小麦N离子束注入诱变育种的应用研究
2016-05-14李强任立凯陈凤孙中伟王康君郭明明
李强 任立凯 陈凤 孙中伟 王康君 郭明明
摘要:为了探寻新的育种手段,解决目前小麦常规育种中普遍存在的种质资源匮乏问题以提升育种效率,开展了小麦N离子束注入诱变育种与常规育种相结合的应用研究。以N离子不同剂量注入小麦品种连麦6号籽粒进行诱变处理,经过M1代损伤效应分析,M2代突变体的分类统计和筛选,M3代常规系谱法稳定性鉴定,考察了N离子注入对连麦6号农艺性状的诱变效应。结果表明:N离子注入对连麦6号的诱变效应在M1代显著地表现在株高、结实率、抽穗期和成株率上,M2代的突变率随N离子注入剂量的增加而提高,由低到高3种剂量处理的突变率分别为5.38%、8.98%和11.02%,筛选出的3个超亲变异性状(矮秆多蘖、早熟性、大穗)在M3代因不同的剂量处理其重复稳定性存在明显差异;试验明确了不同剂量的离子注入处理对小麦生理性状影响的变异效应不同。
关键词:小麦;N离子束;诱变育种
中图分类号:S512.103.52文献标识号:A文章编号:1001-4942(2016)07-0018-05
黄淮麦区是我国小麦的主产区,多年的常规杂交育种致使小麦育种的遗传背景变得单一,品种的同质化现象严重。实践中,借助远缘杂交来打破此瓶颈,又遇属间不育性障碍且很难突破。诱变育种技术在种质创新和品种改良上具有独特作用,它可以诱发基因突变,打破基因连锁,在相对较短的世代周期内获得常规育种方法难以获得的新类型、新性状、新基因,可以丰富育种材料的遗传背景。在众多诱变手段中离子束的诱变作为后起之秀倍受关注。这不仅是因为离子束生物工程的应用发端于我国[1,2],更是因为它不象其它辐射诱变只限于能量作用引起 DNA链断裂,更有其能量、质量、电荷的三因子协同作用产生的生物分子原子移位、重组和化合的优势[3],具有损伤轻、存活率高、突变率高和突变谱广、重复性好等特点[4~6]。
目前离子束诱变育种研究正在国内外蓬勃展开,涉及方面从突变体库的选择构建、品性改良、种质创新和离子束介导转基因[7~12]到以优化离子束诱变效应的诱变离子、剂量、时间及不同诱变靶标材料的对应选择方面[13~17]。从前人的这些研究成果看,离子束诱变边际效应最大化的受制因素往往与靶标材料密切相关。因此要获得好的诱变效率,诱变源的参数最佳组合必须以诱变材料为转移,不能教条地套用。为了提高处理后代的突变频率和突变类型,通常诱变育种选用对本地区生态环境适应性好、综合性状优良、遗传背景丰富且某些性状尚存在改良余地的品种为主。本院自育品种连麦6号从亲本遗传背景和生产生态适应性看大致符合这些条件,为此我们选择该品种开展了小麦N离子束注入诱变育种应用研究,试图找到一套适合本地品种和生态条件的与常规育种相得益彰互为补充的育种方法。
1材料与方法
1.1供试材料
供试小麦品种连麦6号为本院自产。
1.2辐照处理
离子注入委托中囯科学院合肥物质科学研究院技术生物与农业工程研究所完成。注入离子为N离子,能量为30 keV,束流大小为10 mA,真空度为10-2~10-3 Pa,注入剂量分别为2×1017 、4×1017、6×1017 N+/cm2。每个离子注入剂量处理均为1 000粒干种子。
1.3田间试验方法
试验在连云港市农业科学院玉带河试验场进行。2012年11月5日播种M1代,行距 22.5 cm,株距2 cm,每小区播种500粒,重复2次。苗期每小区定点调查田间发芽势、发芽率和成活率,考察不同剂量处理对连麦6号种子的损伤情况,同时单株挂牌跟踪调查生育期。收获前每小区随机取样20株室内考种。M1代成熟时不做选择,按处理单收主茎穗。
2013年10月15日对上年收获的单株主茎穗按不同诱变处理单株单行种植M2代,每单株行长1 m, 每隔10行种1行野生型连麦6号为对照,以便于对比观察。调查各处理M2代群体的生育期、株高、分蘖、穗长等农艺性状,观察记载其它性状(株、粒、叶的形状、颜色等)变异,以超野生型连麦6号(CK)不同数量性状平均值3倍标准差之外的个体作为突变株,统计各处理不同类型的突变率,同时按处理根据小麦不同生育进程有所侧重地分茎秆、熟性、穗部三大类变异性状进行株选淘汰,保留性状向优突变的变异株。
2014年10月20日再对中选的变异株M2代按不同的离子注入剂量处理依常规育种系谱法建立株系谱,针对所筛选的变异性状田间观察和室内考种对M3代每个株系进行稳定性重复鉴定,选择性状向优突变且遗传特性相对稳定的材料。
2结果与分析
2.1N离子束不同注入剂量对连麦6号M1代的影响
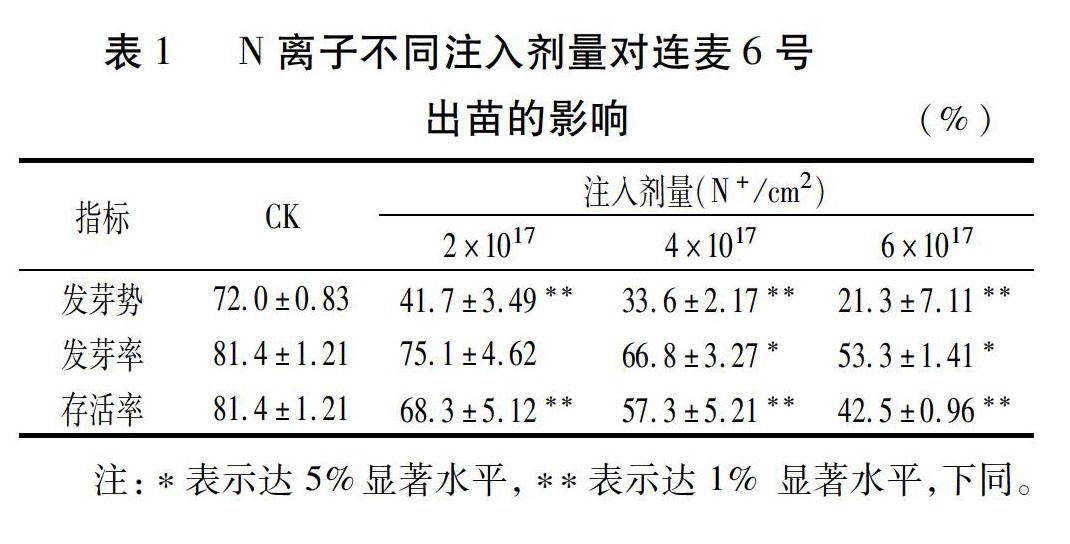
考虑传统的物理诱变的遗传通常表现为一代平、二代突、三代稳的特点,所以在种植M1代时特意采用了晚播密植的方法,以尽可能降低分蘖数,利于集中突显M1代主茎的诱变表达。从表1中可以看出,N离子的注入对连麦6号的损伤很明显,M1代不仅出现了发芽势弱的情况,而且发芽率和存活率亦出现双低现象,三项指标与CK的差异几乎均达显著或极显著水平。从存活率指标看,3个N离子注入剂量处理的存活率2个超过57%,1个接近50%,说明选择2×1017~6×1017 N+/cm2这个注入诱变剂量范围对连麦6号是适宜的。从田间对比观察看到,M1代也出现了一些诸如叶宽色黄、抽穗不一、穗子畸形、高矮不齐等现象,这显然也是离子注入致损的表现。
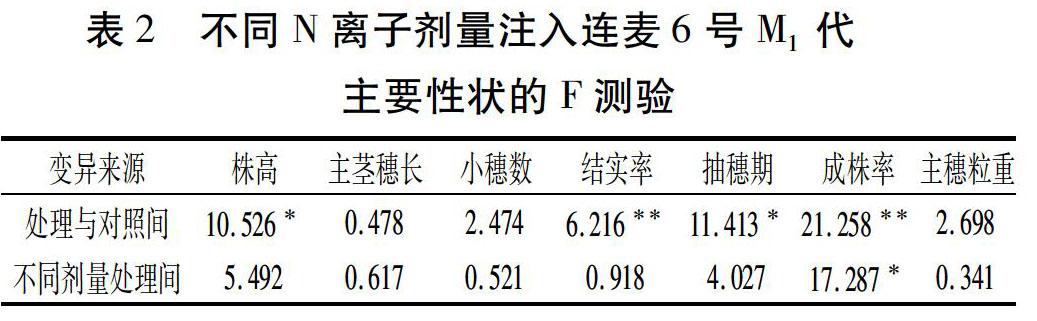
为了更科学地作出评估判断,进一步对大量的室内考种数据和田间观察数据作统计分析,结果见表2。可以看出,凡经N离子注入处理的株高、结实率、抽穗期、成株率与CK差异达极显著或显著水平,而其它三项穗部性状表现差异不显著。再从不同剂量处理间的比较看,除了成株率的差异达显著外,其余的性状指标均不显著。 由此表明与其它的诱变源相比较,N离子束的诱变效果在M1代就得到突现,而且这种表现是随注入剂量增加而增大。但考虑M1代植株在遗传上一般为复杂的突变嵌合体,其损伤效应出现的变异一般不能传递给后代,所以只收主茎穗而不作选择。
2.2N离子束不同注入剂量处理连麦6号M2代的变异表现
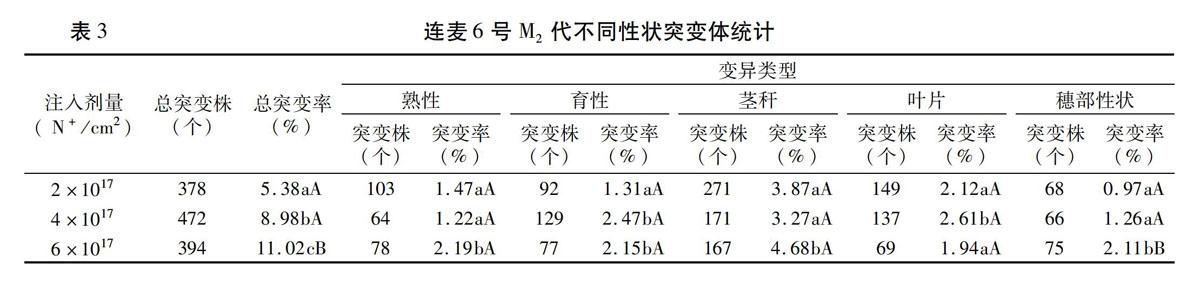
3个离子注入剂量处理由低到高分别收获了593、486、395个主茎穗。M2代是诱变育种选择的关键世代,为便于观察甄别M2代的变异分离,对M1代主茎穗没有混收混种,而是严格按不同N离子注入剂量处理建立起同源穗行种植圃。田间观察,连麦6号M2代的变异分化非常丰富,不同剂量处理的1 474个穗行的15 857个单株中共有1 244 株出现一到多个性状的突变,总表型突变率达7.85%。由表3可以看出,连麦6号M2代的突变率随N离子注入剂量的增加而提高,由低到高 3种剂量处理的突变率分别为5.38%、8.98%和11.02%,多重比较差异显著。这种趋势显然也与M1代的损伤效应相吻合。细分变异类型的差异还发现,突变群体中以茎秆的突变频率最高,其次为叶片、育性和熟性的变异,而穗部性状的突变频率相对较低。进一步纵向比较处理间的突变差异,高剂量的穗部、茎秆和熟性突变率显著高于中低剂量的突变,中剂量的叶片突变又显著高于高剂量的突变。这表明对不同性状的选择存在着一个与之对应的最佳注入诱变剂量。综观1 244个M2代变异株,负向弱亲变异的有1 126株,占比高达90.5%,正向超亲变异的占比很低,而且还多是正负变异性状混搭株,仅占9.5%。
2.3N离子束不同注入剂量处理的连麦6号M3代的稳定性表现
按矮秆多蘖、早熟性、大穗三种突变类型从M2后代中选出相符的变异突变株57株,其中涉及茎秆性状的24株、穗部性状的18株、熟性的15株。按M2的种植方法建立株系圃,分别考量鉴定熟性、茎秆和穗部性状变异的遗传稳定性。由表4可见,选出的3个超亲变异性状在不同剂量处理M3代上表现的重复稳定性存在明显差异。不同处理早熟性变异株抽穗期比CK提前1~3天,中低剂量处理的差异显著;矮秆变异性状稳定性突出,3个剂量处理的平均株高分别比CK矮14.0、8.5 cm和11.7 cm,差异达显著或极显著水平;分蘖变异性状,低剂量处理的统计差异显著,单株平均分蘖比CK增2.6个,另两个处理虽也表现增加,但不显著;穗部变异性状的稳定性主要测定了主茎穗的粒数、穗长和粒重3个指标,从不同剂量处理的大穗突变株与之对应的测定数据看,中高剂量处理的大穗突变株的大穗特性得到保持,3个测定指标都显著高于CK。综上可见,从M2代筛选的3个超亲突变性状都基本得到保持,但各性状尚存在较大的变幅,说明这些性状在M3代的稳定性还不够,还须进一步作世代稳定性筛选。
3讨论与结论
试验完整观察统计分析了N离子束不同剂量处理的连麦6号3个世代的农艺性状表型变异情况,可以看出,3种剂量的N离子束注入对连麦6号田间出苗的损伤随注入剂量的增加而加剧,但并未呈现“马鞍形”的存活率剂量效应曲线[18]。这恰恰反映出不同小麦品种由于基因型的差异对注入离子剂量的耐受力和敏感性的差异。统计分析表明低能N离子束注入对连麦6号多项生物学指标产生的影响在M1代就有显著的表现。这与以往认为诱变处理产生的M1代突变大多为隐性突变的现象有较大的出入,这种差异可能与注入的有生物活性的N离子与生物分子作用很强有关。但为避免M1代可能出现的突变嵌合体的干挠导致变异的丢失,所以收获时未对其进行选择。
试验再次证明,M2代是诱变分离最大、出现变异类型最多的一个世代,3个剂量处理的总突变率为7.85%。就连麦6号而言,突变率是随N离子注入剂量的增加而提高,试验设定条件下2×1017~6×1017 N+/cm2的剂量范围内M1代的损伤程度愈重M2代的突变率愈高。总体上看突变群体中以茎秆的突变频率最高,其次为叶片、育性和熟性的变异,而穗部性状的突变频率相对较低。6×1017 N+/cm2注入剂量的诱变效率最好,其总突变率达11.02%,细分的茎秆、熟性以及穗部性状的突变率都显著高于其它剂量处理。值得注意的是:获得的1 244个M2代变异株中负向弱亲变异占绝对多数,正向超亲变异的早熟、矮秆、优良株型等表形突变以及穗粒数、粒重等微突变的占比相对较低,可供育种选择的正向超亲变异或混合型变异株仅有9.5%。但本试验中不同剂量间表现出的诱变差异还是给我们留下了对离子束诱变小麦性状变异可调控的努力空间。
比较同源株系圃间的性状稳定性差异,一些超亲的变异性状在M3代上得到保持。2×1017 N+/cm2处理的矮秆多蘖突变性状保持更好,而4×1017 N+/cm2和6×1017 N+/cm2注入剂量的大穗突变株的大穗特性得到保持。这表明,尽管离子束的诱变总体上表现为随机性,但依作物品种和性状选择而定的不同诱变剂量还是相对能找到一个与之相对应的最适选择。从试验中还可看出,所选的变异性状在M3代虽得以保持,但还存在较大变幅,而且大部分还是优良互掺的混合型变异,这就需要紧扣育种目标进一步与常规育种相结合对其世代进行稳定或杂交改造。通过N离子束诱变可在相对短的时间内获得连麦6号的一些超亲突变体,这不仅大大丰富了育种材料,也能提高育种效率。本试验表明不同剂量的离子注入处理对小麦生理性状影响的变异效应不同,如何进一步提高小麦N离子束诱变效率,诱发筛选出符合育种目标的突变类型,丰富种质资源,则是今后需要深化研究的方面。
参考文献:
[1]余增亮,何建军,邓建国,等.离子注入水稻诱变育种机理初探[J].安徽农业科学,1989,17(1):12-16.
[2]余增亮.离子束与生命科学——一个新的研究领域[J].物理,1997,26(6):333-338.
[3]余增亮.离子束生物技术引论[M].合肥:安徽科学技术出版社,1998.
[4]卫增泉.重离子生物效应研究中的一些基本物理问题[J]. 核物理动态,1996,13(1):40-46.
[5]陈恒雷,吕杰,曾宪贤.离子束诱变育种研究及应用进展[J].生物技术通报,2005,24(2):45-47.
[6]黄国伟,毛培宏,金湘,等.低能离子注入介导外源DNA转化的原理与应用[J].基因组学与应用生物学,2009,28(4):815-820.
[6]甘斌杰,杨赞林,余增亮,等.离子束诱变小麦新品种皖麦42号的特征特性和栽培技术[J].中国农学通报,2003,19(5):25-27,36.
[7]许学,刘斌美,宋美,等. N离子束与γ射线辐照日本晴和“9311”水稻突变体库的筛选[J]. 核农学报,2008,22(4):389-393.
[8]靳文奎,廖平安,郭春强,等. 离子束诱变对小麦后代湿面筋含量变化的影响[J].安徽农业科学,2005,33(6):962.
[9]焦浈,史艳芹,秦广雍.离子束诱变对M4代小麦醇溶蛋白和氨基酸组分的影响[J]. 河南农业科学,2009(6):25-27.
[10]赵连芝,王浩瀚,王勇,等. 重离子辐照选育春小麦新品种初探[J]. 西北农业学报,2006,15(3):17-19.
[11]陈秀兰,柳学采.利用离子注入诱变创造小麦新种质[J].江苏农学院学报,1998,19(2):62-64.
[12]吴丽芳,李红,宋道君,等.建立低能离子束介导小麦转基因方法并获得转GUS基因植株[J].遗传学报,2000,27(11):982-991.
[13]郭向萌,押辉远. 离子辐射诱变小麦育种研究进展[J]. 安徽农业科学,2011,39(11):6334-6335,6410.
[14]程备久,朱苏文,李培金,等.不同离子辐照对离体质粒DNA损伤与转化活性的影响[J]. 激光生物学报,2001,10(1):40-43.
[15]薛建明,王宇钢,卢希庭,等.低剂量离子在物体样品中透射行为的研究[J]. 核技术,1999,22(8):449-453.
[16]李兴林,卫增泉,王晓娟,等.12C6+注入小麦种子胚乳引起M1代的变化[J]. 激光生物学报,2000,9(4):276-280.
[17]卫增泉,颉红梅,韩光武,等.110keV 56Fe1+离子注入麦胚中的能量沉积分布[J].安徽农业大学学报,1994,21(3):250-254.
[18]宋道军,姚建铭,邵春林,等. 离子注入微生物产生“马鞍形”存活曲线的可能作用机制[J]. 核技术,1999,22(3):129-132.
