云南高黎贡山赧亢东白眉长臂猿的鸣叫及其行为
2016-05-07吴建普周伟李明会
吴建普,周伟,李明会
(1.西南林业大学林学院,云南省森林灾害预警与控制重点实验室,云南 昆明650224;
2.西藏自治区林业调查规划研究院,西藏 拉萨850000)
云南高黎贡山赧亢东白眉长臂猿的鸣叫及其行为
吴建普1,2,周伟1,李明会1
(1.西南林业大学林学院,云南省森林灾害预警与控制重点实验室,云南昆明650224;
2.西藏自治区林业调查规划研究院,西藏拉萨850000)
摘要:为探讨东白眉长臂猿的鸣叫功能,于2007年春、秋季和2008年春季对分布在高黎贡山赧亢的1♀1♂(成体)1幼的家群和独猿(1♀,成体)跟踪观察,记录其鸣叫频次及行为。研究结果表明,家群年间鸣叫时间和频次差异性显著(p=0.05)。三个季节比较,以2008年春季家群的持续鸣叫时间最长。导致这种情况的原因有三,首先是2007年春季家群需要保护幼猿,所以减少鸣叫;其次,2008年幼猿死亡后家群中雌性个体发情和交配准备导致鸣叫频次和持续时间增加;再次,2008年春季家群和独猿的相对距离较近,导致春季长臂猿鸣叫时间和频次较高。秋季家群成功的二重唱频次高于春季,其二重唱行为更多地支持加强群内雌雄配对关系假说,而非空间和资源防卫假说。群间的合唱还与家群的警戒和防御有关;对雌性独猿而言,长时间对家群的尾随和回应性鸣叫,其功能含义还需深入研究验证。
关键词:东白眉长臂猿;鸣叫行为;鸣叫频次;持续时间;高黎贡山
鸣声是长臂猿类重要的行为之一。长臂猿的鸣声分析主要应用于种间的分类讨论[1];还可以监测长臂猿类种群数量分布变化等。长臂猿鸣声的主要功能为群间的隔离、群体的定位、资源防御(如领域、食物资源或配偶)、群体的形成与维持、配偶吸引、配偶稳固和宣传配偶信息等[2~8]。
白眉长臂猿属(Hoolock)隶属灵长目(Primates)长臂猿科(Hylobatidae),目前具2个种,东白眉长臂猿(Hoolockleuconedys)和西白眉长臂猿(Hylobateshoolock)[9]。白眉长臂猿为典型的一夫一妻制,具领域性和鸣叫习性。国内对滇西东白眉长臂猿的鸣叫研究集中在时间特征分析。研究结果显示,它在不同季节鸣叫发生时间有显著差异,同一季节异地种群间鸣叫持续时间差异显著。当地气候、猿群密度、栖息地状态等对鸣叫有影响[10]。对云南腾冲大塘东白眉长臂猿的鸣叫行为研究表明,鸣叫主要发生在黎明后4h内,平均每次鸣叫持续14min 59s,不同群体间鸣叫持续时间不同。天气情况影响其鸣叫相对黎明的开始时间和持续时间;鸣叫频率在10月和11月明显高于其他月份[11]。对于中国东白眉长臂猿鸣声行为的功能分析还未见报道。
高黎贡山赧亢是东白眉长臂猿的一个分布区。本文旨在比较高黎贡山赧亢3个季节间东白眉长臂猿一个家群(1♀1♂的成体和1幼)和一只独猿(1♀成体)共4只个体的鸣叫及其行为,探讨其鸣叫功能。研究结果可加深对该物种生态生物学的理解,并可为该物种的有效保护提供生态行为基础数据。
1研究地点和研究方法
1.1研究地概况
高黎贡山赧亢地处保山市隆阳区、龙陵县和腾冲县的交界处(25°49′44″N,98°46′7″E),是连接高黎贡山国家级自然保护区的最南端与龙陵小黑山云南省级自然保护区的生境走廊带,面积约13.3km2[12]。高黎贡山南段气候为中国西部型季风气候的暖性湿润型。最热月平均气温14~18℃,最冷月平均气温2~7℃,年平均气温9~13℃,年降水量1 700~2 900mm,属湿润区[13]。本工作集中在赧亢片区东坡的惠坡(最高海拔2 311m)、大马河塘和栽山沟等地。
高黎贡山南段山体西高东低,林分为中山湿性常绿阔叶林,以喜温凉、湿润的壳斗科石栎属(Lithocarpus)、青冈属(Cyclobalanopsis)种类为典型优势的复层混交林。林木高大,林内分布有苔藓、地衣等附生植物。东坡优势树种主要为青冈-石栎林、薄片青冈林、曼青冈林、刺栲-马蹄荷林、虎皮楠-硬斗石栎林等。山脊多为3~10m的中小乔木,而山中部和山谷多为10~30 m的高大乔木。林内乔木、灌木和竹子(云南方竹和带鞘箭竹)混生,密度较高[13]。
1.2研究方法
1.2.1鸣叫定义
长臂猿物种均具有响亮、长而固定模式的叫声,称为鸣唱(songs)。长臂猿个体或群体鸣叫间断不超过10min的一段时间内的鸣叫称一次鸣叫[14]。鸣叫的生物声学定义(Definitions of bioacoustic)参照Tenaza[15],Kappeler[16]:
鸣唱(song)在一段时间内有规律、连续并以一定概率重复着音符(notes)和短语(phrase)的鸣叫。
二重唱(duet song)两个个体以相同的发声法或发声模式共同发出叫声。
激情鸣叫(great call)在一个鸣唱回合中,由雌性长臂猿发出的、音律固定的、最易识别的短语,频率最快。在实地观察中,伴随着激情鸣唱,雌性在树枝间激烈跳动。
合唱(chorus)由多个体鸣叫回合临时交替在一个连续时间内,这个时间通常超越了所有参加个体的鸣叫时间。
尖叫回合(scream bouts)边界冲突时群间的叫声,包括烦恼鸣叫回合。
烦恼鸣叫回合(harassing call bouts)有潜在捕食者存在所作出反应发出的叫声。
1.2.2跟踪方法
跟踪观察时2人为一组。发现目标动物后,采用焦点动物取样法[17],分别跟踪观察家群中的一只目标动物,记录其鸣叫行为。野外跟踪观察自2007年3月9日-4月8日,2007年10月23日-11月29日,2008年4月3日-4月29日,共计96d,观察对象包括1♀1♂(成体)1幼(2008年春季未发现幼体)的家群和1♀(成体)的独猿(2007年秋季未发现独猿)。
1.2.3数据收集
(1)录音及鸣叫时间记录发现东白眉长臂猿鸣叫即以普通MP3录音,持续录音至鸣叫结束,保存声音文件为*.wav格式。录音时尽量靠近鸣叫树,以保证录音质量。使用秒表(型号PC3210)记录鸣叫开始和结束时间及各种类型鸣叫的持续时间。特殊鸣叫类型,如尖叫回合、烦恼鸣叫回合等的持续时间不在鸣叫持续时间和频次中统计。合唱时间不用于二重唱的时间计算。收集录音数据后,在室内重复放音,记录激情鸣叫的开始和结束时间。时间记录都精确到s。
(2)鸣叫行为的记录在跟踪观察中,发现东白眉长臂猿鸣叫并能观察到个体,则记录鸣叫类型及其伴随的行为,收集引发其鸣叫行为变化的事件。收集和记录方法均采用焦点动物取样法和全事件记录法[16],将事件发生的整个过程记录下来。记录鸣叫行为后GPS定位并标记鸣叫树。事件收集包括以下方面。
①鸣叫时个体的主要行为模式(取食、移动、静止、玩耍、逃离、守望等)及时间花费,同时记录与每个行为对应的鸣叫模式。统计各行为模式花费时间占该鸣叫模式花费总时间的百分比。
②引发鸣叫行为的原因,一是发现威胁,如观察者或其他动物;二是发现非家群的其他东白眉长臂猿个体等。
1.3数据处理
频率的计算为鸣叫次数与总观察的天数比。用Kolmogorov-SmirnovZ-检验3个季节每次鸣叫的持续时间数据的正态性。当数据符合正态分布时,采用独立样本t-检验,比较两组样本间的差异;当数据不符合正态分布时,采用非参数Mann-WhitneyU-检验,比较两组样本间的差异。开始鸣叫的集中时间点采用圆形统计法计算[18]。数据分析处理均用Microsoft Excel 2003和SPSS 13.0 for Windows。
2结果与分析
2.1鸣叫时间特征
2.1.1鸣叫时间和频次

表1 东白眉长臂猿鸣叫时间和频率
2.1.2鸣叫的持续时间
Kolmogorov-SmirnovZ-检验结果表明,2007年秋季东白眉长臂猿家群(Z=0.364,P=0.999;n=10),2008年春季家群(Z=0.565,P=0.907;n=11)和独猿(Z=0.687,P=0.733;n=12),2007年春季群体(Z=0.947,P=0.331;n=23)和2008年春季群体(Z=0.823;P=0.508;n=23)的持续鸣叫时间数据均符合正态分布,因此做t-检验。结果表明,唯有2008年春季家群与2007年秋季家群每次鸣叫持续时间数据差异性显著;2008年春季群体(独猿和家群)与2007年春季群体(独猿和家群)每次鸣叫持续时间数据差异性显著(表2)。
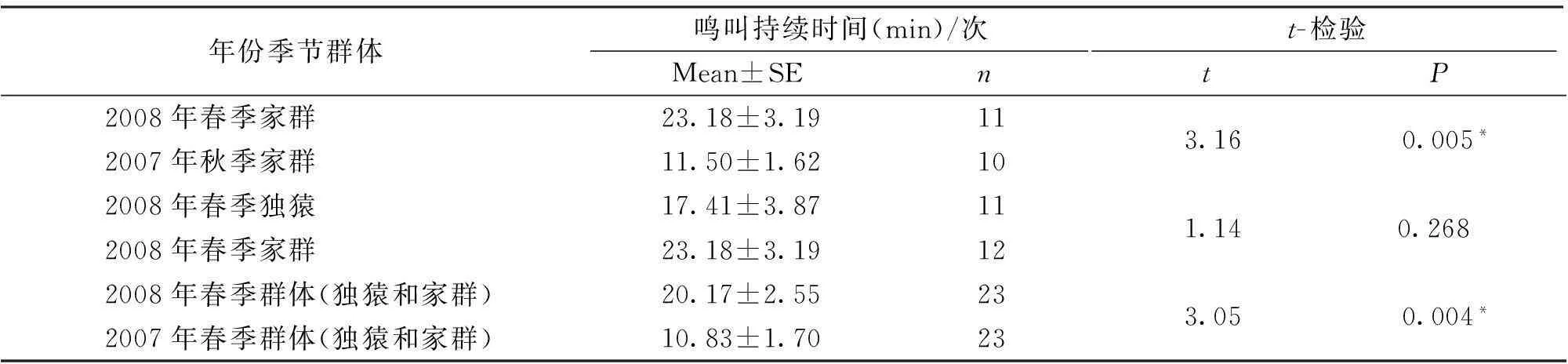
表2 东白眉长臂猿鸣叫持续时间差异性检验
2.2鸣叫类型与伴随行为
东白眉长臂猿每次激情鸣叫时间最短,同时伴随着上下跳跃的行为,偶尔有激烈摇动树枝的行为。有非家群东白眉长臂猿个体在附近活动时,则家群二重唱花费时间较长;无非家群个体在附近活动时,家群二重唱花费的时间较短,伴随行为以静止为主。合唱在所有鸣唱类型中花费时间最长,行为以两群互相守望为主,移动较少。尖叫回合为东白眉长臂猿雄性驱赶独猿时,独猿发出的鸣叫,时间较短,独猿急速逃离后静止,移动较少(表3)。未发现家群中雌性驱赶独猿(1♀,成体)的现象。

表3 不同鸣叫类型的持续时间和伴随行为及原因
3讨论
3.1鸣叫时间和频次的差异及其原因
东白眉长臂猿鸣叫持续时间的年间差异与3个因素有关。首先,与抚育和保护幼体有关。2007年春、秋季家群处于抚育幼体时期,其鸣叫持续时间明显低于没有抚育幼体时期的鸣叫持续时间。一方面,抚育期需要较多的时间照顾幼体,故持续鸣叫时间减少;另一方面,因为鸣叫持续时间较长将增加幼体遭受天敌威胁程度的可能性,影响幼体的安全。黑冠长臂猿(Nomascusconcolor)的鸣叫也有类似现象[18]。其次,与幼猿死亡后家群中雌雄个体发情有关。2007年冬季云南经历了少有的酷寒,到次年春季,原来3个成员的家群仅剩成年雌雄个体,幼猿消失。结果显示,丢失幼猿后成年雌性的激情鸣叫次数高达20次,远比2007年高(表3),且2008年春季家群的鸣叫持续时间亦明显高于2007年春季,差异显著(表2)。而且研究组在2008年4月份观察到该家群雌雄个体的交配行为。综合上述情况,成年雌性个体高频率的激情鸣叫行为应该与发情或者交配行为有关。有研究显示,成功的雌性激情鸣叫次数反映了雌雄之间的配对关系,新配对的群体增加鸣叫频次有利于配偶间的稳定关系[19]。在本项研究中,家群中的雌雄个体属稳定配对关系。再次,群间距离是影响猿群鸣叫频次的另一重要原因。在家群和独猿距离较近的春季,双方鸣叫频次和持续时间均显著高于相互间距离较远的秋季(表2)。该现象反映出鸣叫空间位置展示和个体定位的功能[20~22]。邻近猿群越多,距离越近,鸣声发生的可能性也就越大,次数也就越多[7],结果导致2008年春季的鸣叫频次和持续时间显著高于秋季。
独猿一天两次鸣叫的意义值得关注。独猿的首次鸣叫属于正常的晨鸣,而再次鸣叫发生于与家群相遇后。云南盈江县木笼河的东白眉长臂猿也有类似现象[10]。后一次鸣叫的生物学意义是向家群示好,还是对家群的一种防范,这值得今后关注,认真观察双方的行为。
3.2群间鸣叫行为及功能分析
在研究区,东白眉长臂猿家群的二重唱行为更多地支持加强群内雌雄配对关系假说,而非空间和资源防卫假说。该结论有两个论据:(1)在独猿远离家群的2007年秋季,研究区域内仅该一个家群生活,数据收集期间,该研究区其他灵长类如灰叶猴(Trachypithecusphayrei)、短尾猴(Macacaarctoides)等极少在其家域内出现。这一时期家群成功的二重唱频次最高,其功能应以加强家群内个体间配对关系为主。(2)在家群与独猿频繁相遇的春季,家群成功的二重唱频次并不高(表3),独猿往往在家群二重唱中途插入激情鸣叫,并成功打断家群二重唱的延续。虽然对独猿的这种行为还无法提出合理的解释,但该行为发生时独猿与家群的距离并不远,最近距离不到50m,处于家群领域的核心位置。所以空间和资源防卫假说并不成立。实际上对长臂猿二重唱的空间和资源防卫假说的许多研究结果并不相同。在对灰长臂猿(Hylobatesmuelleri)和白掌长臂猿(H.lar)的野外观察中,研究者发现一个家群的二重唱几乎同时引起相邻家群的二重唱回应。鸣声回放试验证实,在一个家群家域内播放二重唱鸣声会引起该家群的回应,但如果在其家域外播放,则不会引起该家群的回应[23~24]。该结果验证了二重唱的空间和资源防卫假说。但也有学者提出不同意见,克氏长臂猿(H.kossii)鸣叫时多数处在其家域的中心位置而非家域的外围边缘,无法向临近家群传达其家域范围的信息[25],而敏长臂猿(H.agilis)的鸣叫树遍布整个家域范围,与其重要的食物树并无相关性[26]。长臂猿二重唱包括一系列复杂的音节和频率的变化[8],对其功能的确认仍需大量的试验研究。
群间的合唱还与家群的警戒和防御有关。合唱往往在家群与独猿距离较近时发生,且持续时间最长。通常由家群最先鸣叫,独猿随后加入形成合唱。在合唱的过程中,家群和独猿相互守望。在合唱末期,家群开始朝独猿相反的方向移动,而独猿则停留一段时间后继续尾随家群。对印尼瓜哇岛中部银白长臂猿(H.moloch)研究表明,群间合唱花费时间较多,与群间相互警戒和防御有关[1]。对雌性独猿而言,长时间对家群的尾随和回应性鸣叫,其功能含义还需深入研究验证。
参考文献:
[1]Geissmann.T.Duet-splitting and the evolution of gibbon songs [J].Biological Reviews,2002,77:57-76.
[2]Koda H,Nishimura T,Tokuda IT,etal.Soprano singing in gibbons [J].Am J Phys Anthropol,2012;149(3):347-355.
[3]Oyakawa C,Koda H,Sugiura H.Acoustic features contributing to the individuality of wild agile gibbon (Hylobatesagilis)songs [J].Am J Primatol,2007,69(7):777-790.
[4]Lemasson A,Ouattara K,Zuberbühler K.Social learning of vocal structure in a nonhuman primate [J].BMC Evol Biol,2011,11:362.
[5]Koda H,Lemasson A,Oyakawa C,etal.Possible role of mother-daughter vocal interactions on the development of speciesspecific song in gibbons [J].Plos One,2013,8(8):e71432.
[6]Koda H,Oyakawa C,Kato A,etal.Experimental evidence for the volitional control of vocal production in an immature gibbon [J].Behaviour,2007,144(6):681-692.
[7]Savini T,Boesch C,Reichard UH.Varying ecological quality influences the probability of polyandry in white-handed gibbons(Hylobateslar)[J].Biotropica,2009,41:503-513.
[8]Clarke E,Reichard UH,Zuberbühler K.Acoustic measures of gibbon(Hylobateslar)hoo calls in different contexts [J].Evolutionary Biology,2015,15:1-11.
[9]Geissmann T.Status reassessment of the gibbons:results of the Asian Primate Red List workshop 2006 [J].Gibbon Journal,2007,3:5-15.
[10]蓝道英,马世来,李寿昌,等.白眉长臂猿鸣叫的时间特征[J].动物学研究,1999,20(4):273-277.
[11]张岛,袁胜东,崔亮伟,等.云南高黎贡山大塘东部白眉长臂猿鸣叫行为[J].四川动物,2011,30(6):856-860.
[12]白冰,周伟,艾怀森,等.高黎贡山赧亢白眉长臂猿春季栖息地利用[J].动物学研究,2007,28(2):179-185.
[13]薛纪如.高黎贡山国家自然保护区[M].北京:中国林业出版社,1995:14,30,61-90.
[14]Haimoff EH.Acoustic and organizational features of gibbon songs [M].In:Preuschoft H,Chivers DJ,Brockelman WY,Creel N(eds).TheLesserApes:Evolutionaryandbehaviouralbiology.Edinburgh:Edinburgh University Press,1984:333-353.
[15]Tenaza RR.Songs,choruses and counter singing of kloss gibbons(Hylobatesklossii)in Siberut Island [J].Zeit Tierpsychol,1976,40:37-52.
[16]Kappeler M.Vocal bouts and territorial maintenance in the Moloch gibbon [M].In:Preuschoft H,Chivers DJ,Brockelman WY,Creel N(eds).The Lesser Apes:Evolutionary and behavioural biology.Edinburgh:Edinburgh University Press:1984:376-389.
[17]Altmann J.Observational study of behavior:Sampling methods [J].Behaviour,1974,49:227-267.
[18]Fowler J,Cohen L,Jarvis P.Practical Statistics for Field Biology(2nded.)[M].Chichester:John Wiley & Sons,1998.
[19]Fan PF,Xiao W,Huo S,etal.Singing Behavior and Singing Functions of Black-Crested Gibbons(Nomascusconcolorjingdongensis)at Mt.Wuliang,Central Yunnan,China [J].American Journal of Primatology,2009,71:539-547.
[20]Barelli C,Mundry R,Heistermann M,etal.Cues to Androgens and Quality in Male Gibbon Songs [J].Plos One,2013,8(12):1-9.
[21]Fan PF,Liu CM,Luo WS,etal.Can a Group Elicit Duets from Its Neighbours? A Field Study on the Black-Crested Gibbon(Nomascusconcolorjingdongensis)in Central Yunnan,China [J].Folia Primatol,2007,78:186-195.
[22]Helen M,Dooley,Debra S,etal.Singing by male and female Kloss gibbons(Hylobatesklossii)in the Peleonan Forest,Siberut Island,Indonesia [J].Primates,2013,54:39-48.
[23]Mitani JC.Gibbon song duets and intergroup spacing [J].Behaviour,1985,92:59-95.
[24]Raemaekers JJ,Raemaekers PM.Field playback of loud calls to gibbon(Hylobateslar):territorial,sex-specific and species-specific responses [J].Animal Behaviour,1985,33:481-493.
[25]Whitten A.The ecology of singing in Kloss gibbons(Hylobatesklossii)on Siberut Island,Indonesia [J].International Journal of Primatology,1982,3:33-51.
[26]Gittins SP.The Behavior and Ecology of the Agile Gibbon(Hylobatesagilis)[D].PhD dissertation,University of Cambridge,Cambridge,1979.
Singing Frequency and Behavior Significance of Eastern White-browed Gibbon (Hoolock leuconedys)at Nankang,Mt.Gaoligong of Yunnan
WU Jian-pu1,2,ZHOU Wei1,LI Ming-hui1
(1.Key Laboratory of Forest Disaster Warning and Control of Yunnan Province,Southwest Forestry University,Kunming Yunnan 650224,P.R.China;2.Research Institute of Forest Inventory and Planning of Tibet Autonomous Region,Lasa Tibet 850000,P.R.China)
Abstract:In order to investigate the calls function of Eastern White-browed Gibbon(Hoolock leuconedys),a family with the pair bond and their baby,and a solitary female were tracking observed and their singing frequency and behavior were recorded in the spring and autumn of 2007,spring 2008 at Nankang of Mt.Gaoligong,Yunnan.The results showed that there was significant difference(p=0.05)between different year in terms of singing time period and frequency.The call duration of the family was longest in the spring of 2008 because of the following three reasons.Firstly,the less frequency of singing in 2007 was needed due to the family has to protect their baby.Secondly,the female of family was in oestrus and was preparing coition in 2008 and increased their singing frequency and time of duration after lost their cub in 2007 winter.Thirdly,the closer distance between the family and the solitary female in 2008 spring caused increasing of the call frequency.In this study the frequency of successful duet bout of family in spring was higher than that in winter.The function of duet more supported the hypothesis that it could strengthen the relationship of the pair bond rather than the hypothesis that it played the defense function of territory and resource.The chorus between the family and solitary female showed the difference functions.For family,the chorus was associated with the warning and defense function.For solitary female,the function of chorus need to be further studied.
Key words:Hoolock leuconedys;singing behavior;singing frequency;time of duration;Mt.Gaoligong
中图分类号:Q 958
文献标识码:A
文章编号:1672-8246(2016)02-0129-06
通讯作者简介:周伟(1957-),男,博士生导师,主要从事野生动物与自然保护区管理研究。E-mail:weizhouyn@163.com
第一作者简介:吴建普(1982-),男,硕士,主要从事野生动物保护与自然保护区规划研究。E-mail:jianpuwu@163.com
基金项目:西南林业大学重点基金项目(200608Z)和云南省林业厅项目(2009001)资助, 云南省重点学科野生动植物保护与利用建设项目 (XKZ200904) 支持。
*收稿日期:2015-08-02

