结果表明随着干旱胁迫程度的增强,云锦杜鹃叶片气孔导度明显降低。
单一环境因子对植物叶片气孔导度的影响以及生理学解释见表1。

表1 植物气孔导度与环境因子的关系及生理学解释
1.1.2叶片气孔导度与综合环境因子的关系
由于不同植物的遗传特性、测定时的环境、时间尺度的不同,以及未考虑各个环境因子的相互作用对气孔导度的影响,由此得到的气孔导度与环境因子之间的关系也不尽一致[19- 20]。单一环境因子对气孔导度的影响国内外学者已经进行了大量研究,然而多个环境因子对气孔导度的综合影响及主导因子的确定却鲜有研究。
左应梅[21]等分析了土壤相对含水量、光合有效辐射和空气相对湿度对华南8号木薯的综合影响,结果发现当土壤相对含水量较低时,土壤相对含水量是影响气孔导度的主导因子,随着土壤相对含水量的增加,影响气孔导度的主导因子由土壤相对含水量转变为光合有效辐射,存在过渡临界值,而空气湿度受其他环境因子的干预相对较小。卢振明[22]等通过对冬小麦气孔导度与土壤含水量、光照强度、气温及空气相对湿度的综合影响关系的研究,得出了相反的结论,认为土壤水分含量较低时,光照强度、气温及空气相对湿度对气孔导度的影响显著,随着土壤含水量升高,土壤相对含水量是影响气孔导度的主导因子。以上学者得出相反结论的原因可能是研究方法和条件的不同,在研究环境因子对气孔导度的综合影响时,应对各环境因子进行系统控制与同步观测,而不能简单采用测定当日的平均值。因此,确定不同条件下影响气孔导度的主导因子以及探讨环境因子对气孔导度的综合影响机理将成为下一步研究的重点。
1.2冠层气孔导度与环境因子的关系
冠层气孔导度是森林生态系统响应环境变化的敏感性指标,可被用来监测水分胁迫的变化及其对森林生态功能的影响,是进行生态、气象和空气质量监测模拟时使用的重要参数[23- 24]。冠层气孔导度在研究和模拟冠层尺度或更大尺度的冠层与大气间的相互作用方面也是一个极其重要的参数[25]。对较小的时间尺度而言,光辐射和水汽压亏缺是驱动冠层气孔导度变化的主要因子,气温、土壤水分条件也是影响冠层气孔导度变化的重要因素[23]。
朱丽薇[26]等研究了荷木人工林的冠层气孔导度对环境因子的响应,结果表明气孔导度与光合有效辐射呈线性正相关;当光合有效辐射>1000μmol m-2s-1且水汽压亏缺>2kPa时,气孔导度与水汽压亏缺呈线性负相关;气孔导度与土壤含水量没有显著的相关性。赵平[27-28]等利用树干液流值,以边材面积为转换因子计算了马占相思林的蒸腾量和单位叶面积蒸腾速率等参数值,然后根据其他学者提出的经验公式计算了森林冠层的气孔导度,进而分析了冠层气孔导度对环境因子的响应,结果表明冠层气孔导度最大值随水汽压亏缺的上升呈对数函数下降的趋势,对光合有效辐射的响应则呈双曲线函数增加的趋势。李仙岳[29]等研究了樱桃冠层气孔导度对环境因子的响应,结果表明太阳辐射增大总体上会导致樱桃冠层气孔导度变大,特别是不同辐射条件下冠层气孔导度的最低值明显是随着辐射强度变大而变大,当太阳辐射升到300W/m2后,太阳辐射对冠层气孔导度的影响明显降低;当温度在20—30℃时,樱桃冠层气孔导度较大,升高或降低温度都会导致冠层气孔导度下降;冠层导度也随着水汽压亏缺呈下降趋势。
2气孔导度模型介绍及应用研究
2.1常用气孔导度模型及应用
目前,被学术界广泛用于估算气孔导度对环境因子响应的模型分为Jarvis模型[30]和BWB模型[31]两类,随后,Leuning等对BWB模型进行了改进,提出了BBL模型[32],其实质仍是BWB模型。Jarvis模型是经验模型,而BWB模型和BBL模型是考虑了植物生理活动影响的半经验模型。
2.1.1Jarvis模型
Jarvis模型在模拟叶片气孔导度时因时间尺度和环境条件的不同而表现出明显差异,环境变量的变化会对模型模拟结果产生影响。齐华[33]等在Jarvis模型基础上构建了柑橘叶片气孔导度对主要环境因子的响应模型,结果表明月份尺度和全年尺度上用Jarvis模型模拟柑橘叶片气孔导度时,环境变量必须包括光合有效辐射在内的2个及其以上环境变量。温度较适宜的生长季,为使模型模拟具有较高精确度,除光合有效辐射外,环境变量还应包含饱和水汽压差或叶片温度;温度、大气中CO2浓度变化较大的季节,环境变量对Jarvis模型精确度的影响较为复杂,除光合有效辐射外,至少需包含饱和水汽压差、叶片温度、大气中CO2浓度3个环境变量中的某个变量。Jarvis模型不仅可以模拟叶片尺度的气孔导度,也可以模拟冠层尺度的气孔导度。朱仲元[34]等基于Jarvis模型研究了杨树种群气孔导度与环境因子之间的响应关系,在此基础上用S-W双涌源模型以小时为时段模拟计算了浑善达克沙地天然杨树种群的蒸散量,取得了较好的结果。Granier[35]等基于Jarvis模型,构建了冠层平均气孔导度模型,孙林[36]等运用这一模型对华北落叶松冠层平均气孔导度进行了模拟,取得了良好的结果。韩磊[37]基于Jarvis模型模拟计算了黄土半干旱区主要造林树种(侧柏、油松和白榆)的冠层气孔导度,其模拟结果比Oguntunde[38]等描述的线性模型和Lu[39]等描述的负指数关系模型的模拟结果要好得多。许文滔[40]等基于Jarvis模型模拟计算了华南地区马占相思林的冠层气孔导度,取得了良好的结果。
2.1.2BWB模型
苏永红[41]等基于BWB模型模拟计算了额济纳荒漠河岸胡杨林叶片的气孔导度,并与实测的气孔导度进行了比较,结果表明二者之间呈密切线性关系,说明胡杨叶片的气孔导度与光合速率的关系符合BWB模型的理论基础。李红生[42]等基于BWB模型模拟计算了黄土丘陵沟壑区沙棘叶片的气孔导度,利用沙棘旱区的气象资料及未参与模型参数拟合的沙棘叶片气孔导度实测资料对所建模型进行了验证,结果表明该模型能较好地模拟沙棘叶片气孔导度的变化。周莉[43]等基于BWB模型和非直角双曲线光合模型模拟计算了盘锦湿地芦苇叶片的气孔导度,该模型考虑了气孔导度与光合之间的相互作用,利用实验数据对气孔导度模型进行了验证,取得了良好的效果。
2.1.3BBL模型
袁国富[2]等基于BBL模型,确定了适合于冬小麦气孔导度模拟的空气湿度、土壤湿度和植物水势响应函数参数,为各类气孔导度模型的使用提供了参考。
2.2常用气孔导度模型及扩展模型应用
以上模型虽已被广泛应用于不同尺度的气孔导度模拟中,但是学者们研究发现,这些模型的模拟能力随着研究对象、试验区域、环境条件的改变而表现出一定的差异[44]。
2.2.1常用气孔导度模型应用比较
魏征[45]等对华北地区冬小麦叶片气孔导度进行了模拟,结果表明BBL模型较Jarvis模型能更好地解释华北地区冬小麦叶片气孔导度对环境因子的响应变化。郑怀舟[46]等基于Jarvis模型和BWB模型,对长汀水土流失区马尾松与木荷的气孔导度进行了模拟,经实测数据验证后结果表明这两个树种的气孔导度最优模拟模型均为BWB模型。BWB模型构建的前提是气孔导度与净光合速率之间呈线性关系,因此这两位学者基于这一前提进行的实验其结论均表明BWB模型的模拟结果较好。而在干旱半干旱或者典型高海拔山地气候条件下,土壤含水量和气温决定了植物气孔的开闭,进而限制了CO2的同化速率,这与BWB模型的构建前提不一致。因此,BWB模型在这些区域没有Jarvis模型的模拟效果好。王玉辉[47]等对松嫩平原草地的羊草叶片气孔导度进行了模拟,结果表明Jarvis模型比BBL模型具有更好的模拟效果。唐凤德[48]等基于Jarvis模型和BWB模型,对长白山阔叶红松林叶片的气孔导度进行了模拟,并用实测数据进行验证,结果表明Jarvis模型比BWB模型更适于阔叶红松林叶片的气孔导度模拟。另外,鱼腾飞[49]等研究了极端干旱区多枝柽柳叶片气孔导度对空气湿度的响应,结果表明BWB模型拟合效果较好,BBL模型结果显著但效果较差。
2.2.2常用气孔导度模型与扩展模型应用比较
很多学者在Jarvis模型和BWB模型的基础上推导了新的模型,还有部分学者从数学方程和生物机理角度出发提出了新的模型,对植物气孔导度进行了模拟。李永秀[50]等对扬麦13的气孔导度及相关生理生态指标进行了观测,分别从Jarvis模型和BWB模型中各选用两组不同的响应函数表达式进行模拟,结果表明Jarvis模型1模拟效果最好。王治海[51]等利用Jarvis模型和BWB模型对冬小麦品种鲁麦23进行了气孔导度模拟比较,结果表明BWB模型2预测精度最高,而BWB模型1预测效果最差。气孔导度模型在不同地区的适应性差异通过作物品种直接表现,但不同小麦品种适应于不同地区,因此,模型的适应性差异应当归结为品种和地域差异的综合表现,至于这种差异的最主导因素究竟是品种还是地域则有待进一步研究。杨再强[52]等基于BWB模型模拟计算了温室番茄气孔导度,与用单隐层的BP神经网络模型模拟的结果进行比较,结果表明BP神经网络模型对番茄品种“大红”、“苏粉8号”的叶片气孔导度的模拟值与实际观测值基于1∶1线的决定系数高于BWB模型,说明所建立的BP神经网络模型模拟叶片气孔导度的精度明显高于BWB模型。米娜[53]等基于BWB模型、BBL模型以及Wang[54]等利用生态系统通量观测数据在检验BWB模型和Leuning模型的基础上提出的两个新模型:power-h模型和power-D模型,对玉米的气孔导度进行了模拟,结果表明BWB模型在空气较为湿润的环境条件下,模拟结果存在较大的低估;BBL模型降低了模拟结果的误差,但仍然不能很好地描述气孔导度在较湿润状况下的显著升高;power-h模型和power-D模型的模拟精度较高。石建红[55]等组合了14种气孔导度模型,对鄱阳湖流域的典型树种(杉木、湿地松、马尾松、沉水樟、柑橘)夏季的气孔导度进行了模拟,结果表明杉木、湿地松、马尾松的最优模拟模型均为BBL-B2模型;沉水樟的最优模拟模型为BBL-B1模型;柑橘的最优模拟模型为Jarvis-J12模型。叶子飘[56]等从CO2分子在叶片气孔中扩散这个最基本的物理过程出发,应用物理学中的分子扩散和碰撞理论、流体力学与植物生理学等知识,推导出叶片气孔导度的机理模型,基于此模型以及BWB模型对冬小麦气孔导度进行了模拟,结果表明推导的气孔导度机理模型与BWB模型相比能更好地描述冬小麦的气孔导度与净光合速率之间的关系。
以往学者大多没有考虑各模型的适用条件及优缺点,因而模拟结果会产生显著差异。表2对各模型的表达式、参数意义、适用条件及优缺点进行了总结,在今后运用气孔导度模型进行模拟时,应结合试验设计方案及环境条件选择合适的模型进行模拟。
2.3其他气孔导度模型
在理论研究中,除学者们普遍认可的Jarvis、BWB和BBL模型之外,部分学者还从不同的角度出发提出了一些新的气孔导度模型。Yu[57]等在BWB模型的基础上,引入了新的参数(总同化率Ag),提出了一个新的气孔导度模型。该模型与Jarvis模型和BWB模型相比,从模拟效果与模型结构上均得到了提升。与BWB模型相比,通过对原始模型中部分参数的修正,使得模拟效果更好;与Jarvis模型相比,在同样考虑了多个环境因子对气孔导度影响的同时,模型结构更加简单。Belinda E. Medlyn[58]等耦合了气孔导度经验模型与最优气孔控制模型,提出了一个新的气孔导度模型,该模型既具有与经验模型相似的表达式,也拥有与最优模型相同的推导过程。通过模拟验证,结果显示该模型可以很好的解释不同环境条件下的气孔行为,提高了模拟精度。另外,傅伟和王天铎[59]基于气孔运动的机理模型,建立了模拟VPD、CO2、土壤水势、光照和温度对气孔影响的数学模型。该模型的优势在于可以在不同组合的环境因子下模拟气孔导度,也可作为研究气孔整体行为的理论框架。表3列出了上述几个新的气孔导度模型的表达式及各参数的意义。
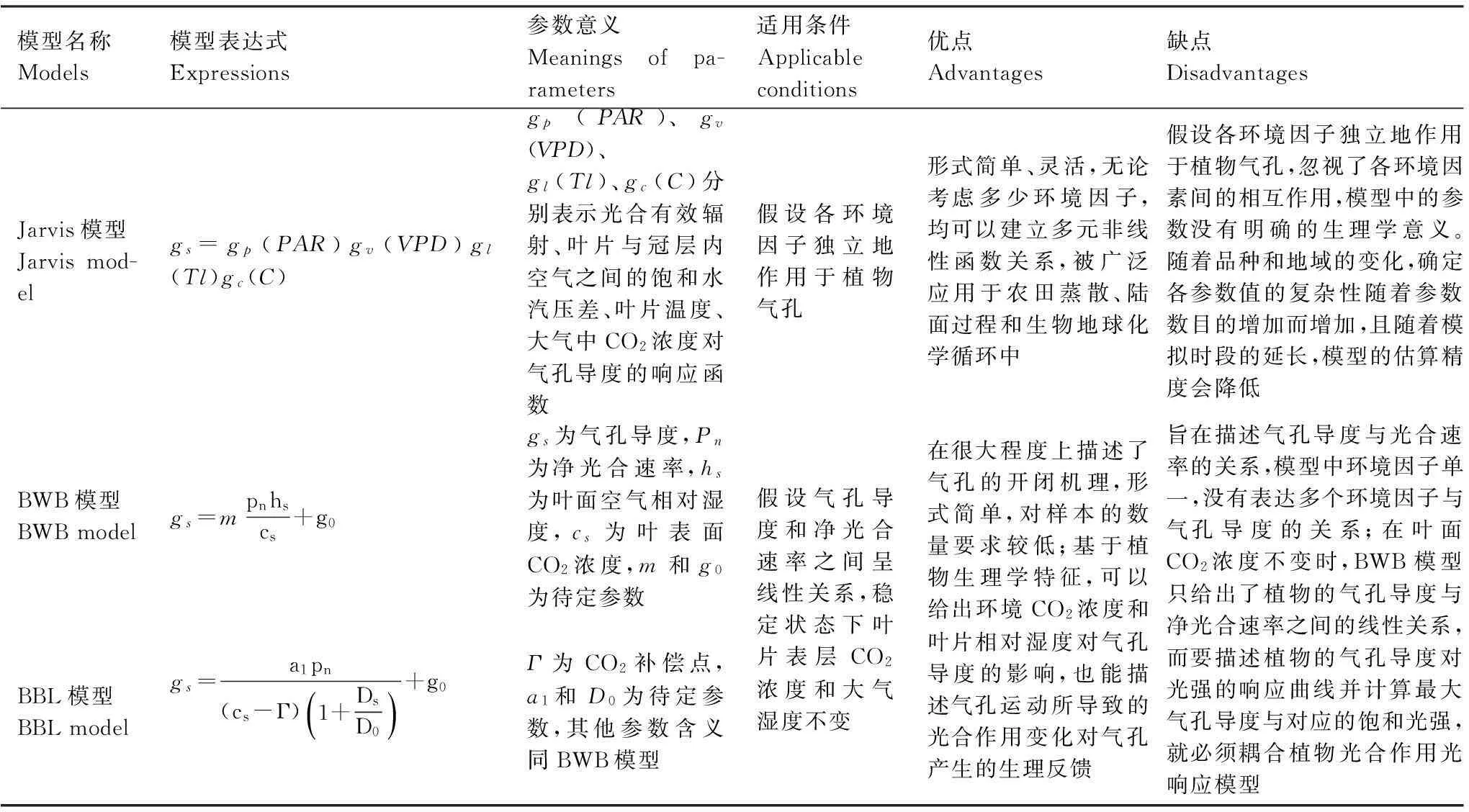
表2 Jarvis模型、BWB(Ball-Woodrow-Berry)模型及BBL(Ball-Berry-Leuning)模型介绍

表3 其他气孔导度模型的表达式及参数意义
3气孔导度尺度扩展研究
叶片气孔导度模拟及其向冠层导度的尺度提升是实现蒸散发尺度转换的基础。探讨气孔导度在叶片、冠层及区域尺度间的转换及累积效应,对更好地认识植被与大气间的水热运移过程、合理评价植被在陆面过程中的地位和作用具有重要意义[60]。
3.1叶片-冠层气孔导度扩展3.1.1基于气体交换特性的尺度扩展
迄今,冠层气孔导度的模拟研究大多基于叶片尺度的气孔导度对环境响应的乘合模型或气孔导度与光合的耦合模型[61]。许多学者基于Penman-Monteith(P-M)模型,引入了叶面积指数、光合有效辐射等参数,对冠层气孔导度进行了模拟计算。P-M模型假定植被是由单层的大叶片构成的,并且蒸发面是水分的单涌源。黄辉[62]等通过引入叶面积指数,基于Yu[63]等提出的Jarvis模型的优化模型,提出了由潜在气孔导度和相对气孔开度乘积形式表达的组合模型,模拟了华北平原冬小麦农田生态系统的冠层气孔导度,并用P-M方程估算的表面导度进行了验证,结果表明二者具有较好的一致性。曹庆平[64]等基于Jarvis和Mcnaughton[65]提出的P-M方程的改进形式,模拟计算了华南荷木林冠层导度,通过叶面积指数转换求出了冠层气孔导度,结果表明与气孔气体交换方法获得的冠层气孔导度有较好的一致性。刘薇[66]等引用Sellers推导的冠层气孔导度计算公式,模拟计算了褒河流域不同植被的冠层气孔导度,引入基于叶面积指数的P-M方程,模拟了褒河流域蒸散发能力的时空分布,分析比较了不同植被覆盖下蒸散发能力的变化规律,取得了良好的结果。张宝忠[67]等以光合有效辐射作为尺度转换因子,基于P-M方程,在假定下垫面均匀分布且忽略土壤蒸发影响以及冠层内水汽压亏缺变化状况下,构建了冠层气孔导度估算模型,结果表明基于光合有效辐射和饱和水汽压差构建的冠层气孔导度估算模型可较好地实现从叶片气孔导度向冠层气孔导度的尺度转换提升,但是模拟值与实测值之间存在一定误差,且冠层气孔导度越大误差越显著,这或许是由于利用 P-M 方程反推冠层气孔导度的过程中含有来自地表阻力的影响,而采用尺度提升方法模拟的冠层气孔导度中却并未考虑土壤蒸发所致。除P-M模型外,任传友[68]等对叶片尺度上的气孔导度-光合-蒸腾耦合模型(SMPT-SB)在冠层尺度上进行了扩展,探讨了模型的参数化方法,建立了冠层尺度上的生态系统光合-蒸腾耦合模型,并用涡度相关观测数据对模型进行了验证,取得了良好的结果。
3.1.2基于冠层蒸腾量的尺度扩展
基于液流测定值,以边材面积为转换因子计算得到的冠层蒸腾量,或者通过波文比-能量平衡法和涡度相关技术直接测算得到的冠层蒸腾量,利用经验公式或梯度原理便可间接算出冠层气孔导度。由于目前还没有直接测定冠层气孔导度的仪器和方法,因此,基于冠层蒸腾量计算得到的冠层气孔导度值可以用来对基于气体交换方法所得冠层气孔导度模拟结果进行验证。
3.2冠层-区域气孔导度扩展
冠层-区域的尺度转换是大尺度大气环流模式对陆面下边界条件进行空间平均时重点需要解决的问题[69],遥感结合地面观测资料可以较为准确地估测诸如气孔导度等植被陆面过程参数。遥感的优势在于频繁和持久地提供地表特征的面状信息,这对于传统的以稀疏离散点为基础的对地观测手段是一场革命性的变化[70],同时也为冠层-区域导度的尺度转换提供了良好的技术平台[71- 75]。张佳华[76]等利用NOAA-AVHRR数据并结合遥感,根据计算气孔导度的半理论半经验公式,估算了冬小麦各生育期的平均气孔导度,并将计算结果扩展为区域尺度范围内的研究。结果表明冬小麦返青-拨节期气孔导度较低,长势较好的南部气孔导度高于长势较差的西北部。在气候干旱的季节,华北部分地区气孔导度下降;随着降雨量增多,华北北部地区气孔导度上升。中南部处于收获期,气孔导度呈下降趋势。然而,基于遥感反演所得到的结果与真实值之间本身存在一定的差距[77],由此通过公式计算得到气孔导度值其不确定性将增大。
4结论与展望
目前,国内外学者关于植物气孔导度的研究主要集中于气孔导度与环境因子的关系、气孔导度的模拟以及尺度扩展等方面。随着越来越多的学者对模型进行了修正,各气孔导度模型的模拟精度也越来越高。但是,以往的研究中仍然存在一些不足之处:
(1)气孔导度模型的使用。无论是经验模型还是半经验模型,在对气孔导度进行模拟时,应综合考虑研究对象所在的区域及环境条件,选择最优模型进行模拟。
(2)气孔导度的环境响应。在分析植物气孔导度与环境因子的关系时,大多学者对单一环境因子与气孔导度的关系进行了分析,未来的研究应综合考虑环境因子之间的相互作用及其对气孔导度的累积影响。
(3)BWB模型与光合模型的耦合。运用BWB模型进行模拟时,需要输入净光合速率值,而实际应用中,光合速率的长期测量和气孔导度一样不便。现有的各种尺度模型预测中,光合速率通常由模型计算得到,导致BWB模型在应用时必然要涉及到光合作用模型,后者的参数数量和预测精度必然会影响到前者。因此,BWB模型与光合模型的耦合问题将成为后续研究的重点。
(4)模拟结果验证。目前,关于叶片尺度的气孔导度模拟结果的验证较为成熟,基于实测数据的验证精度也较高,而关于冠层或者更大尺度的气孔导度模拟结果由于实测数据较难获取,对其模拟精度的验证也较为困难,如何提高大尺度范围内的气孔导度模拟精度将是下一步研究的重要方向。
参考文献(References):
[1]石培华, 冷石林, 梅旭荣. 气孔导度、表面温度的环境响应模型研究. 中国农业气象, 1995, 16(5): 51- 54.
[2]袁国富, 庄伟, 罗毅. 冬小麦叶片气孔导度模型水分响应函数的参数化. 植物生态学报, 2012, 36(5): 463- 470.
[3]董树亭, 胡昌浩, 周关印. 玉米叶片气孔导度、蒸腾和光合特性研究. 玉米科学, 1993, 1(2): 41- 44.
[4]拉夏埃尔W. Plant Ecophysiology. 北京: 科学出版社, 1981.
[5]张守仁, 高荣孚. 白杨派新无性系气孔生理生态特性的研究. 生态学报, 1998, 18(4): 358- 363.
[6]王芸, 吕光辉, 高丽娟, 任曼丽, 苏前, 孙丽君. 荒漠植物白麻气孔导度特征及其影响因子研究. 干旱区资源与环境, 2013, 27(8): 158- 163.
[7]郑凤英, 彭少麟. 不同尺度上植物叶气孔导度对升高CO2的响应. 生态学杂志, 2003, 22(1): 26- 30.
[8]Volin J C, Reich P B, Givnish T J. Elevated carbon dioxide ameliorates the effects of ozone on photosynthesis and growth: species respond similarly regardless of photosynthetic pathway or plant functional group. New Phytologist, 1998, 138(2): 315- 325.
[9]Rey A, Jarvis P G. Long-term photosynthetic acclimation to increased atmospheric CO2concentration in young birch (Betulapendula) trees. Tree Physiology, 1998, 18(7): 441- 450.
[10]Garcia R L, Long S P, Wall G W, Osborne C P, Kimball B A, Nie G Y, Pinter P J, Lamorte R L, Wechsung F. Photosynthesis and conductance of spring- wheat leaves: field response to continuous free-air atmospheric CO2enrichment. Plant, Cell & Environment, 1998, 21(7): 659- 669.
[11]Bunce J A. Stomatal conductance, photosynthesis and respiration of temperate deciduous tree seedlings grown outdoors at an elevated concentration of carbon dioxide. Plant, Cell & Environment, 1992, 15(5): 541- 549.
[12]Anderson P D, Tomlinson P T. Ontogeny affects response of northern red oak seedlings to elevated CO2and water stress: I. Carbon assimilation and biomass production. New Phytologist, 1998, 140(3): 477- 491.
[13]Tjoelker M G, Oleksyn J, Reich P B. Seedlings of five boreal tree species differ in acclimation of net photosynthesis to elevated CO2and temperature. Tree Physiology, 1998, 18(11): 715- 726.
[14]Roberntz P, Stockfors J. Effects of elevated CO2concentration and nutrition on net photosynthesis, stomatal conductance and needle respiration of field- grown Norway spruce trees. Tree Physiology, 1998, 18(4): 233- 241.
[15]王建林, 温学发. 气孔导度对CO2浓度变化的模拟及其生理机制. 生态学报, 2010, 30(17): 4815- 4820.
[16]陶汉之, 严子范. 茶树叶片蒸腾速率、气孔导度和水分利用率的研究. 安徽农学院学报, 1992, 19(1): 33- 38.
[17]申双和, 陶寅, 张方敏. 棉花阴、阳叶的气孔导度和光合作用的观测对比及模型应用. 南京气象学院学报, 2008, 31(4): 468- 472.
[18]柯世省, 魏燕, 陈贤田, 葛勇, 吴秀珍, 陶梦希. 云锦杜鹃气孔导度和蒸腾速率对水分的响应. 安徽农业科学, 2007, 35(21): 6363- 6369.
[19]成雪峰, 张风云, 柴守玺. 春小麦对不同灌水处理的气孔反应及其影响因子. 应用生态学报, 2010, 21(1): 36- 40.
[20]覃盈盈, 甘肖梅, 蒋潇潇, 李军伟, 韦锋, 梁士楚. 红树林生境中互花米草气孔导度的动态变化. 生态学杂志, 2009, 28(10): 1991- 1995.
[21]左应梅, 陈秋波, 邓权权, 唐建, 罗海伟, 巫铁凯, 杨重法. 土壤水分、光照和空气湿度对木薯气孔导度的影响. 生态学杂志, 2011, 30(4): 689- 693.
[22]卢振明, 牛文远, 张翼. 土壤含水量对冬小麦气孔导度开启程度的影响. 植物学报, 1986, 28(4): 419- 426.
[23]Pataki D E, Oren R, Katul G, Sigmon J. Canopy conductance ofPinustaeda,LiquidambarstyracifluaandQuercusphellosunder varying atmospheric and soil water conditions. Tree Physiology, 1998, 18(5): 307- 315.
[24]Running S W, Coughlan J C. A general model of forest ecosystem processes for regional applications I. Hydrologic balance, canopy gas exchange and primary production processes. Ecological Modelling, 1988, 42(2): 125- 154.
[25]Kelliher F M, Leuning R, Raupach M R, Schulze E D. Maximum conductances for evaporation from global vegetation types. Agricultural and Forest Meteorology, 1995, 73(1/2): 1- 16.
[26]朱丽薇, 赵平, 蔡锡安, 曾小平, 邹绿柳, 王权. 荷木人工林蒸腾与冠层气孔导度特征及对环境因子的响应. 热带亚热带植物学报, 2010, 18(6): 599- 606.
[27]赵平, 饶兴权, 马玲, 蔡锡安, 曾小平. 马占相思林冠层气孔导度对环境驱动因子的响应. 应用生态学报, 2006, 17(7): 1149- 1156.
[28]赵平, 马玲, 孙谷畴, 饶兴权, 蔡锡安, 曾小平. 利用基于sap flow测定值的冠层气孔导度和13C甄别率测定森林的碳同化率. 科学通报, 2006, 50(15): 1620- 1626.
[29]李仙岳, 杨培岭, 任树梅, 任亮. 樱桃冠层导度特征及模拟. 生态学报, 2010, 30(2): 300- 308.
[30]Jarvis P G. The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 1976, 273(927): 593- 610.
[31]Ball J T, Woodrow I E, Berry J A. A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions // Biggins J. Progress in Photosynthesis Research. Netherlands: Springer, 1987: 221- 224.
[32]Leuning R. A critical appraisal of a combined stomatal‐photosynthesis model for C3plants. Plant, Cell & Environment, 1995, 18(4): 339- 355.
[33]齐华, 于贵瑞, 刘允芬, 王建林. 柑橘叶片气孔导度的环境响应模型研究. 中国生态农业学报, 2004, 12(4): 43- 48.
[34]朱仲元, 朝伦巴根, 高志强, 高瑞忠. 基于Shuttleworth-Wallace双源模型的天然杨树蒸散量日变化研究. 水利学报, 2007, 38(5): 582- 590.
[35]Granier A, Loustau D, Bréda N. A generic model of forest canopy conductance dependent on climate, soil water availability and leaf area index. Annals of Forest Science, 2000, 57(8): 755- 765.
[36]孙林, 管伟, 王彦辉, 徐丽宏, 熊伟. 华北落叶松冠层平均气孔导度模拟及其对环境因子的响应. 生态学杂志, 2011, 30(10): 2122- 2128.
[37]韩磊. 黄土半干旱区主要造林树种蒸腾耗水及冠层蒸腾模拟研究[D]. 北京: 北京林业大学, 2011.
[38]Oguntunde P G, van de Giesen N, Savenije H H. Measurement and modelling of transpiration of a rain-fed citrus orchard under subhumid tropical conditions. Agricultural Water Management, 2007, 87(2): 200- 208.
[39]Lu P, Yunusa I A M, Walker R R, Müller W J. Regulation of canopy conductance and transpiration and their modelling in irrigated grapevines. Functional Plant Biology, 2003, 30(6): 689- 698.
[40]许文滔, 赵平, 王权, 饶兴权, 蔡锡安, 曾小平. 基于树干液流测定值的马占相思 (Acaciamangium) 冠层气孔导度计算及数值模拟. 生态学报, 2007, 27(10): 4122- 4131.
[41]苏永红, 朱高峰, 冯起, 常宗强, 司建华. 额济纳荒漠河岸胡杨林叶片气孔导度与微环境因子关系的模拟研究. 西北植物学报, 2008, 28(7): 1434- 1439.
[42]李红生, 刘广全, 陈存根, 王鸿喆, 徐怀同, 周海光. 黄土丘陵沟壑区沙棘光合特性及气孔导度的数值模拟. 西北农林科技大学学报: 自然科学版, 2009, 37(4): 108- 114.
[43]周莉, 周广胜, 贾庆宇, 吕国红, 谢艳兵, 赵先丽. 盘锦湿地芦苇叶片气孔导度的模拟. 气象与环境学报, 2006, 22(4): 42- 46.
[44]吴大千, 徐飞, 郭卫华, 王仁卿, 张治国. 中国北方城市常见绿化植物夏季气孔导度影响因素及模型比较. 生态学报, 2007, 27(10): 4141- 4148.
[45]魏征, 刘钰, 许迪, 蔡甲冰, 张宝忠. 基于叶片气孔导度提升的冬小麦冠层阻抗估算模型的应用和对比. 科学通报, 2013, 58(18): 1775- 1783.
[46]郑怀舟, 李机密, 黄儒珠, 王健, 朱锦懋. 长汀水土流失区马尾松与木荷气孔导度的模拟. 福建师范大学学报: 自然科学版, 2010, 26(5): 92- 96.
[47]王玉辉, 何兴元, 周广胜. 羊草叶片气孔导度特征及数值模拟. 应用生态学报, 2001, 12(4): 517- 521.
[48]唐凤德, 武耀祥, 韩士杰, 张军辉. 长白山阔叶红松林叶片气孔导度与环境因子的关系. 生态学报, 2008, 28(11): 5649- 5655.
[49]鱼腾飞, 冯起, 司建华. 极端干旱区多枝柽柳叶片气孔导度的环境响应模拟. 植物生态学报, 2012, 36(6): 483- 490.
[50]李永秀, 娄运生, 张富存. 冬小麦气孔导度模型的比较. 中国农业气象, 2011, 32(1): 106- 110.
[51]王治海, 刘建栋, 刘玲, 邬定荣, 毕建杰. 几种气孔导度模型在华北地区适应性研究. 中国农业气象, 2012, 33(3): 412- 416.
[52]杨再强, 黄川容, 费玉娟, 彭晓丹, 朱凯, 扎西才让. 基于Bp神经网络的温室番茄气孔导度的模拟研究. 东北农业大学学报, 2012, 42(11): 70- 77.
[53]米娜, 张玉书, 纪瑞鹏, 蔡福, 赵先丽, 张淑杰, 王宏博. 基于叶片尺度观测数据的气孔导度模型评价. 生态学杂志, 2011, 30(2): 389- 394.
[54]Wang S S, Yang Y, Trishchenko A P, Barr A G, Black T A, McCaughey H. Modeling the response of canopy stomatal conductance to humidity. Journal of Hydrometeorology, 2009, 10(2): 521- 532.
[55]石建红, 周锁铨, 余华, 孙善磊, 荆大为. 鄱阳湖流域典型树种夏季气孔导度模型及影响因素比较. 环境科学研究, 2010, 23(1): 33- 40.
[56]叶子飘, 于强. 植物气孔导度的机理模型. 植物生态学报, 2009, 33(4): 772- 782.
[57]Yu Q, Zhang Y Q, Liu Y F, Shi P L. Simulation of the stomatal conductance of winter wheat in response to light, temperature and CO2changes. Annals of Botany, 2004, 93(4): 435- 441.
[58]Medlyn B E, Duursma R A, Eamus D, Ellsworth D S, Prentice I C, Barton C V M, Crous K Y, de Angelis P, Freeman M, Wingate L. Reconciling the optimal and empirical approaches to modelling stomatal conductance. Global Change Biology, 2011, 17(6): 2134- 2144.
[59]傅伟, 王天铎. 一个气孔对环境因子响应的机理性数学模型. 植物生理学报, 1994, 20(3): 277- 284.
[60]王笑影, 李丽光, 谢艳兵, 李荣平, 李广霞, 周广胜. 植被-大气相互作用中的气孔导度及其尺度转换. 生态学杂志, 2008, 27(3): 454- 459.
[61]Leuning R. Modelling stomatal behaviour and photosynthesis ofEucalyptusgrandis. Australian Journal of Plant Physiology, 1990, 17(2): 159- 175.
[62]黄辉, 于贵瑞, 孙晓敏, 王秋凤, 赵风华, 李俊, 欧阳竹. 华北平原冬小麦冠层导度的环境响应及模拟. 生态学报, 2007, 27(12): 5209- 5221.
[63]Yu G R, Nakayama K, Matsuoka N, Kon H. A combination model for estimating stomatal conductance of maize (ZeamaysL.) leaves over a long term. Agricultural and Forest Meteorology, 1998, 92(1): 9- 28.
[64]曹庆平, 赵平, 倪广艳, 朱丽薇, 牛俊峰, 曾小平. 华南荷木林冠层气孔导度对水汽压亏缺的响应. 生态学杂志, 2013, 32(7): 1770- 1779.
[65]Jarvis P G, McNaughton K G. Stomatal control of transpiration: scaling up from leaf to region. Advances in Ecological Research, 1986, 15: 1- 49.
[66]刘薇, 陈翠英, 陈常梅, 刘晓帆. 基于 LAI 的 PM 蒸散发计算模型及应用. 人民黄河, 2012, 34(7): 33- 35.
[67]张宝忠, 刘钰, 许迪, 蔡甲冰, 赵娜娜. 基于夏玉米叶片气孔导度提升的冠层导度估算模型. 农业工程学报, 2011, 27(5): 80- 86.
[68]任传友, 于贵瑞, 王秋凤, 关德新. 冠层尺度的生态系统光合-蒸腾耦合模型研究. 中国科学D辑地球科学, 2004, 34(增刊II): 141- 151.
[69]Raupach M R. Vegetation-atmosphere interaction and surface conductance at leaf, canopy and regional scales. Agricultural and Forest Meteorology, 1995, 73(3/4): 151- 179.
[70]李小文. 地球表面时空多变要素的定量遥感项目综述. 地球科学进展, 2006, 21(8): 771- 780.
[71]Running S W, Nemani R R, Peterson D L, Band L E, Potts D F, Pierce L L, Spanner M A. Mapping regional forest evapotranspiration and photosynthesis by coupling satellite data with ecosystem simulation. Ecology, 1989, 70(4): 1090- 1101.
[72]Taconet O, Olioso A, Ben Mehrez M, Brisson N. Seasonal estimation of evaporation and stomatal conductance over a soybean field using surface IR temperatures. Agricultural and Forest Meteorology, 1995, 73(3/4): 321- 337.
[73]Sakai R K, Fitzjarrald D R, Moore K E. Detecting leaf area and surface resistance during transition seasons. Agricultural and Forest Meteorology, 1997, 84(3/4): 273- 284.
[74]Kalluri S N V, Townshend J R G, Doraiswamy P. A simple single layer model to estimate transpiration from vegetation using multi- spectral and meteorological data. International Journal of Remote Sensing, 1998, 19(6): 1037- 1053.
[75]Boegh E, Soegaard H, Thomsen A. Evaluating evapotranspiration rates and surface conditions using Landsat TM to estimate atmospheric resistance and surface resistance. Remote Sensing of Environment, 2002, 79(2/3): 329- 343.
[76]张佳华, 符淙斌, 王长耀. 利用遥感信息研究区域冬小麦气孔导度的时空分布. 气象学报, 2000, 58(3): 347- 353.
[77]辛晓洲, 田国良, 柳钦火. 地表蒸散定量遥感的研究进展. 遥感学报, 2003, 7(3): 233-240.
Environmental response simulation and the up-scaling of plant stomatal conductance
GAO Guanlong1,2, ZHANG Xiaoyou1,*, CHANG Zongqiang1, YU Tengfei1, ZHAO Hong3
1ColdandAridRegionsEnvironmentalandEngineeringResearchInstitute,ChineseAcademyofSciences,Lanzhou730000,China2ChineseAcademyofSciences,BeiJing100049,China3ZhejiangRuiqiTestingTECHCO.,Limited,Hangzhou310000,China
Abstract:Stomatal conductance is a key indicator of the balance and cycles of heat, H2O, and CO2 fluxes at the vegetation-atmosphere interface. Thus, it is essential to study the heat and H2O transfer in the soil, vegetation, and atmosphere andevaluate the status and role of vegetation, by considering the relationships between stomatal conductance and environmental factors, and by modeling and extrapolating the cumulative effect of stomatal conductance at the leaf, canopy, and regional levels. This paper summarized three aspects of research in this area: (1) the relationships between stomatal conductance and environmental factors at the leaf and canopy levels, (2) the simulation of stomatal conductance, and (3) the up-scalingof stomatal conductance from the leaf to canopy level and from the canopy to regional level. The results showed that the relationships between stomatal conductance and environmental factors were not consistent, because hereditary characteristics, environmental conditions, and time-scales varied; in addition, the comprehensive effects of environmental factors were not considered. A hypothesis was made concerning the mechanism underlying the relationship between the environmental factors and stomatal conductance. Systematic control and simultaneous observation should receive more attention when studying these combined influences. At present, Jarvis and BWB are the most commonly used stomatal conductance models. The suitability of these two models differed among research goals, study sites, and environmental conditions, so model selection should be carried out on a case-by-case basis. This paper also summarized other models. The results of up-scaling stomatal conductance from the leaf to canopy level and from canopy to the regional level showed that while research on the former is well established, the latter is still difficult to validate. Finally, we recommend that future research should focus on the following: (1) choosing the best model to simulate stomatal conductance, based on the environmental conditions; (2) considering interactions between environmental factors, and take into consideration their combined influence on stomatal conductance; (3) studying the coupling between the BWB and photosynthetic models; and (4) improving simulation accuracy at large scales.
Key Words:stomatal conductance; environmental response simulation; environmental factors; up-scaling
DOI:10.5846/stxb201408211652
*通讯作者
Corresponding author.E-mail: zhangxy@lzb.ac.cn
收稿日期:2014- 08- 21; 网络出版日期:2015- 07- 29
基金项目:国家自然科学基金项目(41271037); 国家自然科学基金青年科学基金项目(41401033); 中国科学院内陆河流域生态水文重点实验室(90Y290F41)
高冠龙, 张小由, 常宗强, 鱼腾飞, 赵虹.植物气孔导度的环境响应模拟及其尺度扩展.生态学报,2016,36(6):1491- 1500.
Gao G L, Zhang X Y, Chang Z Q, Yu T F, Zhao H.Environmental response simulation and the up-scaling of plant stomatal conductance.Acta Ecologica Sinica,2016,36(6):1491- 1500.
