ZmBK2L3在番茄果实中的特异表达及其功能研究
2016-04-22杨潇怡唐晓凤曹徐绿岳俊阳刘永胜
杨潇怡, 隋 媛, 唐晓凤, 曹徐绿, 岳俊阳, 刘永胜
(合肥工业大学 生物与食品工程学院,安徽 合肥 230009)
ZmBK2L3在番茄果实中的特异表达及其功能研究
杨潇怡,隋媛,唐晓凤,曹徐绿,岳俊阳,刘永胜
(合肥工业大学 生物与食品工程学院,安徽 合肥230009)
摘要:文章通过构建玉米BK2L3基因果实特异表达的过量表达载体,对转基因番茄果实进行研究,实现以基因工程手段延长番茄果实货架期的目的。利用逆转录聚合酶链式反应(reverse transcription polymerase chain reaction,RT-PCR)技术从玉米cDNA中扩增出ZmBK2L3基因全长,导入启动子为果实特异表达的TFM7植物表达载体pBI121上,经农杆菌介导转入野生型番茄中,获得转基因阳性植株。分别以野生型番茄和转基因番茄为材料,分析BK2L3在果实中的表达情况,观察绿熟期果实果皮细胞壁结构,测定红熟期果实的硬度、货架期和果皮纤维素质量比。结果表明:转基因番茄果实中BK2L3的表达量明显高于野生型果实。转基因番茄果实的硬度、果皮纤维素质量比和果皮厚度均比野生型有所提高,果实货架期明显增长。因此,BK2L3基因的过量表达与细胞壁代谢和货架期有关,可改良果实的品质。
关键词:BK2L3基因;货架期;过量表达;果实特异表达;转基因番茄;遗传转化
COBRA基因影响植物细胞壁纤维素和微纤丝的正确定位,对细胞的定向伸长起关键作用[1]。COBRA基因的突变体导致正在伸长的根部组织细胞性状异常和纤维素含量的明显减低[2]。目前,已有多个COBRA基因突变体的研究证实,纤维素含量的降低引起明显的易脆表型[3-7]。COBRA基因家族在玉米中有9个成员[8]。通过对COBRA基因在植物中的表达分析,ZmBK2L3属于玉米ZmBK2L家族成员,与拟南芥AtCOB具有很高的同源性,其编码的蛋白也含有CCVS保守基序、N端的跨膜信号肽、 C-末端的疏水性尾部以及GPI锚定ω-位点[9]。ZmBK2L3在玉米发育的各个时期与各个器官中均有表达,尤其是在根系组织、叶片组织、外果皮和营养器官中有较高表达量,ZmBK2L3参与了纤维素合成相关基因的调控[10]。
外源基因在其自身不表达的同时也造成与之同源的内源基因的沉默的现象称为同源共抑制现象[11],高度转录的外源基因产生的紊乱RNA引发了RNA干涉机制。TFM7果实特异性SlCOBRA-like-OE转基因植株,在生长至绿果时期时,其果实出现了裂果表型[9]。文献[10]指出拟南芥COBRA基因会发生100%同源共抑制。
果蔬的运输、贮藏、采后处理、货架期的延长等一直是食品保鲜技术关注的问题。番茄果实硬度是衡量果实货架寿命长短的重要指标,而且可溶性果胶的含量直接影响番茄果实的硬度[12]。因此,本研究通过构建TFM7启动子的果实特异性表达载体,将外源的玉米ZmBK2L3基因转入野生型番茄,有效避免转基因植株的同源共抑制现象,不产生裂果表型,实现促进基因的高效稳定表达、减少株系差异的目的,为反向遗传学的进一步研究提供有价值的参考信息。通过转基因技术可有效提高植物细胞壁中的纤维素质量比和果实硬度,延长货架期。同时,通过制作石蜡切片,观察果皮细胞壁结构,为玉米ZmBK2L3基因参与细胞壁的生物合成假说提供证据。转基因番茄的营养品质良好,实现了果实品质的改良。这为进一步使用基因工程手段解决食品贮藏问题提供了新思路。
1材料与方法
1.1材料
野生型番茄(Solanumlycopersicum)来自美国康乃尔大学THOMPSON植物研究所;大肠杆菌DH5α菌株、农杆菌EH105、番茄果实特异表达载体pBI121-TFM7均为本实验室保存;感受态细胞Trans1-T1 Chemically Competent Cell、克隆载体pEASY-Blunt Simple Cloning Kit均购于TaKaRa公司。
Trizol购自Invitrogen公司;反转录酶、Taq DNA聚合酶、限制性内切酶和T4 DNA连接酶购于TaKaRa公司;反转录试剂盒购于Transgen公司;质粒提取试剂盒、琼脂糖凝胶回收试剂盒购自TIANGEN公司;聚合链式反应(polymerase chain reaction,PCR)引物由南京金斯瑞公司合成,测序由南京金斯瑞公司完成;组培试剂购于Sigma公司;其余试剂均为进口分装或国产分析纯产品。
1.2方法
1.2.1ZmBK2L3目的基因的克隆
按照GenBank中玉米ZmBK2L3序列(登录号EF078698.1)设计全长引物BK2L3-F1和BK2L3-R1及BK2L3-F2和BK2L3-R2引物具体序列如下。
BK2L3-F1:GCGGCTCTGTATCTATCTGTCG。
BK2L3-R1:CAACTTGAAACTTGCTAAACCTCT。
BK2L3-F2:TCTAGAATGGCGGCGAGCGGCA,酶切位点在TCTAGA(XbaⅠ)。
BK2L3-R2:GAGCTCCACTAATCATGCATAAGCC
AACAGAGC,酶切位点在GAGCTC(SacⅠ)。
NPTⅡ-F:AGACAATCGGCTGCTCTGAT。
NPTⅡ-R:TCATTTCGAACCCCAGAGTC。
ACTIN-F:CGAGCAGTGTTTCCCAGTATT。
ACTIN-R:AGCCTGGATAGCAACATACATAG。
SLBK2L3-F:CTCCATCCTGCTGTGTATCTCT。
SLBK2L3-R:CGTATCATTTATGCCACCACC。
以野生型玉米cDNA为模板,通过巢式PCR扩增ZmBK2L3基因全长编码序列。其中,PCR反应条件为:94 ℃预变性5 min;94 ℃变性30 s,58 ℃退火30 s,72 ℃延伸80 s,29个循环; 72 ℃延伸5 min。然后将PCR产物与pEASY-Blunt载体连接后转化进入大肠杆菌并进行PCR验证,最后阳性克隆送至南京金斯瑞公司测序。
1.2.2ZmBK2L3果实特异过表达载体构建
参照载体pBI121-TFM7的酶切位点,采用Sequencher软件对ZmBK2L3基因序列进行限制性内切酶分析,运用Primer Premier5软件设计含有酶切位点的特异引物。将已测序正确的ZmBK2L3全长序列进行XbaⅠ和SacⅠ双酶切后插入植物表达载体的多克隆位点区域,构建成番茄果实特异的pBI121-TFM7-BK2L3OE过表达载体。将得到的连接产物转化进入大肠杆菌感受态细胞,筛选阳性菌落,提取质粒酶切验证。
1.2.3番茄的遗传转化及转基因植株的鉴定
将重组质粒pBI121-TFM7-BK2L3-OE通过冻融法导入农杆菌EH105,在含有利福平和卡那霉素双抗生素的培养基上,28 ℃暗培养2 d,筛选阳性菌用以转化野生型番茄。
将野生型番茄的种子用自来水在37 ℃浸泡12 h,然后用20%次氯酸钠溶液浸泡15 min,再经过无菌水漂洗10次后播种至1/2 MS培养基。25 ℃暗培养4 d,露白后转至光照条件下培养,待子叶展平,剪下置于预培养基上培养3 d,接下来用带有目的基因的农杆菌(经28 ℃过夜培养)侵染子叶10~15 min后转入共培养基上培养2 d。经过不同梯度抗性筛选,培育出转基因材料,最后移入温室土壤中栽培。
以转基因番茄幼苗叶片为材料,提取番茄基因组DNA。根据pBI121载体筛选标记基因新霉素磷酸转移酶基因NPTⅡ(登录号:AF485783)的序列合成引物NPTⅡ-F和NPTⅡ-R,进行PCR鉴定。PCR反应条件:94 ℃预变性5 min;94 ℃变性30 s;64 ℃退火30 s,72 ℃延伸45 s,29个循环;72 ℃延伸5 min。
1.2.4半定量RT-PCR分析
分别提取野生型和转基因T1代番茄绿熟期果实总RNA,经DNase处理,反转录合成第1条cDNA,以此为模板进行半定量逆转录聚合酶链式反应(reverse transcription polymerase chain reaction,RT-PCR)检测。以番茄ACTIN(登录号:AB199316.1)作为内参基因,设计BK2L3基因半定量引物。反应条件:94 ℃预变性5 min;94 ℃变性30 s;64 ℃退火30 s,72 ℃延伸80 s,24个循环;72 ℃延伸5 min。
1.2.5石蜡切片实验
分别取野生型和转基因T1代番茄绿熟期果实,选取果实赤道部位的果皮,经FAA固定、脱水至透明,石蜡包埋后,用石蜡切片机切片,制得厚度为10 μm的石蜡切片,固绿染色后,在光学显微镜下观察果皮细胞壁结构。
1.2.6果皮细胞壁成分提取及分析
分别取野生型和转基因T1代番茄绿熟期果实,称15 g果皮在液氮中充分研磨,经70%乙醇、氯仿甲醇混合液(体积比为1∶1)、丙酮、90% DMSO依次浸提,离心,去上清,用丙酮清洗2次后在37 ℃烘箱烘干至恒质量,保存于真空干燥器中[13]。
将提取的果皮细胞壁成分先经酸性水解,再进行浓硫酸脱水,在620 nm下对其与蒽酮结合反应后产物进行比色,测定细胞壁纤维素质量比。
1.2.7果皮质构分析及货架期实验
分别取野生型和转基因T1代红熟期番茄果实,选取果实赤道部位三等分点,利用物性仪TA-XT Plus检测果皮硬度。选用P/2N探头,以2 mm/s测速,50%应力,进行穿刺实验[10]。
分别取野生型和转基因T1代红熟期番茄果实,于25 ℃温度、60%~65%湿度条件下贮存。每隔5 d进行称质量,计算果实失水率。
2结果与分析
2.1ZmBK2L3基因的扩增和测序
经以BK2L3-F1和BK2L3-R1为引物的PCR扩增产物凝胶电泳检测,扩增产物约为1 400 bp的特异条带,结果如图1所示,由图1可看出,与预期的玉米ZmBK2L3基因片段大小一致。测序结果经比对,与GenBank的序列一致。

M—DNA marker DL 2000 plus 1.BK2L3 PCR扩增产物
2.2ZmBK2L3-OE载体鉴定
将载体pEASY-Blunt-BK2L3-OE和pBI121-TFM7-BK2L3-OE分别用XbaⅠ、SacⅠ双酶切,所得到的片段大小均为1 400 bp左右,结果如图2所示,图2证明了获得pBI121-TFM7-BK2L3-OE重组载体。
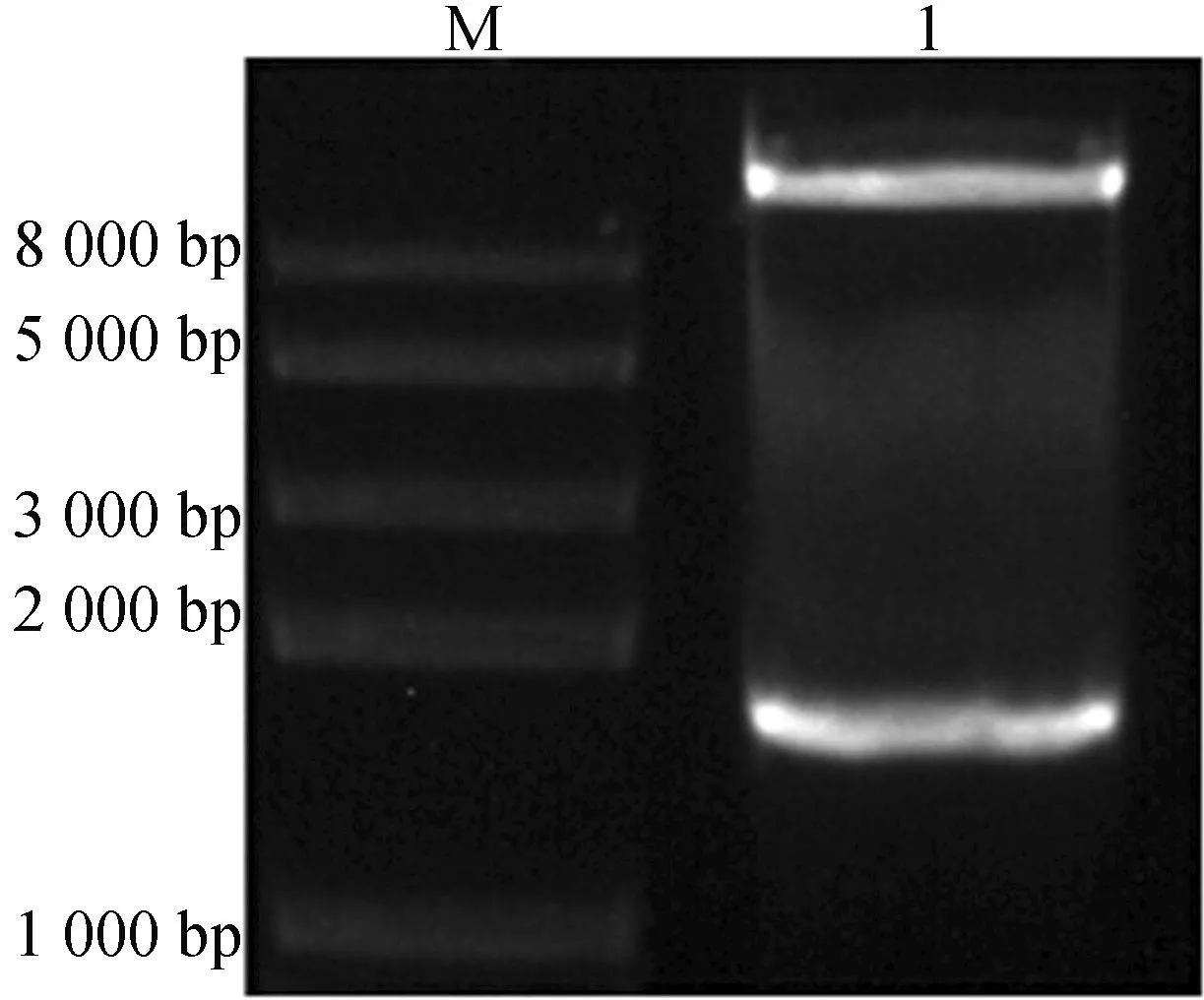
M—DL 2000 plus 1.XbaⅠ和SacⅠ双酶切
2.3转基因植株阳性鉴定
以番茄叶片为材料提取基因组DNA,利用NPTⅡ引物进行PCR鉴定,结果如图3所示。图3表明,转基因植株扩增出857 bp的条带,而阴性对照野生型植株未扩增出857 bp的条带,说明NPTⅡ基因已整合于植株核染色体组中。
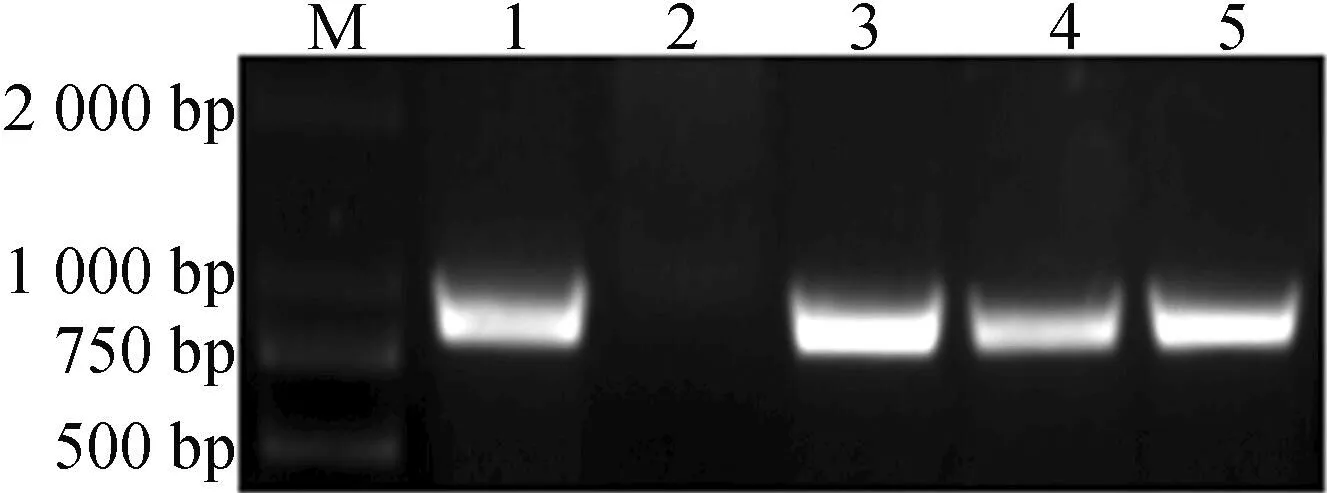
M—DL 2000 plus 1.阳性对照
2.4ZmBK2L3表达分析
分别提取野生型和转基因番茄绿熟期果实的RNA,利用半定量RT-PCR方法,分析转基因植株中ZmBK2L3基因的表达水平,结果如图4所示。由图4可知,ZmBK2L3基因在野生型番茄果实中无表达,而在ZmBK2L3-OE转基因番茄果实中有较好表达;同时,不同株系的转基因果实表达量有差异。

WT—野生型 1~3. 转基因
2.5石蜡切片实验
分别取野生型和转基因T1代番茄绿熟期果实,选取果实赤道部位的果皮,制得石蜡切片后,在光学显微镜下观察果皮细胞壁结构,结果如图5所示。

图5 番茄果皮纤维结构
由图5可看出,绿熟期转基因番茄果实的果皮厚度比野生型明显增加,而且转基因番茄中果皮细胞大小均匀。
2.6细胞壁纤维素质量比
提取番茄细胞壁纤维素成分后,每株系取红熟期番茄果实5个,共3个株系,3次重复测定果实细胞壁纤维素的质量比,结果如图6所示。由图6可看出,转基因番茄细胞壁纤维素的质量比野生型提高约42%。
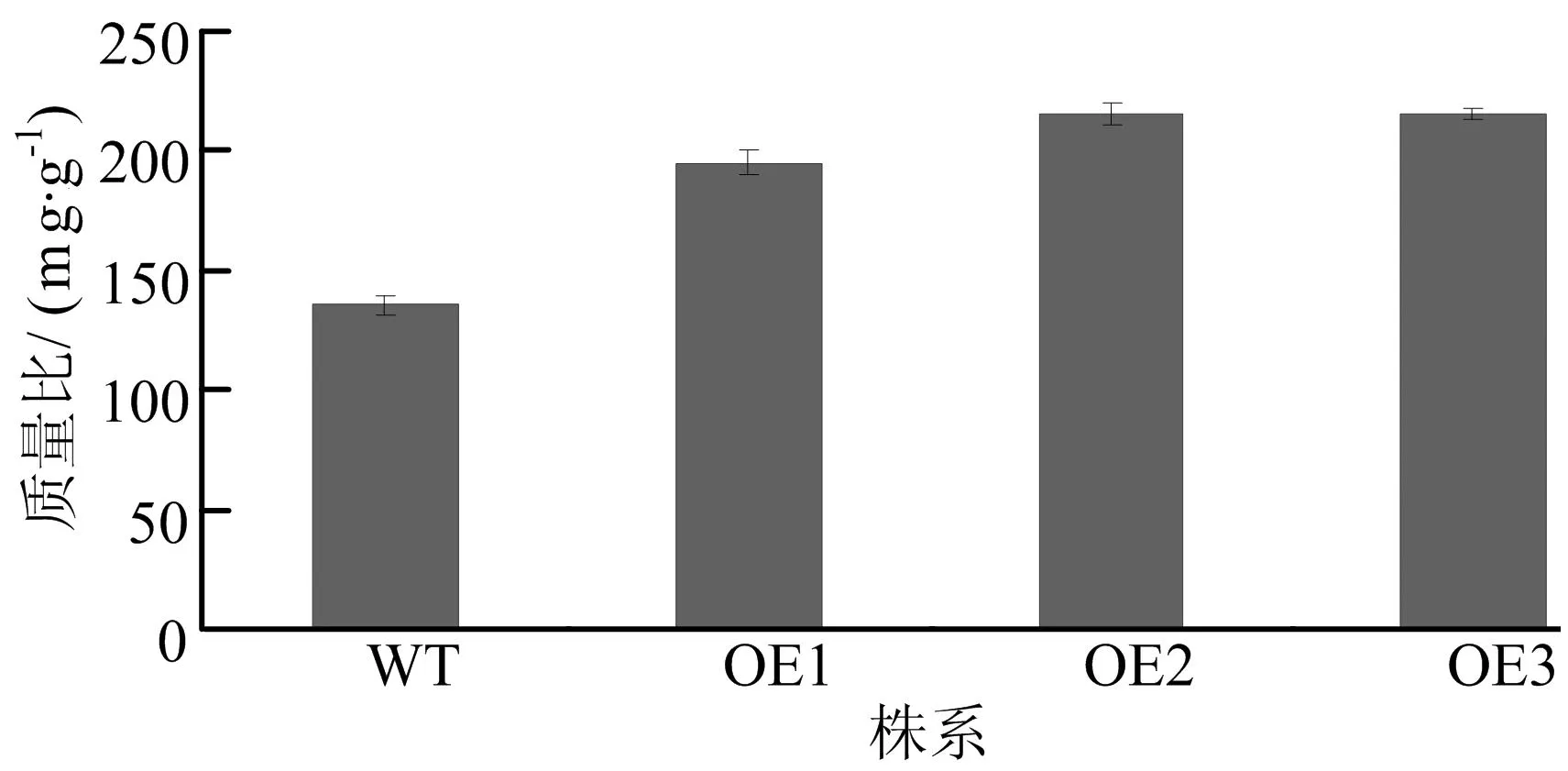
WT—野生型 OE1~OE3.转基因
2.7转基因番茄果皮硬度、细胞壁结构和货架期
每个株系取红熟期番茄果实5个,共3个株系,测定果实硬度,每个果实赤道面三等分位置取平均值,结果如图7所示,转基因番茄果皮硬度比野生型高约48%。

WT—野生型 OE1~OE3.转基因
每一个株系取红熟期番茄果实9个,共3个株系,失水率结果及果实照片如图8所示。


(b)
由图8a可知,转基因番茄干质量变化较小,野生型果实在20 d后失水率显著增加,果皮开始皱缩。40 d后番茄果实照片如图8b所示,野生型果实已发生完全形变,水分严重损失,而转基因果实仍形态完好。
3讨论
玉米ZmBK2L3基因属于COBRA基因家族,广泛涉及调控植物生长发育以及果实成熟和采后病原体易感性。表达模式分析结果表明,COBRA基因在幼嫩果实组织中表达量较高,随果实成熟度增加,其表达量急剧下降[2]。本文通过构建转基因载体并转入番茄组织,分析BK2L3在果实中的表达水平后,证实与果实的成熟软化有一定相关性。本研究以3个不同的转基因番茄株系为主要研究对象,较好地验证了基因的表达效果。转基因番茄果实纤维素质量比明显增高,果实硬度增加,因此赋予果实更好的坚韧度,有效延长货架期。
与前人报道的番茄COBRA-like基因的功能性基本一致。这一转基因技术应用于食品行业,实现了以基因工程为手段,改善番茄果实品质,创新食品保鲜技术。本文采用的外源基因转化方式可消除同源共抑制现象,有效避免RNA干涉机制引发的内源基因沉默,从而促进基因高效稳定表达,减少转基因株系间差异,为遗传学研究提供了新思路。
[参考文献]
[1]李玲,赖钟雄,陈裕坤,等.文心兰OnCOBRA基因克隆及表达模式分析[J].热带作物学报,2014,35(8):1551-1558.
[2]曹颖,唐晓凤,刘永胜,等.番茄COBRA基因克隆、表达模式及生物信息学分析[J].植物研究,2012,32(3):304-310.
[3]Schindelman G,Morikami A,Jung J,et al.COBRAencodes a putative GPI-anchored protein, which is polarly localized and necessary for oriented cell expansion inArabidopsis[J].Genes & Development,2001,15(9):1115-1127.
[4]Brummell D A,Harpster M H.Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants [J].Plant Mol Biol,2001,47(1):311-340.
[5]Smith D L,Gross K C.A family of at least seven β-galactosidase genes is expressed during tomato fruit development [J].Plant Physiol,2000,123:1173-1183.
[6]Roudier F,Femandez A G,Fujita M,et al.COBRA, anArabidopsisexreacellular glycosyl-phosphatidyl inositol-anchored protein, specifically controls highly anisotropic expansion through its involvement in cellulose microfibrill orientation [J]. The Plant Cell,2005,17(6):1749-1763.
[7]Sindhu A,Langewisch T,Olek A,et al. Maize brittle stalk 2 encodes aCOBRA-like protein expressed in early organ developmeng but required for tissue flexibility at maturity [J]. Plant Physiol,2007,145(4):1444-1459.
[8]高志民,陈颖,胡陶,等.绿竹BoCOBL基因的分子特征及其表达分析[J].热带亚热带植物学报,2013,21(6):560-565.
[9]Cao Ying,Tang Xiaofeng, Giovannoni J,et al.Functional characterization of a tomatoCOBRA-like gene functioning in fruit development and ripening [J].BMC Plant Biology,2012,12:211.
[10]Brady S M,Shuang S,Dhugga K S,et al.Combining expression and comparative evolutionary analysis:theCOBRAgene family [J]. Plant Physiol,2007,143(1):172-187.
[11]孔莹莹,蒋丽,韩凝,等.植物转基因中同源共抑制的机制及其解决措施[J].生命科学,2012,24(5):399-402.
[12]魏宝东,姜炳义,冯辉.番茄果实货架期硬度变化及其影响因素的研究[J].食品科学,2005,26(3):249-252.
[13]Updegraff D M.Semimicro determination of cellulose in biological materials [J].Analytical Biochemistry,1969,32:420-424.
(责任编辑闫杏丽)
Fruit-specific gene expression ofZmBK2L3 in tomato and its function
(School of Biotechnology and Food Engineering, Hefei University of Technology, Hefei 230009, China)
Abstract:Fruit-specific overexpression vector of maize BK2L3 was constructed to investigate the effect of gene engineering on fruit firmness and shelf life. The full-length cDNA sequence of maize ZmBK2L3 gene was amplified by reverse transcription polymerase chain reaction(RT-PCR) and pBI121-BK2L3 overexpression construction driven by a fruit-specific promoter TFM7 was made to generate Agrobacerium mediated tomato transformants. In view of the transgenic tomatoes and the wild type tomatoes, the BK2L3 expression in the fruit was analyzed, the pericarp cell wall structure of fruit at green ripe stage was observed, and the firmness, cellulose content and shelf life of fruit at red ripe stage were determined. The result of molecular analysis demonstrated a significant increase of BK2L3 expression in the transgenic tomatoes. The positive transgenic tomatoes displayed enhanced pericarp thickness. The firmness, cellulose content and shelf life of the transgenic red ripe fruits were higher than those of the wild type species. These results demonstrated that fruit-specific overexpression of maize BK2L3 in tomato could improve fruit quality in terms of cell wall metabolism and shelf life.
Key words:BK2L3 gene; shelf life; overexpression; fruit-specific expression; transgenic tomato; genetic transformation
中图分类号:S641.2;Q786
文献标识码:A
文章编号:1003-5060(2016)03-0410-05
doi:10.3969/j.issn.1003-5060.2016.03.023
作者简介:杨潇怡(1988-),女,安徽合肥人,合肥工业大学硕士生;刘永胜(1964-),男,重庆市人,博士,合肥工业大学教授,博士生导师.
基金项目:国家自然科学基金资助项目(31171179)
收稿日期:2015-01-12;修回日期:2015-03-27
