中国伞形科棱子芹属果实微形态及其分类学研究
2016-05-10魏雪莹刘玫茹剑王宇婷王超群郭诗琪
魏雪莹,刘玫,茹剑,王宇婷,王超群,郭诗琪
(哈尔滨师范大学生命科学与技术学院,黑龙江省普通高等学校植物生物学重点实验室,黑龙江 哈尔滨 150025)
中国伞形科棱子芹属果实微形态及其分类学研究
魏雪莹,刘玫*,茹剑,王宇婷,王超群,郭诗琪
(哈尔滨师范大学生命科学与技术学院,黑龙江省普通高等学校植物生物学重点实验室,黑龙江 哈尔滨 150025)
摘要:本文利用光镜首次深入研究了中国伞形科棱子芹属21种植物果实的微形态(包括翅,外果皮,中果皮,维管束,油管,结合面,胚乳及结晶)。结果显示,棱子芹属不同物种果实结构变化较大,可分为3组:组1(喜马拉雅棱子芹等3种)和组2(归叶棱子芹等7种)翅非波状,外果皮细胞外壁平滑,与中果皮不易分离,但球状结晶仅存在于组1。组3(松潘棱子芹等11种)翅缘波状,外果皮具瘤突,且细胞外壁向外突起,易与中果皮分离,中果皮无结晶。果实微形态为分子系统学揭示的棱子芹属是复系的,喜马拉雅棱子芹和云南棱子芹亲缘关系较近,松潘棱子芹及翼叶棱子芹等物种应移入棱子芹族提供了形态学依据,并为进一步研究世界棱子芹属及其相关类群提供了形态学基础。
关键词:棱子芹属;果实;翅;伞形科;分类学
棱子芹属(Pleurospermum)隶属于伞形科(Apiaceae),芹亚科(Apioideae),美味芹族(Smyrnieae)。约50种,分布于欧洲及亚洲,尤其喜马拉雅地区为多[1-3]。中国有39种,其中22种为特有种[3]。此属曾被认为是伞形科最有问题的属之一[1,4-8]。Pimenov和Kljuykov[9]研究的狭义棱子芹属(Pleurospermumssensustricto)只包括2种,Pleurospermumaustriacum和棱子芹(P.uralense),这2种形态相近,有时被作为1种的2亚种[10]。位于美味芹族的另外4属,沟芹属(Aulacospermum),藏香芹属(Hymenidium),膜苞芹属(Hymenolaena)及圆翅芹属(Pterocyclus)曾被作为棱子芹属的异名[3,9]。
分子系统学研究表明棱子芹属为一复系类群,喜马拉雅棱子芹(Pleurospermumhookeri)和云南棱子芹(P.yunnanense)构成一个分支,而另一个分支包括松潘棱子芹(P.franchetianum),翼叶棱子芹(P.decurrens),丽江棱子芹(P.foetens)及棱子芹(P.uralense)[11]。Zhou等[12-13]的分子系统学研究同样证明棱子芹属是复系的,其中松潘棱子芹,瘤果棱子芹(P.wrightianum)及粗茎棱子芹(P.wilsonii)位于Downie等[14]建立的棱子芹族(Pleurospermeae),而喜马拉雅棱子芹(P.hookeri)和云南棱子芹位于丝瓣芹属分支(Acronemaclade)[12-13]。
伞形科果实变异丰富且结构稳定,是分类的重要依据。其翅,油管,内果皮,心皮柄及结晶等特征被用于亚科,族及属的分类[1,15-17]。近年来伞形科果实微形态的研究,特别是从三维结构的角度揭示果实特征,为分子系统学的研究提供了重要的形态学依据[18-22]。已有的关于棱子芹属果实的描述多限于其形状及油管数目[3,23-25],果实的许多解剖结构不清楚。本文旨在通过进一步深入研究我国棱子芹果实的微形态,完善其属的形态学,为分子系统学的研究提供形态学依据,并为进一步研究世界棱子芹属提供了形态学基础。
1材料与方法
1.1材料
本文所用的研究材料为中国棱子芹属21种植物的成熟果实,物种名称及凭证标本信息见表1。材料于2014年分别取自于中国科学院成都生物研究所植物标本馆(CDBI-Herbarium, Chengdu Biology Institute),中国科学院沈阳应用生态研究所标本馆(IFP-Institute of Applied Ecology, CAS),中国科学院昆明植物研究所标本馆 (KUN-Herbarium, Kunming Institute of Botany, CAS),中国科学院植物研究所标本馆(PE-Herbarium, Institute of Botany, CAS)及中国科学院新疆生态与地理研究所标本馆(XJBI-Herbarium, Xinjiang Institute of Ecology and Geography, CAS)。
1.2方法
分离果皮:将干燥的果实(每物种取2~3个)放入热水中(约90℃)浸泡,待材料完全吸水膨胀并沉入水底时(约2 h),将其取出。用解剖镜(Olympus SP-350)观察果皮外部形态,然后分离果皮,观察中果皮内分泌结构的特征。之后,将带有结晶的部分中果皮放在载玻片上,加1~2滴50%甘油水溶液后盖上盖玻片。用具成像系统(Olympus DP 70)的显微镜(Olympus BX 51)观察结晶的形态及分布,并照相。
GMA(glycol methacrylate,乙二醇甲基丙烯酸酯)切片:根据Feder和O’Brien[26]的方法,每个物种取2~3个果实,经吸水膨胀后放入FAA(37% formalin-glacial acetic acid-70% alcohol,福尔马林-冰醋酸-酒精=5 mL∶5 mL∶90 mL)固定,时间不少于24 h。材料经50%酒精(4~6 h),100%酒精(4~6 h),异丙醇(6 h)和正丁醇(6 h)脱水,每步各2次。材料进入GMA渗透3次,第1和2次各1 d,第3次不少于5 d。之后,将材料及GMA装入无色胶囊,并置于60℃温箱聚合(24 h)。用Leica Ultracut R切片机切片,厚度3~4 μm。同样根据Feder和O’Brien[26]的方法,将切片用甲苯胺蓝染色,中性树胶封片。
2结果与分析
2.1果实结构
本文详细地描述了中国棱子芹属果实结构,包括翅(wing),外果皮(exocarp),中果皮(mesocarp),维管束(vascular bundle),伴生油管及非伴生油管(rib duct and vitta),结合面(commissure),胚乳(endosperm) 及结晶(crystal)(图1~3),主要的分类特征见表2。

表1 用于研究果实特征的中国棱子芹属物种的凭证标本信息及采集地

图1 中国棱子芹属分果,示分果外形,翅,油管及心皮柄
A:美丽棱子芹P.amabile;B:归叶棱子芹P.angelicoides;C:雅江棱子芹P.astrantioideum;D:宝兴棱子芹P.benthamii;E:二色棱子芹P.bicolor;F:鸡冠棱子芹P.cristatum;G:翼叶棱子芹P.decurrens;H:丽江棱子芹P.foetens;I:松潘棱子芹P.franchetianum;J:垫状棱子芹P.hedinii;K:芷叶棱子芹P.heracleifolium;L:喜马拉雅棱子芹P.hookeri;M:长果棱子芹P.longicarpum;N:疏毛棱子芹P.pilosum;O:青藏棱子芹P.pulszkyi;P:心叶棱子芹P.rivulorum;Q:单茎棱子芹P.simplex;R:青海棱子芹P.szechenyi;S:棱子芹P.uralense;T:瘤果棱子芹P.wrightianum;U:云南棱子芹P.yunnanense. c:心皮柄Carpophore;lr (lw):侧脊(侧翅)Lateral rib (lateral wing);mar (maw):边脊(边翅)Marginal rib (marginal wing);mer (mew):中脊(中翅)Median rib (median wing);vv:沟中油管Vallecular vitta. 标尺Scale bar:2 mm.
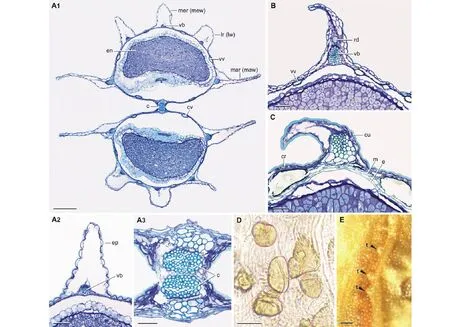
图2 中国棱子芹属果实横切面(A~C),中果皮(D)及外果皮(E),示果皮表面,脊,翅,维管束,分泌油管,结合面,胚乳,心皮柄及结晶Fig.2 Transverse sections of fruits (A-C), mesocarp (D) and exocarp (E) of Pleurospermum of China, showing variation in pericarp surface, ribs, wings, vascular bundle, secretory ducts, pericarps, endosperms, commisure sides, carpophores, and crystals A1~A3:宝兴棱子芹P. benthamii;B:二色棱子芹P. bicolor;C, D:美丽棱子芹P. amabile;E:瘤果棱子芹P. wrightianum.c:心皮柄Carpophore;cr:结晶Crystal;cu:角质层Cuticle;cv:结合面油管Commissural vitta;e:内果皮Endocarp;en:胚乳Endosperm;ep:表皮Epidermis;lr (lw):侧脊(侧翅)Lateral rib (lateral wing);m:中果皮Mesocarp;mar (maw):边脊(边翅)Marginal rib (marginal wing);mer (mew):中脊(中翅)Median rib (median wing);rd:伴生油管Rib duct;t:瘤状突起Tuber;vb:维管束Vascular bundle;vv:沟中油管Vallecular vita;A1的标尺Scale bar in A1:0.5 mm;其他的标尺Scale bar in the others:0.1 mm.
果实形状、脊及翅:棱子芹属果实的两分果均为背腹压扁(图2A1)。其中一些物种的果实较短(如,美丽棱子芹和瘤果棱子芹),而另一些果实明显较长(如,归叶棱子芹及长果棱子芹)。分果长宽比为1.3∶1~3.5∶1。分果具5个脊,即中脊、侧脊及边脊(图2A1,图3B),脊处的外果皮及中果皮向外延伸形成翅(图2A1),即中翅,侧翅及边翅(图3A,图3B)。翅较薄,存在于研究的全部物种(图1A~U,图3A~U)。翅的边缘为平滑,如美丽棱子芹,归叶棱子芹及芷叶棱子芹等物种(图1A,B,K);呈深波状,如雅江棱子芹,鸡冠棱子芹及青海棱子芹(图1C,F,R);或浅波状,如宝兴棱子芹,二色棱子芹及单茎棱子芹(图1D,E,Q)。多数物种的5个翅近等宽,如归叶棱子芹,二色棱子芹及翼叶棱子芹(图3B,E,G),少数物种的边翅宽于其中翅及侧翅,如雅江棱子芹,鸡冠棱子芹及松潘棱子芹(图3C,F,I)。边翅较宽,为分果宽度的40%~60%,如疏毛棱子芹,单茎棱子芹及瘤果棱子芹(图3N,Q,T),或相对较窄,为分果宽度的30%~40%,如美丽棱子芹,喜马拉雅棱子芹及长果棱子芹(图3A,L,M)。

图3 中国棱子芹属分果的横切面,示脊,翅,维管束,分泌油管,果皮,结合面,胚乳及心皮柄Fig. 3 Transverse sections of Pleurospermum mericarps of China, showing variation in ribs, wings, vascular bundle, secretory ducts, pericarps, endosperms, commisure sides, and carpophores A:美丽棱子芹P. amabile;B:归叶棱子芹P. angelicoides;C:雅江棱子芹P. astrantioideum;D:宝兴棱子芹P. benthamii;E:二色棱子芹P. bicolor;F:鸡冠棱子芹P. cristatum;G:翼叶棱子芹P. decurrens;H:丽江棱子芹P. foetens;I:松潘棱子芹P. franchetianum;J:垫状棱子芹P. hedinii;K:芷叶棱子芹P. heracleifolium;L:喜马拉雅棱子芹P. hookeri;M:长果棱子芹P. longicarpum;N:疏毛棱子芹P. pilosum;O:青藏棱子芹P. pulszkyi;P:心叶棱子芹P. rivulorum;Q:单茎棱子芹P. simplex;R:青海棱子芹P. szechenyi;S:棱子芹P. uralense;T:瘤果棱子芹P. wrightianum;U:云南棱子芹P. yunnanense. c:心皮柄Carpophore;co:结合面Commissure;cv:结合面油管Commissural vitta;e:内果皮Endocarp;en:胚乳Endosperm;lr (lw):侧脊(侧翅)Lateral rib (lateral wing);m:中果皮Mesocarp;mar(maw):边脊(边翅)Marginal rib (marginal wing);mer (mew):中脊(中翅)Median rib (median wing);rd:伴生(或脊中)油管Rib duct;vb:维管束Vascular bundle;vv:沟中油管Vallecular vitta. 标尺Scale bar:1 mm.
果皮:多数物种的外果皮(即表皮)平滑,如归叶棱子芹,长果棱子芹及青藏棱子芹(图1B,M,O)。少数物种外果皮的一些相邻细胞共同向外突起,形成瘤状,如青海棱子芹,棱子芹及瘤果棱子芹(图2E)。一些物种外果皮细胞的外壁明显向外突起,如宝兴棱子芹,鸡冠棱子芹及丽江棱子芹(图2A2),而有些物种的外壁平直,如二色棱子芹,心叶棱子芹及云南棱子芹(图2B)。外果皮细胞外壁通常具较厚的角质层(图2C)。中果皮细胞为2~5层,内果皮为1层细胞(图2C)。中果皮及内果皮细胞均轻微木化,如美丽棱子芹,雅江棱子芹及翼叶棱子芹(图2C),或仅由薄壁细胞组成,如归叶棱子芹,二色棱子芹及瘤果棱子芹(图2B)。
维管束:每个分果具5个维管束,分别位于分果的中脊,侧脊及边脊(图2A1)。所有研究的物种维管束均较小,由少数木化细胞构成,位于翅的基部,即靠近内果皮并围绕着种子(图2A2,图3A~U)。
伴生油管:伴生油管是与维管束相伴的油管,存在于分果脊内维管束的外侧(图2B)。在所研究物种中伴生油管只存在于二色棱子芹,翼叶棱子芹,丽江棱子芹及棱子芹(图3E,G,H,S),其他物种均无伴生油管。伴生油管的腔小,他们存在于5个脊内(图3S),或仅与少数维管束相伴(图3E,G,H)。

非伴生油管:非伴生油管简称油管,不与维管束相伴。他们存在于果实的沟里,即两个脊之间的中果皮,及结合面(图1M,图2A1,B)。多数物种果实的沟里仅具1个油管,结合面2个油管,如长果棱子芹,青海棱子芹及瘤果棱子芹(图3M,R,T),少数物种沟里油管多于1个(多至4个),结合面油管多于2个(多至7个),这些物种包括美丽棱子芹,二色棱子芹,喜马拉雅棱子芹,青藏棱子芹,单茎棱子芹及云南棱子芹(图3A,E,L,O,Q,U)。
胚乳:胚乳在结合面平直,如宝兴棱子芹,松潘棱子芹及喜马拉雅棱子芹(图3D,I,L),微凹,如归叶棱子芹,雅江棱子芹及二色棱子芹(图3B,C,E),或深凹,如美丽棱子芹,鸡冠棱子芹及芷叶棱子芹(图3A,F,K)。
结合面及心皮柄:结合面为两分果的相接合处(图3P)。所研究的物种的结合面通常较窄,美丽棱子芹及长果棱子芹的结合面宽占分果宽的10%,其他物种的结合面为5%。所有物种均具两个较细的,彼此相对心皮柄(图1D2,图2A3,图3A~U)。
结晶:大量的球状结晶存在于美丽棱子芹,喜马拉雅棱子芹及云南棱子芹的靠近外果皮的中果皮(图2C,D),而其他物种的中果皮无结晶。
2.2检索表
根据21种棱子芹果实的主要特征(包括翅,外果皮,中果皮,维管束,油管,结合面,胚乳及结晶),制作检索表如下:
1.中果皮具球状结晶(组1)
2.结合面油管数量>5个
3.结合面占果宽10%,腹面胚乳深凹
美丽棱子芹P.amabile
3.结合面占果宽5%,腹面胚乳微凹
云南棱子芹P.yunnanense
2.结合面油管数量≤5个
喜马拉雅棱子芹P.hookeri
1.中果皮无球状结晶
4.翅非波状(组2)
5.结合面占果宽10%
长果棱子芹P.longicarpum
5.结合面占果宽5%
6.腹面胚乳平直
心叶棱子芹P.rivulorum
6.腹面胚乳凹陷
7.中果皮及内果皮木化
青藏棱子芹P.pulszkyi
7.中果皮及内果皮不木化
8.腹面胚乳深凹
芷叶棱子芹P.heracleifolium
8.腹面胚乳微凹
9.结合面油管6
二色棱子芹P.bicolor
9.结合面油管<6
10.边翅占果宽50%
疏毛棱子芹P.pilosum
10.边翅占果宽40%
归叶棱子芹P.angelicoides
4.翅缘波状(组3)
11.边翅宽于中翅及侧翅
12.翅缘浅波状
13.表皮具瘤状突起
宝兴棱子芹P.benthamii
13.表皮不具瘤状突起
松潘棱子芹P.franchetianum
12.翅缘深波状
14.边翅占果宽30%
鸡冠棱子芹P.cristatum
14.边翅占果宽50%
15.有伴生油管
丽江棱子芹P.foetens
15.无伴生油管
雅江棱子芹P.astrantioideum
11.边翅不宽于中翅及侧翅
16.翅缘深波状
17.中果皮及内果皮木化
青海棱子芹P.szechenyi
17.中果皮及内果皮不木化
瘤果棱子芹P.wrightianum
16.翅缘浅波状
18.边翅占果40%
棱子芹P.uralense
18.边翅占果>40%
19.有伴生油管
翼叶棱子芹P.decurrens
19.无半生油管
20.腹面胚乳微凹
垫状棱子芹P.hedinii
20.腹面胚乳深凹
单茎棱子芹P.simplex
3讨论
所研究的棱子芹属物种的果实均为背腹压扁,每个分果具5个较薄的翅,果实具两个较细的心皮柄,然而根据这个属果皮及细胞内含物的特征可将研究的21种棱子芹分为3组。
组1:翅非波状,外果皮细胞外壁平直,与中果皮不易分离,中果皮有结晶。这个组包括喜马拉雅棱子芹,云南棱子芹及美丽棱子芹。蒲吉霞[11]的细胞学研究指出喜马拉雅棱子芹及云南棱子芹与其他棱子芹不同,其染色体数目为2n=22。Zhou等[12-13]的分子系统学研究显示喜马拉雅棱子芹及云南棱子芹位于丝瓣芹属分支,而这一分支还包括丝瓣芹属(Acronema),囊瓣芹属(Pternopetalum),滇芹属(Meeboldia),芹属(Apium),藳木属(Ligusticum)及小芹属(Sinocarum)。为了探讨这些属与喜马拉雅棱子芹,云南棱子芹及美丽棱子芹的关系,我们观察了位于这些属的一些物种,如丝瓣芹属的星叶丝瓣芹(Acronemaastrantiifolium),滇芹属的滇芹(Meeboldiayunnanensis),囊瓣芹属的澜沧囊瓣芹(Pternopetalumdavidi),小芹属的紫茎小芹(Sinocarumcoloratum)果实的中果皮,发现他们均具有同喜马拉雅棱子芹等一样的球状晶体。大量的研究证明果实中结晶的结构在伞形科分类中起着重要的作用,如深棕亚科(Mackinlayoideae)及牵环花亚科(Azorelloideae)具单晶和晶簇[1,15-16,20],变豆菜亚科(Saniculoideae)以及相关类群(如Polemanniopsis及Steganotaenia)仅具晶簇[18-19]。我们的研究显示尽管喜马拉雅棱子芹及云南棱子芹的一些特征(如外果皮细胞平直,不易同中果皮分离)也存在于棱子芹属的一些其他物种中,但球状结晶是这两个种特有的,因而支持分子系统学显示的二者亲缘关系较近,同时也为分子系统学将二者位于丝瓣芹属分支提供了形态学依据。由于在研究的21种棱子芹中,美丽棱子芹的晶体(球状晶体)及油管(沟里油管多于1个,结合面油管多于2个)特征同喜马拉雅棱子芹及云南棱子芹相同,故作者认为这3种棱子芹应具有较近的亲缘关系。
组2:翅非波状,外果皮细胞平滑,与中果皮不易分离,中果皮无结晶。这个组包括归叶棱子芹,二色棱子芹,芷叶棱子芹,长果棱子芹,疏毛棱子芹,青藏棱子芹及心叶棱子芹。归叶棱子芹与长果棱子芹是两个近缘种,二者果实较长,在外部形态上极为相似[3,23]。郜鹏等[24]认为归叶棱子芹与长果棱子芹在果实微形态上有区别,前者果实表面光滑而后者果实表面粗糙。本研究显示这两种棱子芹果实表面均为平滑,细胞外壁无明显突起,油管均为沟里1个,结合面2个。支持二者为近缘种。而郜鹏等[24]观察到的长果棱子芹具粗糙的果实表面可能是因为作者用的是干燥的实验材料所致。果皮的细胞特征只有在其充分吸水膨胀后才能展现其自然的状态。其他几种棱子芹的果实表面特征同归叶棱子芹与长果棱子芹十分相似,果实多数较长,分果长宽比可达3.0~3.5∶1(图1B,M),沟里除了青藏棱子芹沟里油管3个,结合面油管6~7个(图3O)外均为沟里1个,结合面2个(图3B,M)。
组3:翅缘波状,外果皮细胞外壁向外突起,与中果皮易分离,中果皮无结晶。这个组包括雅江棱子芹,宝兴棱子芹,鸡冠棱子芹,翼叶棱子芹,丽江棱子芹,松潘棱子芹,垫状棱子芹,单茎棱子芹,青海棱子芹,棱子芹及瘤果棱子芹。所有这些物种果实的翅通常宽于其他两组。沟里油管1个,结合面油管2个,只有单茎棱子芹沟里油管可能为2个而结合面油管为3个。鸡冠棱子芹的翅被描述为鸡冠状翅[23],但作者认为鸡冠棱子芹与青海棱子芹的翅并无明显区别(图1F,R),故均称为波状。此外这组棱子芹中许多物种果皮的表面具瘤状突起(如雅江棱子芹,宝兴棱子芹及青海棱子芹),而并非植物志描述的仅瘤果棱子芹具瘤状突起[23]。松潘棱子芹和瘤果棱子芹,以及滇芎属(Physospermopsis)的木里滇芎(P.muliensis)和丽江滇芎(P.shaniana)被分子系统学放入棱子芹族[12-13]。然而Downie等[14]指出棱子芹族除了包括棱子芹属外还包括Eleutherospermum,Molopospermum及Physospermum。蒲吉霞[11]指出松潘棱子芹,翼叶棱子芹,丽江棱子芹及棱子芹亲缘关系较近, 松潘棱子芹和翼叶棱子芹染色体数为2n=18。我们观察了位于棱子芹族的一些物种(Eleutherospermumcicutarium,Molopospermumpeloponnesiacum,Physospermumcicutarium,P.verticillatum,Physospermopsisalepidioides,P.delavayi)果实的中果皮,他们均无球状结晶(作者的观察,尚未发表)。
迄今为止分子系统学研究的棱子芹仅有少数几种[11-14]。本研究显示棱子芹属果实的微形态,即翅缘特征,外果皮细胞形态,外果皮与中果皮是否易分离,中果皮油管数目及是否具有结晶有重要的分类学价值。根据这些特征可以将21种棱子芹分为3组:组1(喜马拉雅棱子芹,云南棱子芹及美丽棱子芹)和组2(归叶棱子芹等7种)果实的外形较为相似,翅不呈波状,边翅占果宽度的30%~40%,外果皮细胞平滑,与中果皮不易分离。但组1果实通常比组2短,中果皮有球状结晶,而组2中中果皮无球状结晶。组3(松潘棱子芹等11种)果实的翅缘波状,边翅占果宽度的40%~70%,外果皮具瘤突,细胞外壁明显向外突起,外果皮与中果皮易分离,中果皮无球状结晶。果实结构为分子系统学揭示的棱子芹属是复系的提供了形态学依据,同时为分子系统学将喜马拉雅棱子芹及云南棱子芹应与其他棱子芹分开,并将部分棱子芹(如松潘棱子芹及翼叶棱子芹等)移入棱子芹族提供了形态学支持。此外美丽棱子芹具有与喜马拉雅棱子芹及云南棱子芹一样的球状结晶,说明三者可能亲缘关系较近。果实结构的研究完善了棱子芹属形态学,并为进一步深入研究世界棱子芹属,以及棱子芹属与其他类群的关系提供了形态学基础。
References:
[1]Drude O. Umbelliferae. In: Engler A, Prantl K. Die Natürlichen Pflanzenfamilien[M]. Leipzig: Wilhelm Engelmann, 1897-1898.
[2]Pimenov M G, Leonov M V. The Genera of the Umbelliferae: a Nomenclator[M]. London: Royal Botanic Gardens, Kew, UK, 1993.
[3]She M L, Pu F T, Pan Z H,etal. Apiaceae. In: Flora of China Editorial Committee. Flora of China (14)[M]. St. Louis, Missouri: Missouri Botanical Garden Press, 2005.
[4]Clarke C B. Umbelliferae. In: Hooker J D. Flora of British India 2[M]. London: L. Reeve and Co., 1879: 665-720.
[5]Wolff H. Neue Umbelliferen-Gattungen aus Ostasien. Notizblatt des Königlichen Botanischen Gartens und Museums zu Berlin-Dahlem, 1925, 9: 275-280.
[6]Wolff H. Umbelliferae:Plantaesinensesa Dre. H. Smith annis 1921-1922 lectae. XVI. Acta Horti Gothoburgensis, 1926, 2: 288-328.
[7]Mukherjee P K, Constance L. Umbelliferae (Apiaceae) of India[M]. New Delhi: American Institute of Indian Studies and Oxford and IBH Publishing, 1993: 279.
[8]Pan Z H, Watson M F. Apiaceae:Physospermopsis,Pleurospermum,Pleurospermopsis,Sinolimprichtia. Flora of China(14) Raven[M]. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press, 2005.
[9]Pimenov M G, Kljuykov E V. Taxonomic revision ofPleurospermumHoffm. and related genera of Umbelliferae. II: The generaPleurospermum,Pterocyclus,Trachydium,Keraymonia,Pseudotrachydium,Aulacospermum, andHymenolaena. Feddes Repertorium, 2000, 111(7-8): 517-534.
[10]Horn af Rantzien H. OmPleurospermumaustriacum(L.) Hoffm. emend Turcz., dess taxonomi, utbredning och ekologi. Svensk Botanisk Tidskrift, 1946, 40: 179-213.
[11]Pu J X. Systematics of GenusPleurospermumin Sichuan of China[D]. Chengdu: Sichuan University, 2007.
[12]Zhou J, Peng H, Downie S R,etal. A molecular phylogeny of Chinese Apiaceae subfamily Apioideae inferred from nuclear ribosomal DNA internal transcribed spacer sequences. Taxon, 2008, 57(2): 402-416.
[13]Zhou J, Gong X, Downie R D. Towards a more robust molecular phylogeny of Chinese Apiaceae subfamily Apioideae: additional evidence from nrDNA ITS and cpDNA intron (rpl16 and rps16) sequences. Molecular Phylogenetics and Evolution, 2009, 53(1): 56-68.
[14]Downie S R, Plunkett G M, Watson M F,etal. Tribes and clades within Apiaceae subfamily Apioideae: the contribution of molecular data. Edinburgh Journal of Botany, 2001, 58(2): 301-330.
[15]Tseng C C. Anatomical Studies of Flower and Fruit in the Hydrocotyloideae (Umbelliferae)[M]. California: University of California Press, 1967: 1-58.
[16]Henwood M J, Hart J M. Towards an understanding of the phylogenetic relationships of Australian Hydrocotyloideae (Apiaceae). Edinburgh Journal of Botany, 2001, 58(2): 269-289.
[17]Wang Y T, Liu M, Cheng X Y. Trichomes and fruit structures ofAngelicaand related genera of the family Apiaceae and their taxonomic significance. Acta Prataculturae Sinica, 2015, 24(4): 66-75.
[18]Liu M, Van Wyk B E, Tilney P M. The taxonomic value of fruit structure in the subfamily Saniculoideae and related African genera (Apiaceae). Taxon, 2003, 52(2): 261-270.
[19]Liu M, Van Wyk B E, Tilney P M. Irregular vittae and druse crystals inSteganotaeniafruit support a taxonomic affinity with the subfamily Saniculoideae (Apiaceae). South African Journal of Botany, 2007, 73(2): 252-255.
[20]Liu M, Van Wyk B E, Tilney P M,etal. Evidence from fruit structure supports in general the circumscription of Apiaceae subfamily Azorelloideae. Plant Systematics and Evolution, 2009, 280(1-2): 1-13.
[21]Liu M, Van Wyk B E, Tilney P M. Unusual carpological characters inMarlothiellagummifera(Apiaceae). South African Journal of Botany, 2012, 83: 19-22.
[22]Liu M, Plunkett G M, Van Wyk B E,etal. The phylogenetic significance of the carpophore in Apiaceae. Annals of Botany, 2012, 110(8): 1531-1543.
[23]Editorial Committee of Flora of Chinese China Academy of Sciences. Flora Repubulicae Popularis Sinicae (Volume fifty-fifth, The third section)[M]. Beijing: Science Press, 1992.
[24]Gao P, Liao C Y, He X J. Micromorphological features of pericarp surface ofPleurospermum(Apiaceae) in China and its taxonomic significance. Acta Botanica Boreali-Occidentalia Sinica, 2001, 31(9): 1765-1773.
[25]Wang H P, Ma X G, Gao P,etal. Fruit anatomical features of sixteen species and one variety ofPleurospermum(Apiaceae) in China and their taxonomic significance. Acta Botanica Boreali-Occidentalia Sinica, 2014, 34(4): 711-719.
[26]Feder N, O’Brien T P. Plant microtechnique: some principles and new methods. American Journal of Botany, 1968, 55(1): 123-142.
参考文献:
[11]蒲吉霞. 四川棱子芹属PleurospermumHoffm系统学研究[D]. 成都: 四川大学, 2007.
[17]王宇婷, 刘玫, 程薪宇. 中国伞形科当归属及相关类群毛状体和果实结构的系统学价值.草业学报, 2015, 24(4): 66-75.
[23]中国科学院中国植物志编辑委员会. 中国植物志(第55卷,第1分册)[M]. 北京: 科学出版社, 1992.
[24]郜鹏, 廖晨阳, 何兴金. 中国伞形科棱子芹属植物果实表面微形态特征及其分类学意义. 西北植物学报, 2011, 31(9): 1765-1773.
[25]王会朋, 马祥光, 郜鹏, 等. 中国棱子芹属16种1变种的果实特征及其分类学意义. 西北植物学报, 2014, 34(4): 711-719.
Taxonomy ofPleurospermum(Apiaceae) species in China based on micromorphology of fruit
WEI Xue-Ying, LIU Mei*, RU Jian, WANG Yu-Ting, WANG Chao-Qun, GUO Shi-Qi
KeyLaboratoryofPlantBiology,CollegeofHeilongjiangProvince,CollegeofLifeScienceandTechnology,HarbinNormalUniversity,Harbin150025,China
Abstract:The micromorphology of the fruits of 21 species of Pleurospermum (Apiaceae) was evaluated for the first time in this study. The wing, exocarp, mesocarp, vascular bundle, secretory duct, commissure, endosperm, and crystals of the fruits of 21 Pleurospermum species were observed under a light microscope. The structure of Pleurospermum fruits varied greatly and the studied taxa could be divided into three groups. In group one (three taxa including P. hookeri) and group two (seven taxa including P. angelicoides), the wing margins were not wavy, the outer walls of the exocarp were flat, and the exocarp was difficult to separate from the mesocarp. Spherical crystals were present in the mesocarp in group one but not group two. In group three (11 taxa including P. franchetianum), the wing margins were wavy, the outer walls of the exocarp bulged, and the exocarp was easily separated from the mesocarp. There were no spherical crystals in the mesocarp of group three. These analyses of the micromorphology of fruits confirmed that the genus Pleurospermum is polyphyletic, that there is a close relationship between P. hookeri and P. yunnanense, and provided evidence to support the placement of P. decurrens, P. franchetianum, and related taxa into the tribe Pleurospermumeae. The results of this study also provide morphological data for further research on this genus and related taxa.
Key words:Pleurospermum; fruit; wing; Apiaceae; taxonomy
*通信作者
Corresponding author. E-mail: m.r.liu@126.com
作者简介:魏雪莹(1992-),女,黑龙江齐齐哈尔人,在读硕士。E-mail: hsdweixueying@163.com
基金项目:国家自然科学基金(31270235及31070169)资助。
*收稿日期:2015-06-02;改回日期:2015-07-17
DOI:10.11686/cyxb2015284
http://cyxb.lzu.edu.cn
魏雪莹,刘玫,茹剑,王宇婷,王超群,郭诗琪. 中国伞形科棱子芹属果实微形态及其分类学研究. 草业学报, 2016, 25(4): 111-120.
WEI Xue-Ying, LIU Mei, RU Jian, WANG Yu-Ting, WANG Chao-Qun, GUO Shi-Qi. Taxonomy ofPleurospermum(Apiaceae) species in China based on micromorphology of fruit. Acta Prataculturae Sinica, 2016, 25(4): 111-120.
