2个环境条件下花生产量性状的遗传分析
2016-04-23江建华王晓帅倪皖莉
江建华,王 嵩 ,王晓帅,倪皖莉 ,汪 清
(1.安徽省农业科学院作物研究所,安徽合肥 230031;2.山东省农村科技教育培训中心,山东济南 250013)
中国是世界上最大的花生生产消费国。花生种植面积居国内农作物的第7位,而单位面积产值却居于第5位[1]。结果枝数、单株结果数和百果质量是花生重要的产量性状。笔者前期曾对126份花生材料的12个农艺性状与单株生产力之间的关系进行了相关性分析,发现单株果数和单株饱果数对花生单株生产力的影响最大,其次为结果枝数和百果质量[2]。前人利用不同的研究材料和分析方法,均认为结果枝数、单株结果数和百果质量与花生产量呈(极)显著正相关,即进行花生高产育种时,应以这3个性状为主攻目标[3-4]。
前人曾利用主基因+多基因混合遗传模型对花生的多个性状进行了遗传分析,但主要集中于花生的品质性状[5-6]和休眠性[7]等方面。而对于农艺性状的主基因+多基因遗传鲜见报道。刘华等[8]利用主基因+多基因遗传模型对郑9001/郑8903 RIL群体的主茎高和侧枝长性状进行了遗传分析,认为这2个性状主要受多基因效应影响。张新友等[9]利用郑8903/豫花4号RIL群体对结果枝数、百果质量等10个农艺及产量性状进行了遗传分析,认为结果枝数受3对独立的等加性主基因控制,无多基因效应;百果重受2对主基因加多基因控制,以主基因遗传为主。笔者[10]前期曾以远杂9102、皖花4号以及远杂9102/皖花4号F1和F2群体为研究对象,利用南京农业大学章元明教授提供SEA-G4F2模型对单个环境中的结果枝数等4个农艺性状进行了主基因+多基因遗传分析。认为结果枝数受2对主基因+多基因控制,以主基因遗传为主。本研究在上述对单个环境中的结果枝数性状进行主基因+多基因遗传分析的基础上,利用SEA-F2模型对合肥环境(E1)和固镇环境(E2)中的远杂9102/皖花4号F2群体的结果枝数、单株结果数和百果质量3个产量性状进行遗传分析,以期获得对上述3个花生产量性状遗传方式更为全面的认识。
1 材料和方法
1.1 试验材料
供试材料为远杂 9102(P1)、皖花 4 号(P2)以及由P1和P2杂交自交获得的F2群体。安徽省农业科学院院内基地(合肥,E1)中的F2群体由133个单株组成,固镇县花生原种场(固镇,E2)中的F2群体由151个单株组成。E1位于江淮之间,北纬31°86′东经 117°29′,马肝泥田,黏性大,湿时黏韧,干时坚硬。花生生长季节雨量充沛。E2位于安徽省东北部,淮河中游北岸,北纬 33°32′,东经 117°32′,砂浆黑土,保水性差。花生生长季节阳光充足。
1.2 试验设计
2014年将P1、P2和F2分别种于安徽省农业科学院院内基地(E1)和固镇县花生原种场(E2)。做畦,畦宽1.5 m,分别于5月29日(E1)和6月20日(E2)人工单粒点播,P1和P2种植2行,F2全部种完,所有材料均是每行10穴,行距40.0 cm,常规栽培管理。
1.3 测定项目与方法
花生成熟后,P1、P2选择长势良好的10个单株、F2群体所有单株参照《花生种质资源描述规范和数据标准》进行结果枝数、单株结果数和百果质量3个产量性状的调查与记录[11]。
1.4 数据分析
F2群体3个产量性状的次数分布按照盖钧镒的方法进行整理[12],采用曹锡文等研制的植物数量性状分离分析软件中的SEA-F2模型对远杂9102/皖花4号的P1、P2和F2群体进行遗传分析。运算结果中AIC值最小的模型即为该性状的最适模型,统计最适模型下的遗传参数[13]。
2 结果与分析
2.1 P1、P2和F2群体3个产量性状的次数分布及其遗传分析
表1表明,E1中远杂9102平均结果枝数9.20±0.92个,皖花4号平均结果枝数7.60±0.97个,F2群体平均结果枝数8.45±3.31个,接近于双亲均值;E2中远杂9102平均结果枝数16.50±3.17个,皖花4号平均结果枝数7.60±1.84个,F2群体平均结果枝数7.94±2.38个,介于双亲之间,接近于低值亲本。2个环境中的F2群体均存在结果枝数多于高值亲本和少于低值亲本的株系,表现连续分布且多峰,说明该组合结果枝数发生了明显的超亲分离。单株结果数和百果重性状在2个环境中的表现与结果枝数性状类似。
以5类24种模型配合表1结果枝数分布求出各种遗传模型的极大似然值和AIC值,AIC值最小的模型为最适模型。结果发现2个环境中的结果枝数性状均是1MG-AD模型的AIC值最小,分别为692.27(E1)和 695.33(E2),说明结果枝数性状在远杂9102/皖花4号组合中表现为受1对加性-显性主基因控制,无多基因效应。
由表2可知,单株总果数性状在E1中2MGAD模型的AIC值(1 232.36),说明E1中该性状在远杂9102/皖花4号组合中表现为受2对加性-显性主基因控制,无多基因效应;而在E2中单株总果数性状1MG-AD模型的AIC值(1 286.20)最小,说明E2中该性状在远杂9102/皖花4号组合中表现为受1对加性-显性主基因控制,无多基因效应。
由表3可知,百果质量性状2个环境中均是2MG-EEAD模型的AIC值最小,分别为1 327.30(E1)和 1 477.27(E2),说明该性状在远杂 9102/皖花4号组合中表现为受2对等显性主基因控制,无多基因效应。

表1 远杂9102/皖花4号组合F2群体结果枝数的次数分布
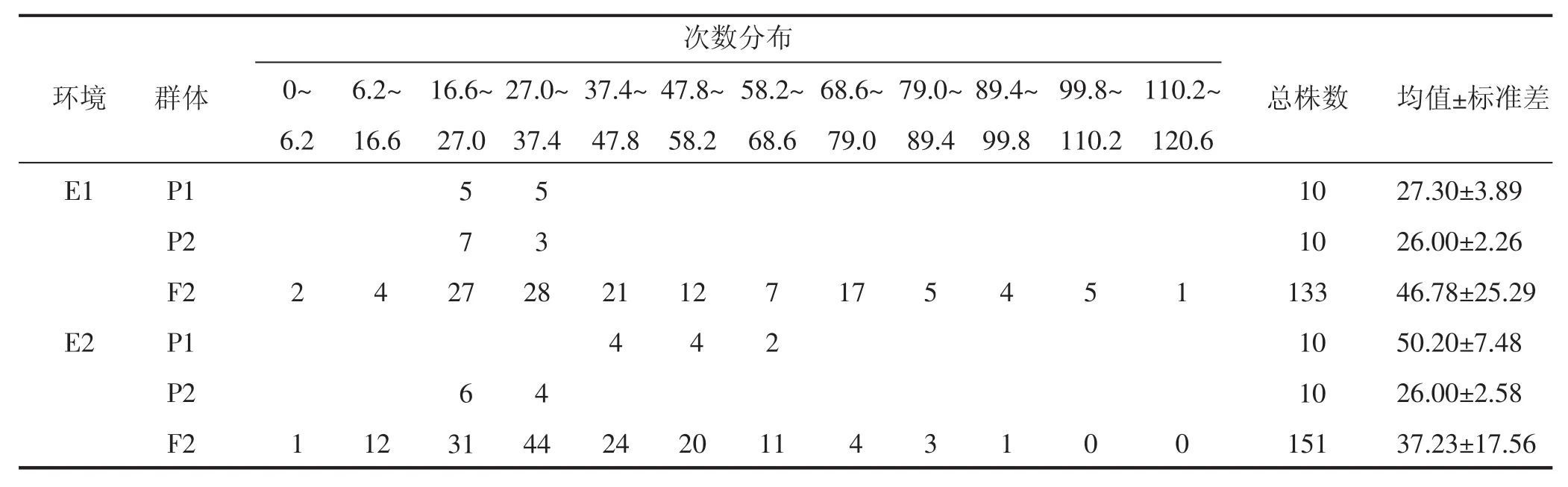
表2 远杂9102/皖花4号组合F2群体单株总果数的次数分布

表3 远杂9102/皖花4号组合F2群体百果质量的次数分布
2.2 3个产量性状的遗传参数估计
表4中的一阶参数显示,结果枝数性状在E1中的|[da(d)]|=|[ha(h)]|,而在E2中的|[da(d)]|略大于|[ha(h)]|,说明控制结果枝数性状中的加性效应与显性效应并重。单株总果数性状E1中的|[da(d)]|+|[db]|略小于|[ha(h)]|+|[hb]|,而E2中的|[da(d)]|略大于|[ha(h)]|,说明控制单株总果数性状中的遗传效应与结果枝数性状基本一致,均呈现出加性效应与显性效应并重。2个环境中百果重性状的|[da(d)]|分别为7.18(E1)和16.12(E2),没有检测到显性效应,说明百果重性状以加性效应为主。
从表4中的二阶参数可知,结果枝数性状在2个环境中的主基因遗传率分别为55.01%(E1)和37.26%(E1);单株总果数性状在2个环境中的主基因遗传率分别为 83.19%(E1)和 56.36%(E1);百果质量性状2个环境中的主基因遗传率分别为43.69%(E1)和 38.77%(E1)。3个性状在 2个环境中均没有检测到多基因效应。

表4 远杂9102/皖花4号F2群体3个产量性状遗传参数的估计值
3 讨论与结论
本研究利用远杂9102/皖花4号F2群体对2个环境中3个产量性状进行了主基因+多基因遗传分析,发现结果枝数性状在2个环境下均表现出受1对加性-显性主基因控制;单株总果数性状不同环境中表现不一,E1中表现出受2对加性-显性主基因控制,而在E2中则表现出受1对加性-显性主基因控制;百果重性状在2个环境下均表现出受2对等显性主基因控制。3个性状在2个环境中均没有检测到多基因效应。笔者前期[10]曾以本研究中的P1、P2、F1和F2 4个世代为研究材料,利用主基因+多基因模型中的SEA-G4F2对单个环境中的结果枝数进行了遗传分析,认为结果枝数性状受2对加性主基因+多基因控制。这与本研究结果既有共同点也有差异,相同的是两者都认为结果枝数性状受主基因控制,加性效应和显性并重;不同的是本研究中认为结果枝数性状受1对主基因控制,不存在多基因效应,这与笔者[10]前期认为的该性状受2对主基因加多基因共同控制不尽相同,这与张新友等[9]认为的结果枝数性状受3对独立的等加性主基因控制,无多基因效应的结果也是不同的。同时张新友等[9]认为百果重性状受2对主基因加多基因控制,这与本研究检测到的控制该性状的主基因对数是一致的,但2对基因之间的作用不同。上述结果表明对于同一个性状,不同研究者所用研究材料不同以及相同材料不同世代所得出的研究结果是不尽相同的。因此,育种实践中,育种者要根据不同的育种材料/组合不同世代采用不同的育种方法对相关性状进行遗传改良。
参考文献:
[1]张晓杰,姜慧芳,任小平,等.中国花生核心种质的主成分分析及相关分析[J].中国油料作物学报,2009,31(3):298-304.
[2]江建华,倪皖莉,于欢欢,等.花生单株生产力与主要农艺性状间的相关性研究[J].中国农学通报,2013,29(36):125-130.
[3]殷冬梅,李拴柱,崔党群.花生主要农艺性状的相关性及聚类分析[J].中国油料作物学报,2010,32(2):212-216.
[4]李清华,黄金堂,陈海玲.花生数量性状的多元遗传分析[J].江西农业学报,2009,21(11):7-9.
[5]Zhang X Y,Zhu S J,Han S Y,et al.Inheritance of Fat and Fatty Acid Compositions in Peanut(Arachis hypogaea L.)[J].Agricultural Science&Technology,2011,12(7):943-946,974.
[6]黄冰艳,张新友,苗利娟,等.花生油酸和亚油酸含量的遗传模式分析[J].中国农业科学,2012,45(4):617-624.
[7]胡晓辉,苗华荣,杨伟强,等.花生种子休眠性的遗传分析及影响因素的研究[J].核农学报,2013,27(10):1449-1455.
[8]刘 华,张新友,韩锁义,等.花生主茎高,侧枝长的遗传分析及 QTL 检测[J].中国油料作物学报,2013,35(5):508-514.
[9]张新友.栽培花生产量品质和抗病性的遗传分析与QTL定位研究[D].杭州:浙江大学,2010:76-77.
[10]江建华,王 嵩,汪 清,等.花生4个主要农艺性状的遗传分析[J].花生学报,2015,44(3):7-13.
[11]姜慧芳,段乃雄.花生种质资源描述规范和数据标准[M].北京:中国农业出版社,2006.
[12]盖钧镒.试验统计方法(第3版)[M].北京:中国农业出版社,2000:99-126,157-189.
[13]曹锡文,刘 兵,章元明.植物数量性状分离分析Windows软件包 SEA 的研制[J].南京农业大学学报,2013,36(6):1-6.
