北京山区栓皮栎林优势种群点格局分析
2016-05-07樊登星余新晓
樊登星,余新晓,*
1北京林业大学水土保持学院,北京 1000832水土保持与荒漠化防治教育部重点实验室,北京 100083
北京山区栓皮栎林优势种群点格局分析
樊登星1,2,余新晓1,2,*
1北京林业大学水土保持学院,北京100083
2水土保持与荒漠化防治教育部重点实验室,北京100083
摘要:采用点格局分析方法对北京山区栓皮栎林主要种群和不同发育阶段种群的空间分布格局及相互关系进行分析。结果表明:(1)群落物种组成简单,栓皮栎和油松是群落的优势种群,优势种群以中径木为主,缺乏可供更新的幼苗,种群呈现出衰退趋势;(2)在研究尺度内,栓皮栎、油松单种的空间分布格局以聚集型分布为主,随着尺度的增大呈现出随机分布和均匀分布的趋势;栓皮栎和油松呈现显著负相关;(3)优势种群不同发育阶段的空间分布格局与研究尺度紧密相关,栓皮栎小树和中树以聚集型分布为主、大树呈随机分布,而油松不同发育阶段均以随机分布为主;(4)栓皮栎、油松种群不同发育阶段之间的空间关联主要发生在小尺度范围内,小树和中树以正关联为主,而中树和大树、小树和大树以负关联为主;随着研究尺度的增大趋于无关联。
关键词:栓皮栎林;径级结构;点格局分析;空间分布格局;空间关联性
樊登星,余新晓.北京山区栓皮栎林优势种群点格局分析.生态学报,2016,36(2): 318-325.
Fan D X,Yu X X.Spatial point pattern analysis of Quercus variabilis and Pinus tabulaeformis populations in a mountainous area of Beijing.Acta Ecologica Sinica,2016,36(2): 318-325.
种群分布格局是指种群个体在水平空间的配置状况或分布状况,反映了种群个体在水平空间上彼此间的相互关系[1]。种群分布格局及其关联性对种群的生长、发育、资源利用等具有显著的影响[2]。分析种群分布格局有助于认识种群的生物学特性(如生活史策略、喜光、耐阴等)、生态学过程(如种子扩散、种内和种间竞争、干扰等)及其与环境因子之间的相互关系[1-3]。按照种群内个体的聚集程度和方式,种群分布格局一般可分为随机分布(Random distribution)、均匀分布(Regular distribution)和集群分布(Clumped distribution)3种分布型。
空间分布格局的研究方法很多,传统的方法可以分为样方法、无样地法、点格局分析法和分形理论等[4]。点格局分析的理论由Ripley在1977年提出[5],它克服了传统方法只能分析单一尺度空间分布格局的缺点,目前已被广泛应用于种群布格局的分析中[6-8],该方法通过划定一定面积的研究区域,以植物种群个体在二维空间上的坐标为基础数据,构成点图,再以点图为基础进行格局分析。它可以分析各种尺度下的种群格局和种间关系,在拟合分析的过程中最大限度地利用了坐标图的信息,因而检验能力较强[3]。近年来国内许多学者应用点格局分析方法,研究了内蒙古乌拉山[6]、华北关帝山[9]、东北长白山[10]、新疆天山[11]、北京东灵山[12]、山西灵空山[13]等地区的植物种群分布格局及其关联性,从不同尺度揭示了植物种群的生态学特性和形成过程。
栓皮栎(Quercus variabilis)为壳斗科,栎属植物,是我国分布最广的乔木树种和最重要的造林树种之一[14]。栓皮栎林是北京山区森林植被的顶级群落[15],其在水源涵养、水土保持等方面发挥着重要的作用,在北京山区森林植被中占有重要地位。本研究以北京山区典型的栓皮栎林为研究对象,采用点格局的分析方法对栓皮栎林内优势种群及其种群不同发育阶段的空间分布格局和关联性进行分析,以期深入认识北京山区主要优势种群的空间格局特征,为区域森林经营和管理提供理论依据。
1研究区概况
鹫峰国家森林公园位于北京市海淀区西北部苏家坨镇境内,北纬39°54',东经116°28',南连太行山,北接燕山山脉,横跨海淀和门头沟两个区,面积832.04hm2。鹫峰主峰海拔465m,公园最高峰1153m。该地区具有暖温带的气候特征,夏季炎热多雨,冬季寒冷干燥,年降水量为650—750mm,夏季降水量占年降水量的74%,年均气温12℃,≥10℃积温在4200℃左右,无霜期190—200d。土壤主要发育在花岗岩、石灰岩、凝灰岩、砂岩等岩石风化形成的残积、坡积物母质上,海拔800m以下分布着淋溶褐土,海拔800m以上为棕壤。该地区主要以天然次生林和人工林森林群落为主,主要的树种有油松(Pinus tabulaeformis)、栓皮栎(Quercus variabilis)、侧柏(Platycladus orientalis)和刺槐(Robinia pseudoacacia)等。
2研究方法
2.1样地调查
2009年6月在北京鹫峰国家森林公园采用样地调查法进行实地调查,设置栓皮栎林样地1个,样地面积为100m×100m,样地分成20m×20m的方形网格,网格从左到右、从上到下依次放置(网格按正南、正北方向放置),再以每个网格为调查单元,对样地内的木本植物(DBH≥1cm)进行每木检尺,记录其种名、高度、冠幅及其在网格中的中心位置并挂永久标牌。
2.2龄级划分
本文以树木的胸径(DBH)大小作为表征树木年龄大小的指标,对优势种群栓皮栎进行龄级划分,按胸径大小将栓皮栎活立木划分为3个龄级,胸径DBH≤7.5cm为小树,7.5cm<胸径DBH≤22.5cm为中树,胸径DBH>22.5cm为大树。采用点格局的分析方法,分析不同发育阶段个体的空间分布格局及其关联性。
2.3点格局分析方法
本文采用Ripley's L函数进行空间点格局的分析,Ripley's L函数是由Ripley's K函数改进而来。Ripley's K函数由Ripley1977年提出[5],是以植物个体坐标(位置)为基础,分析种群空间格局的工具,该分析方法能够分析任意尺度下的空间分布格局,是分析种群空间分布格局最常用的方法。
2.3.1种群分布格局
简单来说,Ripley's K函数就是考量研究区(样方)内以某点为圆心,以一定长度r为半径的圆内的植物个体数目的函数。计算公式如下:

式中,A是研究区(样方)的面积; n为个体数; uij为两个点i和j之间的距离; Ir(uij)为指示函数,当uij≤r时,Ir(uij)= 1,当uij>r时,Ir(uij)= 0; wij为权重值,用于边缘校正。
为了更直观更简单地解释实际的空间格局,通常用Ripley's L函数代替:

当L(r)= 0,为随机分布;当L(r)>0,为聚集分布,表明在以任意点为圆心,半径为r的圆内的个体数比在空间完全随机情况下多;当L(r)<0,为均匀分布,表明以任意点为圆心,半径为r的圆内的个体数比在空间完全随机情况下少。采用Monte-Carlo模拟99%置信区间,进行结果偏离随机状态的显著性检验。L(r)值位于置信区间之上,种群呈聚集分布; L(r)值位于置信区间之下,种群呈均匀分布; L(r)值位于置信区间之内,种群呈随机分布。
2.3.2种内及种间空间关系
种间的关系分析实际上是两个种的点格局分析,也可以认为是某个特定种个体间的关系研究,种内及种间空间关系的公式为:

式中,n1和n2分别为种1和种2的个体数i和j分别代表种1和种2的个体。同样,用L12(r)取代K12(r),其公式为:

当L12(r)= 0,表明两个种在r尺度下无关联性;当L12(r)>0,表明二者为空间正关联;当L12(r)<0,表明二者为空间负相关。采用Monte-Carlo模拟99%置信区间,当L12(r)值位于置信区间之上时,2个变量显著正相关;当L12(r)值位于置信区间之下时,2个变量显著负相关; L12(r)值位于置信区间之内,2个变量相互独立。
本文所有数据分析过程都通过SPPA和Excel软件完成。
3结果与分析
3.1群落物种组成
栓皮栎林样地乔木层内共有11种乔木树种,林分密度为742株/hm2,林分郁闭度0.75。栓皮栎和油松的个体数分别为536株、139株,占总个体数的90.97%。根据该群落重要值可知(表1),栓皮栎和油松的重要值之和为90.99,说明栓皮栎和油松在该群落中占有绝对优势,是该群落的优势种群。主要的伴生树种有臭椿(Ailanthus altissima)、黄栌(Cotinus coggygria Scop)、刺槐和侧柏等。

表1 栓皮栎林树种组成Table 1 The tree species composition of Quercus variabilis
3.2径级结构
栓皮栎和油松的径级结构均呈近似的正态分布(图1)。在0—16cm径级范围内,栓皮栎个体数随着径级的增大逐渐增多,在16—28cm径级范围内,个体数随径级的增大而逐渐减少,个体数最多的径级为12—16cm和8—12cm,占栓皮栎个体总株数的64.55%。在0—12cm径级范围内,油松个体数随着径级的增大逐渐增多,在12—28cm径级范围内,个体数随径级的增大而逐渐减少,个体数最多的径级为8—12cm和12— 16cm,占油松个体总株数的67.62%。由径级分布可知,由于栓皮栎林内种间竞争激烈,林木分化明显,主要以中径木分布为主,大树和幼树个体较少,表明该群落缺乏可供更新的幼苗,种群呈现出衰退趋势。
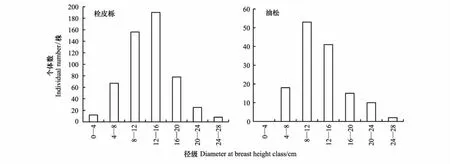
图1 径级分布图Fig.1 The diameter at breast height DBH class distribution
3.3优势种群分布格局
群落和主要种群的个体分布点图如图2所示,从图2可以看出群落和2个主要种群栓皮栎、油松种群都呈明显的集群分布,种群个体分布点图差异较大,种群分布格局与尺度密切相关,但单从图2无法看出种群分布随尺度变化的关系。
主要优势种群栓皮栎和油松的空间分布格局如图3所示。由图3可知:栓皮栎种群在研究尺度0—39m的范围内呈显著性聚集分布,当尺度r=21m时,栓皮栎种群聚集程度最大,此时,L(r)= 2.55;当r>39m时,栓皮栎种群的分布格局呈随机分布,随着尺度的增加,当r>50m时,栓皮栎种群呈现出均匀分布趋势。油松种群在研究尺度0—34m范围内呈现显著性聚集分布,当尺度r = 13m时,油松种群聚集程度最大,此时,L(r)=11.3,当r>34.5m时,油松种群的分布格局呈随机分布,随着尺度的增加,当r>38.5m时,油松种群呈现显著性均匀分布。由此可见,栓皮栎种群和油松种群呈现出了较一致的空间分布趋势,即在研究尺度范围内以聚集性分布为主,但随着研究尺度的增大,先后呈聚集分布、随机分布和均匀分布。由栓皮栎和油松种群的种间关联性分析可知,2个优势种群在整个研究尺度上呈现显著性负相关,且相关性随着尺度的增大而增强,达到显著水平。

图2 群落和主要种群个体分布点图Fig.2 Point diagram of individual distribution of community and main populations in the plot

图3 单种和种间点格局分析Fig.3 Point pattern analysis of Quercus variabilis and Pinus tabulae
3.4栓皮栎不同发育阶段的空间分布格局
栓皮栎种群不同发育阶段的空间分布格局如4所示。由图4可知:栓皮栎小树在0—47.5m内呈显著性聚集分布,当尺度r=28.5m时,其聚集程度最大,此时,L(r)= 6.99,当r>47.5m时,呈随机分布;中树在研究尺度0—36.5m的范围内呈显著性聚集分布,当尺度r=18m时,栓皮栎种群聚集程度最大,此时,L(r)=2.71,当r>36.5m时,分布格局呈随机分布,随着尺度的增加,呈现出均匀分布趋势;大树在整个研究尺度范围内以随机分布为主。
栓皮栎种群不同发育阶段之间的空间关联性随研究尺度的变化而有所不同(图4)。小树和中树在0—41.5m的范围内两者正关联,在41.5—48.5m尺度上无相关性,在大于48.5m尺度上两者负关联,两者的空间关联性先表现为正关联,后随着尺度的增加逐渐变为不相关,最后趋于负关联;小树和大树在8.5—23.5m的尺度范围内呈负关联,在其他尺度范围内无关联;中树和大树在10.5—24m的尺度范围内呈负关联,在其他尺度范围内无关联。小树和大树、中树和大树的空间关联性随尺度的变化而变化,呈现出关联性先由不相关逐渐增大至显著负相关,然后关联性减弱至不关联的变化趋势。

图4 栓皮栎不同发育阶段的点格局分析Fig.4 Point pattern analysis for individuals of Quercus variabilis in different growth stages
3.5油松不同发育阶段的空间分布格局
油松种群不同发育阶段的空间分布格局如图5所示。由图5可知:油松小树在整个研究尺度范围内呈现随机分布;中树在研究尺度2.5—26.5m的范围内呈显著性聚集分布,当尺度r= 7m时,聚集程度最大,此时,L(r)= 2.43,当r>26.5m时,分布格局呈随机分布;大树在研究尺度2—30.5m的范围内呈显著性聚集分布,当尺度r=5m时,聚集程度最大,此时,L(r)=19.1,当r>30.5m时,分布格局呈随机分布。
油松不同发育阶段之间的空间关联性分析表明:小树和中树仅在2—7.5m尺度范围内两者表现出显著正关联,随着尺度的增大趋于无关联;小树和大树在28.5—36m和17—20.5m的尺度范围内呈负相关关系,当r= 35m时,最大负关联强度为14.95,在其他尺度范围内无关联;中树和大树在研究尺度范围内不关联。由此可见,在研究尺度内,油松各发育阶段之间的空间关联性以不关联为主。
4结论与讨论
栓皮栎是北京山区最主要的阔叶造林树种之一,栓皮栎林是北京山区森林植被的顶级群落[15]。在本研究中,群落物种数较少、组成简单,栓皮栎和油松是该群落的优势种群。栓皮栎和油松的径级结构均呈近似的正态分布,主要以中径木分布为主,幼树和大树个体较少,栓皮栎林内缺乏可供更新的幼苗,表明栓皮栎和油松两个优势种群呈现出衰退趋势。林下更新苗匮乏是许多落叶栎林的共同特征[16]。一方面是由于栓皮栎和油松坚果是昆虫、鸟类、啮齿类动物以及微生物的重要食物来源,从坚果成长、成熟、脱落、萌发到幼苗生长等各个阶段一直承受着较大的捕食压力[13];另一方面是由于华北土石山区土层薄,土壤瘠薄,水分条件较差,加之林下郁闭度大,阳光不足,不利于种子萌发生长,从而导致林下更新苗匮乏。栓皮栎和油松都是喜光树种,可以适当地采取择伐等措施创造林窗和林隙,促进其自然更新。
植物种群空间分布格局是植物自身的生物学特性(种子扩散机制、植物繁殖特性、种内种间竞争等)与环境因素(地形、土壤、水分、光照等)共同作用的结果[17-18]。由于栓皮栎和油松的种子体积大籽粒密实,受到重力作用影响种子的下落位置相对集中,主要散落在母树的周围,这种种子传播受限的生物学特性使其在空间上易形成聚集分布的特征。随着栓皮栎和油松幼苗的生长以及个体径级的增大,其对环境资源,如光、水、营养元素和空间等的需求也随之加剧,种内和种间相互作用增强而造成自疏或它疏作用,导致种群的聚集强度逐渐减弱,最终趋于均匀或随机分布,这与蔡飞和范娟等人的研究结果基本一致[19-20]。

图5 油松不同发育阶段的点格局分析Fig.5 Point pattern analysis for individuals of Pinus tabulae in different growth stages
种群的发育过程直接影响其个体的空间分布格局,不同发育阶段的个体往往处于不同林层[21],表现出不同的空间分布格局[22-23]。在本研究中,栓皮栎种群不同发育阶段个体的空间分布格局表现出一定的差异,小树和中树在研究尺度内以聚集分布为主,而大树则呈现出随机分布的趋势。研究表明,随着种群径级(年龄)的增大,受密度制约效应的影响,种群个体死亡率提高,密度下降,种群由聚集分布向随机分布或均匀分布转变[24-26]。种群不同发育阶段个体的分布格局及其相互关系是种群和群落与环境长期相互作用的结果,是种群结构、群落结构合理性的体现,也是种群和群落动态与稳定性的标志[27]。不同发育阶段的个体在空间上的交错分布有利于对各种资源(水、热、光、土壤养分等)的充分利用,并提高其对环境的适应能力,这样有利于整个种群的生存和发展[27]。栓皮栎种群在不同发育阶段的空间分布格局与其生长环境紧密相关,以小树为例,调查发现,样地内的小树多以聚集分布为主,由于小树主要出现在大树个体的斑块间隙,聚集分布有利于小树获取生长所需的光照条件,形成适于自身生长的环境,从而促进小树的成活和生长。
种内联结用来表示同一物种不同龄级在空间分布上的相互关联性,是对一定时期内植物种群组成个体之间相互关系的静态描述,这种关系不仅包括空间分布关系,也包含着个体间的功能关系,揭示了种群的现状,并能反映种群内个体间的相互作用[28-29]。栓皮栎、油松种群的小树和中树在中小尺度上表现出显著的正相关关系,表明两者之间存在正向和相互有利的生态关系[25];而小树和大树、中树和大树在研究尺度上趋于负关联或无关联。这说明同一个群落内在资源限制的条件下,与成熟个体相比较,小树和中树对土壤水分、养分、光照等资源的竞争力微弱,在长期的自然选择过程中形成的生活对策要求小树和中树之间只有相互庇护才有利于发挥群体效应,增大个体存活的机会[7];然而,随着植株个体的不断成长发育,不同发育阶段个体之间对资源需求加剧,它们抵御环境胁迫能力增强,相互之间的庇护作用消弱,从而使得小树和大树以及中树和大树之间的关系呈现出负关联或无关联的趋势[9]。
种群的空间分布格局与空间尺度密切相关,空间分布格局随着尺度的变化而变化,种群空间分布格局的这种尺度变异性受多因素制约,如种内或种间竞争[13]、生境的异质性和斑块性等[21,30]。在本研究中栓皮栎和油松种群在相对较小的尺度上主要呈现聚集分布特征,随着研究尺度的增大,种群的聚集程度减弱并逐渐表现出均匀分布或随机分布的特征,这主要与优势种群的生物学特性以及种内种间竞争有关系。另外,栓皮栎和油松种群不同发育阶段个体之间的正关联或负关联主要发生在小尺度范围内,而且其关联性随着尺度的增大而减弱,并逐渐趋于不关联。以上研究结果体现出了种群空间格局的尺度依赖性[9],说明同一个群落内种群内或不同个体之间的相互作用具有显著的尺度效应,当大于某一个尺度时,个体间的相互作用将明显减弱。
参考文献(References):
[1]DaleM R T.Spatial Pattern Analysis in Plant Ecology.Cambridge: Cambridge University Press,1999: 207-276.
[2]Druckenbrod D L,Shugart H H,Davies I.Spatial pattern and process in forest stands within the Virginia piedmont.Journal of Vegetation Science,2005,16(1): 37-48.
[3]张金屯.数量生态学.北京:科学出版社,2004: 264-268.
[4]张金屯.植物种群空间分布的点格局分析.植物生态学报,1998,22(4): 344-349.
[5]Ripley BD.Modeling spatial patterns.Journal of the Royal statistical Society.Series B,1977,39(2): 172-212.
[6]胡尔查,王晓江,张文军,海龙,张雷,张胜利,徐鹏雁.乌拉山自然保护区白桦种群的年龄结构和点格局分析.生态学报,2013,33(9): 2867-2876.
[7]张震,刘萍,丁易,刘黎明.天山云杉林不同发育阶段种群分布格局研究.北京林业大学学报,2010,32(3): 75-79.
[8]牛丽丽,余新晓,岳永杰.北京松山自然保护区天然油松林不同龄级立木的空间点格局.应用生态学报,2008,19(7): 1414-1418.
[9]闫海冰,韩有志,杨秀清,王丽艳,项小英.华北山地典型天然次生林群落的树种空间分布格局及其关联性.生态学报,2010,30(9): 2311-2321.
[10]王蕾,张春雨,赵秀海.长白山阔叶红松林的空间分布格局.林业科学,2009,45(5): 54-59.
[11]张毓涛,李吉玫,常顺利,李翔,卢建江.天山中部天山云杉种群空间分布格局及其与地形因子的关系.应用生态学报,2011,22(11): 2799-2806.
[12]张育新,马克明,祁建,冯云,张洁瑜.北京东灵山海拔梯度上辽东栎种群结构和空间分布.生态学报,2009,29(6): 2789-2796.
[13]伊力塔,韩海荣,程小琴,康峰峰,张志杰.灵空山林区辽东栎(Quercus liaotungensis)种群空间分布格局.生态学报,2008,28(7): 3254-3261.
[14]吴明作.栓皮栎研究进展.陕西林业科技,1998,(4): 65-69.
[15]吴晓莆,王志恒,崔海亭,方精云.北京山区栎林的群落结构与物种组成.生物多样性,2004,12(1): 155-163.
[16]高贤明,王巍,杜晓军,马克平.北京山区辽东栎林的径级结构、种群起源及生态学意义.植物生态学报,2001,25(6): 673-678.
[17]Condit R,Ashton P S,Baker P,BunyavejchewinS,Gunatilleke S,Gunatilleke N,Hubbell S P,Foster R B,Itoh A,LaFrankie J V,Lee H S,Losos E,ManokaranN,SukumarR,Yamakura T.Spatial patterns in the distribution of tropical tree species.Science,2000,288(5470): 1414-1418.
[18]张春雨,赵秀海,夏富才.长白山次生林树种空间分布及环境解释.林业科学,2008,44(8): 1-8.
[9]Cai F,Song Y C.A study on the structure and dynamics of Schima Superba population on Wuyi Mountain.Acta Phytoecologica Sinica,1997,21(2): 138-148.
[20]范娟,赵秀海,汪金松,张春雨,何俊,夏富才.江西九连山亚热带常绿阔叶林优势种空间分布格局.生态学报,2012,32(9): 2729-2737.
[21]缪宁,刘世荣,史作民,薛泽冰,喻泓,何飞.青藏高原东缘林线杜鹃-岷江冷杉原始林的空间格局.生态学报,2011,31(1): 1-9.
[22]Frost I,Rydin H.Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests.Écoscience,2000,7(1): 38-44.
[23]王绪高,郝占庆,叶吉,张健,李步杭,姚晓琳.长白山阔叶红松林物种多度和空间分布格局的关系.生态学杂志,2008,27(2): 145-150.
[24]Barot S,Gignoux J,Menaut JC.Demography of a savanna palm tree: predictions from comprehensive spatial pattern analyses.Ecology,1999,80(6): 1987-2005.
[25]KenkelNC.Pattern of self-thinning in jack pine: testing the random mortality hypothesis.Ecology,1988,69(4): 1017-1024.
[26]Getzin S,Dean C,He F,Trofymow J A,Wiegand K,Wiegand T.Spatial patterns and competition of tree species in a Douglas‐fir chronosequence on Vancouver Island.Ecography,2006,29(5): 671-682.
[27]张金屯,孟东平.芦芽山华北落叶松林不同龄级立木的点格局分析.生态学报,2004,24(1): 35-40.
[28]王磊,孙启武,郝朝运,田胜尼,张姗姗,陈一锟,张小平.皖南山区南方红豆杉种群不同龄级立木的点格局分析.应用生态学报,2010,21(2): 272-278.
[29]高福元,赵成章,卓马兰草.高寒退化草地不同海拔梯度狼毒种群分布格局及空间关联性.生态学报,2014,34(3): 605-612.
[30]杨华,李艳丽,沈林,亢新刚.长白山云冷杉针阔混交林主要树种空间分布及其关联性.生态学报,2014,34(16): 4698-4706.
Spatial point pattern analysis of Quercus variabilis and Pinus tabulaeformis populations in a mountainous area of Beijing
FAN Dengxing1,2,YU Xinxiao1,2,*
1 School of Soil and Water Conservation,Beijing Forestry University,Beijing 100083,China
2 Key Laboratory of Soil and Water Conservation and Desertification Combating,Ministry of Education,Beijing 100083,China
Abstract:Spatial pattern and spatial association analysis are important methods to study the population characteristics and community structure underlying ecological processes.In this study,spatial point pattern analysis was conducted to examine the spatial distribution pattern of and the spatial associations between the different developmental stages of the Quercus variabilis and Pinus tabulaeformis populations in a mountainous area of Beijing.Only 11 tree populations were observed in the community,in which Q.variabilis and P.tabulaeformis were the dominantspecies.Intermediate-diameter individuals of both species were most prevalent.The populations lacked regenerative saplings,indicating that the populations are in decline.At the study scale,all Q.variabilis and P.tabuliformis individuals were significantly clustered,and as the scale increased,the distribution became more random and regular.The distribution pattern of Q.variabilis populations varied with the spatial scale; they were mainly aggregated at the small scale(0—39 m)and randomly distributed at the large scale(39—50m).The distribution pattern of P.tabuliformis populations was mainly aggregated at the small scale(0—34m)and randomly distributed at the large scale(34—50m).The spatial association between Q.variabilis and P.tabuliformis was significantly negative.The distribution patterns of Q.variabilis and P.tabulaeformis populations at different developmental stages were closely related to the scale.The spatial distribution patterns of young and middle-aged Q.variabilis individuals were significantly clustered,where as those of older trees were random.The spatial distribution pattern of P.tabuliformis wasbook=319,ebook=47significantly random at all scales.Spatial association of Q.variabilis and P.tabulaeformis at different developmental stages was primarily observed at the small scale.The association between middle-aged and young trees was significantly positive at the medium and small scales.However,the spatial association between young,middle-aged,and old trees was negative,and gradually disappeared as the scale increased.These data suggest that the spatial distribution and spatial associations of Q.variabilis and P.tabulaeformis populations were mainly affected by environmental heterogeneity,the biological properties of the populations(especially the seed dispersion mechanism,intraspecific relationships,and interspecific relationships),the differing developmental stages,and changes in spatial scale.
Key Words:Quercus variabilis; DBH class structure; point pattern analysis; spatial distribution; spatial association
*通讯作者
Corresponding author.E-mail: yuxinxiao@ bjfu.edu.cn
收稿日期:2014-02-06;网络出版日期: 2015-06-10
基金项目:国家自然科学基金(41430747,41171028);国家科技支撑项目(2011BAD38B05);国家林业公益性行业科研专项(201104005)
DOI:10.5846/stxb201402060215
