毛竹快速生长期光合固碳特征及其与影响因素的关系
2016-04-28李洪吉蔡先锋袁佳丽曾莹莹于晓鹏温国胜浙江农林大学林业与生物技术学院浙江临安3300浙江农林大学亚热带森林培育国家重点实验室培育基地浙江临安3300
李洪吉,蔡先锋,袁佳丽,曾莹莹,于晓鹏,温国胜,2(.浙江农林大学林业与生物技术学院,浙江临安3300;2.浙江农林大学亚热带森林培育国家重点实验室培育基地,浙江临安3300)
毛竹快速生长期光合固碳特征及其与影响因素的关系
李洪吉1,蔡先锋1,袁佳丽1,曾莹莹1,于晓鹏1,温国胜1,2
(1.浙江农林大学林业与生物技术学院,浙江临安311300;2.浙江农林大学亚热带森林培育国家重点实验室培育基地,浙江临安311300)
摘要:为了研究毛竹Phyllostachys edulis快速生长期光合固碳特征及其与主要生态因子的关系,利用Li-6400光合仪测定不同年龄(Ⅰ度竹、Ⅱ度竹、Ⅲ度竹)的毛竹在其快速生长不同时期(前期、中期、后期)的光响应曲线及生态因子(光照强度、气温、大气相对湿度、大气二氧化碳摩尔分数、胞间二氧化碳摩尔分数、气孔导度)。结果表明:①毛竹在快速生长的不同时期、不同竹龄叶片的光合固碳能力的变化特征归纳为:快速生长前期是老竹高于新竹(Ⅱ度竹最高,Ⅲ度竹次之,Ⅰ度竹最低);而快速生长的中期和后期则为新竹高于老竹(Ⅰ度竹最高,Ⅱ度竹次之,Ⅲ度竹最低)。②在快速生长的不同时期,不同竹龄叶片的光合固碳能力的动态变化规律差异显著(P<0.05),新竹(Ⅰ度竹)的光合固碳能力在其快速生长期逐渐升高,而老竹(Ⅱ度竹和Ⅲ度竹)的光合固碳能力则都是中期最低,前期和后期较高,同时Ⅱ度竹均高于Ⅲ度竹。③毛竹在快速生长期,对毛竹叶片光合固碳能力的影响因子由大到小依次为:胞间二氧化碳摩尔分数>气孔导度>光照强度>大气二氧化碳摩尔分数>气温>大气相对湿度。经相关性分析得出,光照强度、气温、大气相对湿度、气孔导度与净光合速率(Pn)呈正相关,大气二氧化碳摩尔分数和胞间二氧化碳摩尔分数与净光合速率呈负相关。图3表1参18
关键词:植物生理学;毛竹;光响应曲线;影响因素;竹龄
毛竹Phyllostachys edulis广泛分布于中国南方中亚热带,现今中国毛竹林面积达270万hm2,占竹林总面积的64.1%,是中国竹类植物中分布最广、面积最大的竹种[1,2]。毛竹的光合产物——竹材、竹笋、竹叶等给人们带来了巨大的经济效益。光合速率决定着光合产物的积累,在一定程度上也决定了产量的高低。另一个方面,光合速率也反映了毛竹固碳的能力。因此,研究毛竹光合特性不仅可以揭示其丰产机制,研发丰产技术,而且也可以在林业应对全球气候变化中,评价毛竹对改善环境的重要作用和贡献能力[3]。近年来,对毛竹固碳的前期研究中,在毛竹林碳密度和林分结构[4-5]、不同管理模式对毛竹林碳储量的影响[6]、毛竹林生态系统碳储量及其分配特征[7]、毛竹林的气体交换特征[3]、毛竹林叶片固碳能力[8]、毛竹林叶片光能利用效率[9]等方面取得了一些研究成果。笔者以浙江省临安市青山镇毛竹林为研究对象,测定了毛竹在其快速生长不同时期(前期、中期、后期)的光响应曲线及影响因子(光照强度、气温、大气相对湿度、大气二氧化碳摩尔分数、胞间二氧化碳摩尔分数、气孔导度),试图从光合固碳的角度揭示毛竹快速生长的固碳机制,以期为毛竹林固碳增汇技术研究提供科学依据。
1 试验地概况、试验材料和实验方法
试验地位于浙江省临安市青山镇研口村,地处30°14′N,119°42′E,属于亚热带地区,气候类型为季风气候,年平均日照时数为1 920.0 h,年平均降水量为1 442.0 mm。
在样地向阳面较平缓的坡面搭建实验观测塔,分别选取长势良好,竹形相差不大的Ⅰ度(1年生)、Ⅱ度(3年生)、Ⅲ度(5年生)的毛竹各2株作为样株,在毛竹的中上层位置(约8 m)的向阳面,随机选择形状完整、健康的叶片,重复测定2次·株-1。用Li-6400便携式光合仪(Li-6400 Portable Photosynthesis System),选择天气晴朗的2014年3月6日(前期)、4月20日(中期)、5月28日(后期)的上午对Ⅰ度、Ⅱ度和Ⅲ度(1年生、3年生、5年生)毛竹的光响应曲线进行测定。每次测定前先对叶片进行1 500 μmol·m-2·s-1光强的光诱导,光强梯度选择为0,10,30,50,80,100,300,500,800,1 000,1 200,1 500 μmol·m-2·s-1,从高向低测定净光合速率(Pn),气孔导度(Cond),胞间二氧化碳摩尔分数(Ci),气温(Tair),大气二氧化碳摩尔分数(Ca)和大气相对湿度(RH)等。
利用Excel 2003和光合助手软件绘制及拟合光响应曲线[10-11],再通过SPSS 19.0软件对试验结果和各影响因素进行相关性分析。
2 结果与分析
2.1毛竹快速生长的不同时期不同竹龄的光响应曲线比较
毛竹快速生长的不同时期不同竹龄的光响应曲线见图1。不同年龄的毛竹在快速生长前期,Ⅱ度竹的净光合速率最高,即光合作用能力最强(6.7 μmol·m-2·s-1),Ⅲ度竹次之,Ⅰ度竹最弱;在快速生长中期,Ⅰ度竹光合作用能力上升到最强(5.2 μmol·m-2·s-1),Ⅱ度竹第2位,Ⅲ度竹最弱;到了快速生长的后期,基本和中期相同,Ⅰ度竹最强(6.6 μmol·m-2·s-1),随后是Ⅱ度竹和Ⅲ度竹。
在快速生长前期的光合作用能力,Ⅱ度竹和Ⅲ度竹高于Ⅰ度竹,应该是年龄大的毛竹在积累养分,而中后期的Ⅰ度竹均处于最高,说明新竹在快速生长开始以后光合固碳能力大大提高;而无论快速生长的哪个时期Ⅱ度竹的光合作用能力都要高于Ⅲ度竹,说明在老竹中年龄越大其光合作用能力有所下降。
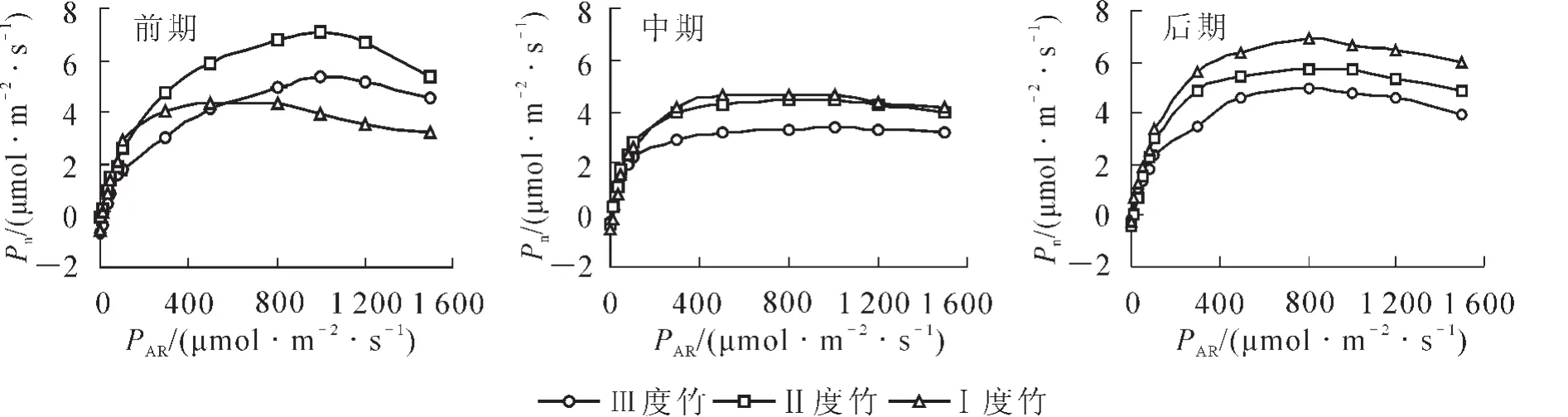
图1 不同时期不同年龄毛竹的光响应曲线比较Figure 1 Comparion of light response curve value for Phyllostachys edulis in different periods and at different ages
2.2毛竹快速生长不同时期不同年龄光合固碳能力的比较
通过SPSS软件,先利用非直角双曲面模型对光响应曲线进行拟合,得到毛竹快速生长的不同时期不同年龄的最大净光合速率(Pmax),然后对不同时期间不同年龄间毛竹作方差分析,分析其差异显著性,再通过Excel绘制固碳能力的柱状图(图2)。
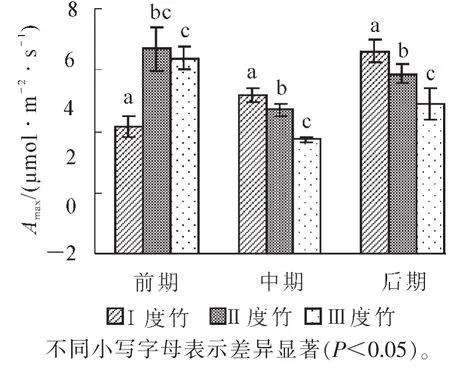
图2 毛竹快速生长不同时期不同竹龄的固碳能力的比较Figure 2 Comparion of photosynthetic carbon fixation capacity for Ph.edulis fast growth period in different periods and at different ages
由图2可以得出:Ⅱ度竹和Ⅲ度竹的净光合速率都呈前后期高,中期低的趋势,不同时期间变化规律显著(P<0.05),中期较前期分别下降了29.8%和41.7%,而后期分别上升了24.8%和31.1%;Ⅰ度竹的净光合速率在前期最弱,中期次之,后期最强,均为升高过程,分别升高24.3%和27.7%。这可以认为老竹在快速生长前期,通过提高其固碳能力储备了较多能量和物质,为新竹的快速生长提供了物质保障,所以Ⅰ度竹在快速生长中期和后期的光合固碳能力会逐渐增强。
2.3毛竹快速生长期光合能力与各影响因素的关系
由图3A可以看出:毛竹快速生长期净光合速率(Pn)在弱光下随着光照强度(PAR)升高而升高,光强在800~1 000 μmol·m-2·s-1时,随着光照强度(PAR)的升高净光合速率(Pn)开始下降;从图3B中可以看出,气孔导度(Cond)越大时,净光合速率(Pn)随之增大,气孔导度(Cond)与净光合速率(Pn)成正相关;图3C和图3D表明:随着胞间二氧化碳摩尔分数(Ci)和大气二氧化碳摩尔分数(Ca)的上升,毛竹快速生长期的净光合速率(Pn)逐渐减小,胞间二氧化碳摩尔分数(Ci)和大气二氧化碳摩尔分数(Ca)与净光合速率(Pn)呈现负相关,且随着净光合速率(Pn)的增加,胞间二氧化碳摩尔分数(Ci)比大气二氧化碳摩尔分数(Ca)的降幅更大;图3E表明:净光合速率(Pn)随着大气相对湿度(RH)的变化规律呈“中间高,两边低”的趋势,净光合速率(Pn)先随着相对湿度(RH)的升高而升高,之后下降,说明相对湿度(RH)的过高过低都会引起净光合速率(Pn)的下降;图3F中,在毛竹快速时期的气温(Tair)变化范围内,气温越高,净光合速率(Pn)值越高,气温(Tair)与净光合速率(Pn)是正相关关系。
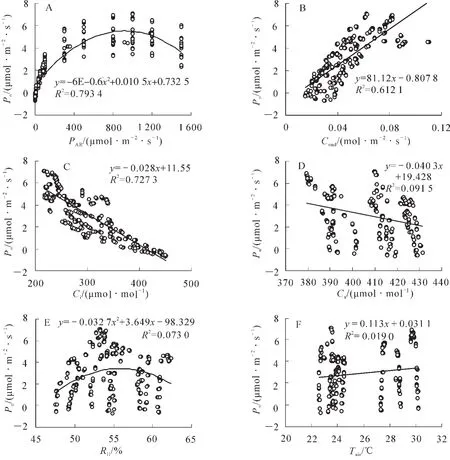
图3 毛竹快速生长期净光合速率与生态因子的关系Figure 3 Relationship between net photosynthetic rate and ecological factors in Phyllostachys edulis fast growth period
2.4光合固碳能力与各影响因子的相关性分析
由表1可见:在毛竹快速生长期内,净光速速率(Pn)与光照强度(PAR),气孔导度(Cond) ,气温(Tair)具有极显著正相关性,且与大气相对湿度(RH)呈显著正相关,而与胞间二氧化碳摩尔分数(Ci)和大气二氧化碳摩尔分数(Ca)则呈极显著负相关。影响程度大小:胞间二氧化碳摩尔分数(Ci)>气孔导度(Cond)>光照强度(PAR)>大气二氧化碳摩尔分数(Ca)>气温(Tair)>相对湿度(RH)。

表1 净光速速率(Pn)与各影响因素的相关性分析Table 1 Correlation analysis of net photosynthetic rate and influencing factors
3 讨论与结论
毛竹具有较强的光合固碳能力[12],而在快速生长时期的有机物质快速积累是其特有的生理特征。通过研究其快速生长不同时期的光响应曲线,比较不同竹龄叶片的光合固碳能力,分析净光合速率与生态因素的关系,有利于深刻认识毛竹林快速生长的固碳机制、固碳增汇技术的研发以及对毛竹林碳通量测定、碳汇估算、碳贸易等都具有十分重要的意义。
研究结果表明:①在幼竹快速生长前期,幼竹(Ⅰ度竹)还处于竹笋状态时,光合作用能力较弱,而老竹(Ⅱ度竹和Ⅲ度竹)光合作用能力增强,其目的是为幼竹的生长积累能量物质;当进入快速生长的中期,随着幼竹的成长,对养分输送的需求量降低,老竹不再需要提供大量有机物,所以其光合作用能力减弱,从而表现出了快速生长前期老竹光合作用能力高于新竹,而在快速生长的中期和后期新竹高于老竹的现象。结合叶淑贤等[13]、施建敏等[14]和WEN等[8]对毛竹光合作用季节变化规律的研究,证明了在快速生长过程中,老竹会为新竹提供一部分所需的营养物质,以提高新竹的生长发育速度。②不同竹龄叶片的光合固碳能力在快速生长的不同时期的动态变化规律也很明显,新竹的光合固碳能力在其快速生长期,随着叶片的发育逐渐升高,而老竹的光合固碳能力则都是中期最低,前期和后期较高,同时Ⅱ度竹高于Ⅲ度竹。这也说明新老竹此消彼长,在新竹快速生长旺期(中期),降低对老竹的资源分配,以优先对新竹的供给[8,14],为在合理安排毛竹林年龄结构,竹林管理等方面提供理论依据。③毛竹在快速生长期,各生理生态因子对毛竹叶片光合固碳能力的显著的影响,通过相关性分析可以得出影响最大的因素为胞间二氧化碳摩尔分数。随着胞间二氧化碳摩尔分数的升高,净光合速率下降,呈极显著正相关;而相关性最小的是相对湿度,表现为显著正相关,毛竹的净光合能力随着相对湿度上升而升高。利用这种关系,一方面能研发生态调控技术,提高毛竹林的固碳能力[15-17];另一方面,可以科学评价毛竹林在应对全球气候变化中的作用和贡献,促进碳交易的健康发展。
同时,在研究过程中发现毛竹林的二氧化碳通量动态、土壤二氧化碳呼吸排放动态、地上地下营养物质的储存、积累动态等因素及其与光合固碳的相互关系都对毛竹的固碳机制都有一定的影响。今后可以进一步研究它们的关系,再从生理整合这一角度来进行深入分析[18]。
4 参考文献
[1]郭起荣,杨光耀,杜天真,等.毛竹学名百年之争[J].世界竹藤通讯,2006,4(1):18-20.GUO Qirong,YANG Guangyao,DU Tianzhen,et al.A debate about moso bamboos latin formal name duration a century[J].Word Bamboo Ratian,2006,4(1):18-20.
[2]邱尔发,陈存及,梁一池,等.不同种源毛竹叶表叶绿素浓度动态[J].福建林学院学报,2002,22(4):312-315.QIU Erfa,CHEN Cunji,LIANG Yichi,et al.Dynamic on chlorophyll concentration of leaves surface of different provenances of Phyllostachys heterocycla cv.pubescens[J].J Fujian Coll For,2002,22(4):312-315.
[3]林琼影.天目山毛竹碳同化特征研究[D].临安:浙江农林大学,2008.LIN Qiongying.Studies on Carbon Dioxide Assimilation Characteristics of Phyllostachys edulis in Tianmu Mountain [D].Lin’an:Zhejiang A &F University,2008.
[4]刘金山,张万耕,杨传金,等.森林碳库及碳汇监测概述[J].中南林业调查规划,2012,31(1):61-65.LIU Jinshan,ZHANG Wangen,YANG Chuanjin,et al.An overview on monitoring of forest carbon pool and carbon sink[J].Cent South For Invent Plan,2012,31(1):61-65.
[5]白尚斌,周国模,王懿祥,等.天目山国家级自然保护区毛竹扩散过程的林分结构变化研究[J].西部林业科学,2012,41(1):77-82.BAI Shangbin,ZHOU Guomo,WANG Yixiang,et al.Stand structure change of Phyllostachys pubescens forest expansion in Tianmushan National Nature Reserve[J].J West China For Sci,2012,41(1):77-82.
[6]周国模,吴家森,姜培坤.不同管理模式对毛竹林碳贮量的影响[J].北京林业大学学报,2006,28(6):51-55.ZHOU Guomo,WU Jiasen,JIANG Peikun.Effects of different management models on carbon storage in Phyllostachys pubescens forests[J].J Beijing For Univ,2006,28(6):51-55.
[7]周国模.毛竹林生态系统中碳储量、固定及其分配与分布的研究[D].杭州:浙江大学,2006.ZHOU Guomo.Carbon Storge,Fiation and Distribution in Mao Bamboo Stands Ecosystem[D].Hangzhou:Zhejing U-niversity,2006.
[8]WEN Guosheng,ZHANG Liyang,ZHANG Ruming,et al.Temporal and spatial dynamics of carbon fixation by moso bamboo(Phyllostachys pubescens)in subtropical China[J].Bot Rev,2011,77(3):271-277.
[9]陈嘉琦,温国胜,王艳红,等.春季毛竹光化学效率空间异质性比较[J].福建林学院学报,2014,34(1):52 -56.CHEN Jiaqi,WEN Guosheng,WANG Yanhong,et al.Comparisom of spatial heterogeneity in photochemical efficiency of Phyllostachys pubescens in spring[J].J Fujian Coll For,2014,34(1):52-56.
[10]刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].中国农学通报,2005,21(8):76-79.LIU Yufeng,XIAO Langtao,TONG Jianhua,et al.Primary application on the non-rectangular hyperbola model for photosynthetic light-respones curve[J].Chin Agric Sci Bull,2005,21(8):76-79.
[11]张利阳,温国胜,王圣杰,等.毛竹光响应模型适用性分析[J].浙江农林大学学报,2011,28(2):187-193.ZHANG Liyang,WEN Guosheng,WANG Shengjie,et al.Four light-response models to estimate photosynthesis of Phyllostachys pubescens[J].J Zhejiang A&F Univ,2011,28(2):187-193.
[12]吴志庄,杜旭华,熊德礼,等.不同类型竹种光合特性的比较研究[J].生态环境学报,2013,22(9):1523-1527.WU Zhizhuang,DU Xuhua,XIONG Deli,et al.A comparative study on photosynthetic characteristics of different types of bamboos[J].Ecol Environ Sci,2013,22(9):1523-1527.
[13]叶淑贤,陆媛媛,朱文强,等.断鞭对毛竹竹笋—幼竹高生长的影响[J].西北林学院学报,2013,28(1):100-103.YE Shuxian,LU Yuanyuan,ZHU Wenqiang,et al.Effects of cutting rhizome on the height growth of shoots and seedlings of Phyllostachys edulis[J].J Northwest For Univ,2013,28(1):100-103.
[14]施建敏,郭起荣,杨光耀.毛竹光合动态研究[J].林业科学研究,2005,18(5):551-555.SHI Jianmin,GUO Qirong,YANG Guangyao.Study on the photosynthetic dynamic variation of Phyllostachys edulis [J].For Res,2005,18(5):551-555.
[15]施建敏,郭起荣,杨光耀.CO2浓度倍增下毛竹光合作用对光照强度的季节响应[J].江西农业大学学报,2007,29(2):215-219.SHI Jianmin,GUO Qirong,YANG Guangyao.Seasonal photosynthetic responses of Phyllostachys edulis to light under doubled CO2concentration[J].Acta Agric Univ Jiangxi,2007,29(2):215-219.
[16]陈建华,毛丹,马宗艳,等.毛竹叶片的生理特性[J].中南林学院学报,2006,26(6):76-80.CHEN Jianhua,MAO Dan,MA Zongyan,et al.Physiological characteristics of leaves of bamboo Phyllostachys pubescens[J].J Cent South For Univ,2006,26(6):76-80.
[17]许大全,李德耀,邱国雄,等.毛竹叶光合作用的气孔限制研究[J].植物生理学报,1987,13(2):154-160.XU Daquan,LI Deyao,QIU Guoxiong,et al.Studies on stomatal limitaion of photosynthesis in the bamboo leaves [J].Plant Physiol J,1987,13(2):154-160.
[18]庄明浩,李迎春,陈双林.竹子生理整合作用的生态学意义及研究进展[J].竹子研究汇刊,2011,30(2):5 -9.ZHUANG Minghao,LI Yingchun,CHEN Shuanglin.Advances in the researches of bamboo physiological integration and its ecological significance[J].J Bamboo Res,2011,30(2):5-9.
Photosynthetic carbon fixation in Phyllostachys edulis during its fast growth period
LI Hongji1,CAI Xianfeng1,YUAN Jiali1,ZENG Yingying1,YU Xiaopeng1,WEN Guosheng1,2
(1.School of Forestry and Biotechnology,Zhejiang A &F University,Lin’an 311300,Zhejiang,China;2.The Nurturing Station for the State Key Laboratory of Subtropical Silviculture,Zhejiang A &F University,Lin’an 311300,Zhejiang,China)
Abstract:To explain the relationship between the photosynthesis and the fast growth of Phyllostachys edulis,the photosynthetic carbon fixation characteristics of Ph.edulis during its fast growth period and relationships between the main ecological factors and net photosynthetic rate(Pn)were studied.Samples of different ages:Ⅰ(1-year old),Ⅱ(3-year old),andⅢ(5-year old);Ph.edulis were selected to determine the light response curve for different periods(prophase,metaphase,and anaphase)and to show the effect on Pnof factors such as light intensity(P(AR)),air temperature(T(air)),relative humidity(HR),stomatal conductance(C(ond)),atmospheric CO2concentration(Ca),and intercellular CO2concentration(Ci)using an Li-6400 Portable Photosynthesis System.Results showed that:(1)in the prophase photosynthetic carbon fixation capacity of the older ages(ⅡandⅢ)was stronger than the younger age(Ⅰ)withⅡbeing the strongest.However,in the metaphase and anaphase,the 1-year old was strongest(P<0.05).(2)Photosynthetic carbon fixation capacity for leaves inbook=12,ebook=15different developmental stages differed with younger plants(Ⅰ)rising gradually and older plants(ⅡandⅢ)being minimal in the metaphase and stronger in the prophase and anaphase.(3)During the fast growth period,leaf photosynthetic carbon fixation capacity influenced variables in the order:Ci>C(ond)>P(AR)>Ca>T(air)>HR.A positive relationship was found for Pnversus P(AR),T(air),HR,or C(ond),and a negative relationship was found for Pnversus Caor Ci.This study indicated that the photosynthetic carbon fixation capability was very strong in the fast growth period of Ph.edulis with older plants being able to store and provide energy for new shoots.[Ch,3 fig.1 tab.18 ref.]
Key words:plant physiology;Phyllostachys edulis;light response curve;influencing factors;age
作者简介:李洪吉,从事森林生态学研究。E-mail:458857732@qq.com。通信作者:温国胜,教授,博士,从事生态学研究。E-mail:wgs@zafu.edu.cn
基金项目:国家自然科学基金资助项目(31270497);浙江省与中国林业科学研究院省院合作林业科技项目(2014SY16)
收稿日期:2014-12-25;修回日期:2015-04-02
doi:10.11833/j.issn.2095-0756.2016.01.002
中图分类号:S718.4
文献标志码:A
文章编号:2095-0756(2016)01-0011-06
