增温和CO2浓度加倍对川西亚高山针叶林土壤可溶性氮的影响
2016-04-13刘芙蓉张咏梅邓书林
刘芙蓉, 张咏梅, 邓书林
1 中国科学院成都生物研究所, 成都 610041
2 中国科学院大学, 北京 100049
增温和CO2浓度加倍对川西亚高山针叶林土壤可溶性氮的影响
刘芙蓉1,2, 张咏梅1,*, 邓书林1,2
1 中国科学院成都生物研究所, 成都610041
2 中国科学院大学, 北京100049
摘要:采用全自动微气候控制的“人工模拟气候实验系统”研究了增温和CO2浓度加倍对川西亚高山针叶林土壤硝态氮-N)、铵态氮-N)、游离氨基酸(FAA)、可溶性有机氮(DON)和可溶性总氮(TSN)的影响。结果表明:①在种植油松苗木组,增温处理显著降低了土壤-N含量,不同处理0—15 cm土层-N含量均显著小于15—30 cm层;而在未种树组,增温处理显著增加了土壤-N含量, 0—15 cm土层-N含量显著高于15—30 cm层,这表明增温促进了油松苗对-N的吸收。②在种植油松苗木组,增温(ET)、增CO2(EC)及两者的共同作用(ETC)均显著增加了土壤-N、DON和TSN含量;在未种树组,ET显著增加了土壤-N、FAA、DON和TSN含量,EC和ETC对、FAA、DON和TSN含量具有微弱影响或没有显著影响。不同处理0—15cm层土壤-N、FAA、DON和TSN的含量显著大于15—30 cm层。③种植油松苗木组土壤-N、FAA、DON和TSN含量均显著低于未种树组,这是由植物对氮素的吸收消耗造成的。研究结果表明,EC、ETC主要通过植物根系作用促进了-N、DON和TSN含量增加,而ET处理通过影响土壤微生物和植物根系来促进-N、FAA、DON和TSN含量的增加。
关键词:增温; 增CO2; 硝态氮; 铵态氮; 游离氨基酸; 可溶性有机氮; 可溶性总氮
氮素作为植物生长的主要营养元素之一,在生态系统养分循环过程中起着重要作用[1]。曾经一些研究者认为自然生态系统中植物吸收的氮主要来自于无机氮,而有机态氮素难以被植物吸收利用,需要在微生物和土壤动物的作用下,通过氮矿化过程转化成无机氮后才能被植物吸收利用[2]。森林生态系统中氮绝大部分以有机氮的形式存在,可被植物和微生物吸收利用的土壤矿质态氮含量一般低于土壤总氮的1%,因此土壤氮素转化是森林生态系统氮素循环的重要组成部分,氮矿化作用是氮循环的核心过程。而近年来有一些研究报道,部分植物还可以利用小分子有机态氮[3- 6],这一研究结果受到越来越多生态学家和土壤学家的关注。由于无机氮在植物体内储存需要较高的维持性消耗,而以氨基酸或蛋白质形式储存的有机氮更有利于植物的氮素储备,当外界氮的供应降低时植物很容易利用这部分有机氮[7],因此可溶性有机氮与无机氮共同影响着植物氮素的营养平衡[8]。目前,国内外学者在土壤碳循环对气候变化响应的研究较多,但在可溶性氮响应方面的研究还相当缺乏。
森林土壤氮素转化十分复杂,其含量往往受到温度、湿度、凋落物化学组成、土壤pH值、土壤养分状况等因子调控。增温和CO2浓度升高对生态系统的影响是全球气候变化研究的重要内容,政府间气候变化专门委员会(IPCC)评估报告指出:自工业化时代以来,人类活动已引起全球温室气体排放增加,破环了气候系统的平衡,预测在本世纪末全球平均气温将上升1.4—5.8 ℃,大气CO2浓度也将达到490—1260 μL/L[9]。川西亚高山针叶林是青藏高原东缘高寒林区的重要组成部分,对全球气候变化十分敏感[10]。近年来,已有许多学者在该区森林生态系统对全球变暖的响应方面进行了模拟,但大多研究主要集中在模拟气候变化对植物生理、凋落物分解和碳循环的影响方面[11- 16],对土壤可溶性氮的响应研究较少。油松作为川西地区典型的树种之一,在植被恢复过程中发挥着重要的生态功能[17]。因此,本文运用全自动微气候控制的“人工模拟气候实验系统”对川西亚高山油松林下土壤可溶性氮进行研究,探索增温与增CO2对土壤可溶性氮的影响,以期揭示全球气候变化背景下青藏高原东缘亚高山针叶林土壤可溶性氮的变化特征,为预测该区森林生态系统对全球气候变暖的响应提供基础数据和理论依据。
1研究地区与研究方法
1.1研究地区概况
研究地点位于四川省阿坝州中国科学院茂县山地森林生态系统定位研究站,海拔1826 m,地理位置为103°54′E,31°42′N。该区地处青藏高原横断山系北段高山峡谷地带的长江支流岷江上游中部,属暖温带气候,年均温为9.3 ℃,年平均降雨量达825.2 mm,年平均蒸发量968.7 mm,年日照时数约1373.8h,无霜期200d左右。该地区是青藏高原东缘和长江上游生态环境的高山峡谷区典型代表,土壤类型主要为棕壤,植被以油松、云杉和冷杉林为主[18]。
1.2实验设置
2010年3月,采集同一油松林原位土壤样品,0—10 cm土层有机碳初始值为9.80 g/kg,全氮为1.16 g/kg。按照林下土壤自然发生层装入96个花盆,(Φ35 cm×35 cm)中,将花盆移入8个“人工模拟气候实验系统”的生长室内,每个生长室内放置12个花盆(花盆分为两组:其中6盆各种植1株5年生油松苗木;另6盆不栽植苗木),即重复数为2个生长室×6个花盆。油松苗平均高25.53 cm,地径3.38 mm,根系深度30 cm。同时,在生长室顶部采用遮阳网覆盖,以控制生长室内的光照强度与采样的自然生长的林下光照强度基本一致(自然光的70%),并通过TDR土壤水分速测仪实时监控保持土壤含水量在30%左右(体积百分含量)。根据IPCC对未来气候变化趋势的预测结果,设置的试验包括8个处理,即:(1)现行环境温度增加(2.5±0.5)℃+CO2浓度加倍+种树处理(ETCP);(2)现行环境温度增加(2.5±0.5)℃+CO2浓度加倍+不种树处理(ETCS);(3)现行环境温度增加(2.5±0.5)℃+种树处理(ETP);(4)现行环境温度增加(2.5±0.5)℃+不种树处理(ETS);(5)现行环境CO2浓度加倍+种树处理(ECP);(6)现行环境CO2浓度加倍+不种树处理(ECS);(7)现行环境温度和CO2浓度+种树处理(CKP);(8)现行环境温度和CO2浓度+不种树处理(CKS)。每年在生长季4—10月采用全自动微气候控制的“人工模拟气候实验系统”对8个独立、自控、封闭的生长室(Chamber)进行控制,生长室底面积9.5 m2,体积约24.5 m3。“人工模拟气候实验系统”的构成和详细控制参数详情见已发表文献[19]。
1.3 土壤样品采集与处理
2012年7月,用土钻按0—15 cm,15—30 cm分层采集花盆中原位土壤,每个处理3盆,去除土样中石块和动植物残体,过2 mm筛后装入布袋,带回实验室,贮存于4℃冰箱,尽快用于硝态氮、铵态氮、游离氨基酸和可溶性有机氮含量的测定,可溶性总氮为硝态氮、铵态氮和可溶性有机氮三者之和。
硝态氮测定:采用双波长紫外分光光度校正因数法,称取土样按土液比1∶5加入去离子水,250 r/min振荡1 h,悬液静置后过滤,测定浸提液在220 nm和275 nm处的吸光度值。
铵态氮测定:采用KCl浸提-靓酚蓝比色法,称取土样放按土液比1∶5加入去离子水,250 r/min振荡1 h,悬液静置后过滤。吸取滤液10 mL,放入50 mL容量瓶中,依次加入5 mL酚溶液和5 mL NaClO碱性溶液,摇匀后在20℃左右室温下放置1 h,加入1 ml掩蔽剂以溶解可能生成的沉淀物,用水定容后在625 nm波长处进行比色。
游离氨基酸测定:采用茚三酮比色法测定。
可溶性有机氮测定:称取土样按土液比1∶5加入去离子水,250 r/min震荡1 h,将土壤上清液过 0.45 um滤膜获得滤液,用德国Elementar Vario TOC 分析仪测定DON 含量。
1.4数据处理
利用SPSS 19进行统计和数据分析,采用多因素方差分析和Duncan法进行差异比较;利用Origin 8.0软件进行图形绘制。
2结果与分析
2.1增温与增CO2对土壤硝态氮含量的影响

图1 增温、增CO2及其共同作用对不同土层土壤含量的影响Fig.1 Effects of ET、EC、ETC on the content of soil at different soil layers.直方柱上不同小写字母表示同一土层不同处理间差异显著 (P < 0.05); ETCP:elevated temperature and CO2 doubling in seeding treatment (增温+增CO2+种树),ECP:CO2 doubling in seeding treatment (增CO2+种树),ETP:elevated temperature in seedling treatment (增温+种树),CKP:ambient temperature and CO2 in seedling treatment (仅种树);ETCS:elevated temperature and CO2 doubling in plant-free treatment (增温+增CO2+不种树),ECS:CO2 doubling in plant-free treatment (增CO2+不种树),ETS:elevated temperature in plant-free treatment (增温+不种树),CKS:ambient temperature and CO2 in plant-free treatment (不种树)
2.2增温与增CO2对土壤铵态氮含量的影响
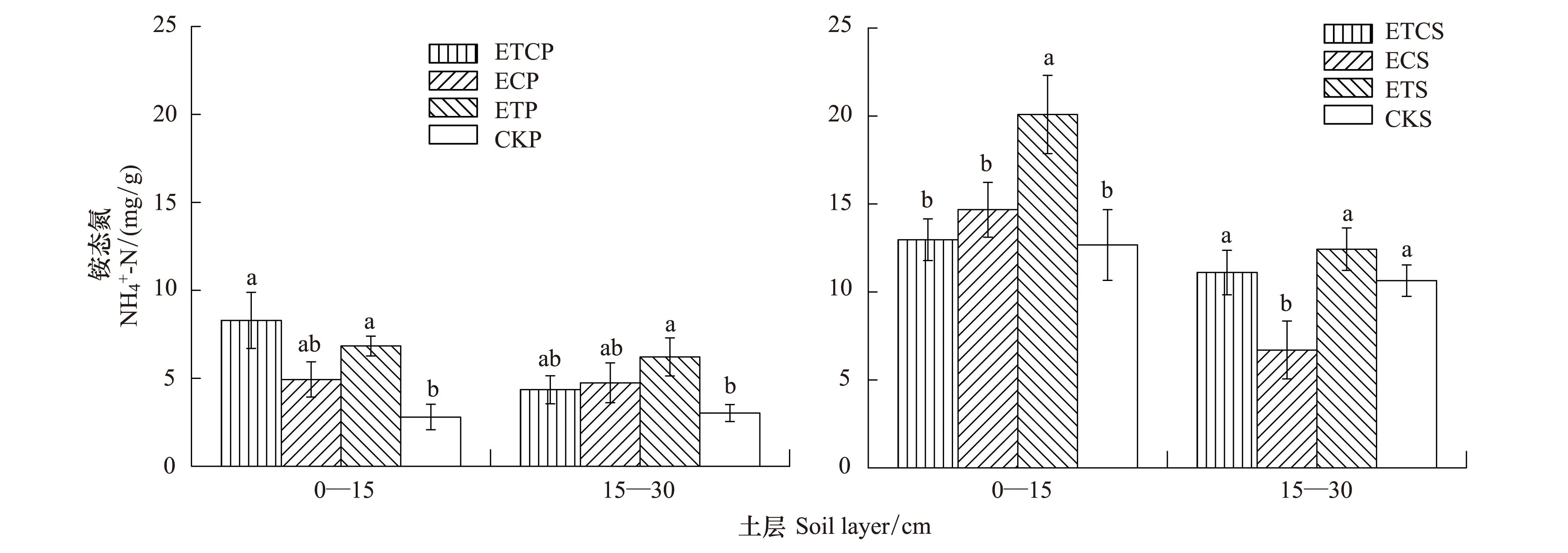
图2 增温、增CO2及其共同作用对不同土层土壤含量的影响Fig.2 Effects of ET、EC、ETC on the content of soil at different soil layers.直方柱上不同小写字母表示同一土层不同处理间差异显著 (P < 0.05)
2.3增温与增CO2对土壤游离氨基酸含量的影响
由图3可知,增温对土壤游离氨基酸含量产生了显著影响,种植油松苗木组土壤游离氨基酸含量显著低于未种树组,且不同处理下 0—15 cm层土壤游离氨基酸含量显著高于15—30cm土层。种植油松苗木组,不同处理下土壤游离氨基酸含量为0.09—0.19 mg/kg,与CKP相比,0—15 cm土层ETP处理显著增加了土壤游离氨基酸含量,而ETCP、ECP、CKP之间差异不显著;15—30 cm层,不同处理间土壤游离氨基酸差异均不显著。
未种树组,0—15 cm土层,不同处理下土壤游离氨基酸含量为0.20—0.38 mg/kg,ETS显著增加了土壤游离氨基酸含量,而ETCS、ECS与CKS均没有显著差异;15—30 cm土层,不同处理的土壤游离氨基酸含量为0.07—0.29 mg/kg,ETS显著增加了土壤游离氨基酸含量。
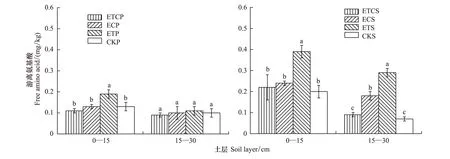
图3 增温、增CO2及其共同作用对不同土层土壤游离氨基酸含量的影响Fig.3 Effects of ET、EC and ETC on the content of soil free amino acids (FAA) at different soil layers直方柱上不同小写字母表示同一土层不同处理间差异显著 (P < 0. 05)
2.4增温与增CO2对可溶性有机氮含量的影响
由图4可知,增温处理对DON含量产生了显著影响,种植油松苗木组土壤DON含量显著低于未种树组,且0—15 cm层土壤DON含量显著高于15—30 cm土层。在种植油松苗木组,0—15 cm土层,ETP、ECP和ETCP均显著增加了土壤DON含量;15—30 cm土层,ECP和ETCP也显著增加了土壤DON含量。在未种树组,0—15 cm土层与15—30 cm土层相似,ETS显著增加了土壤DON含量。
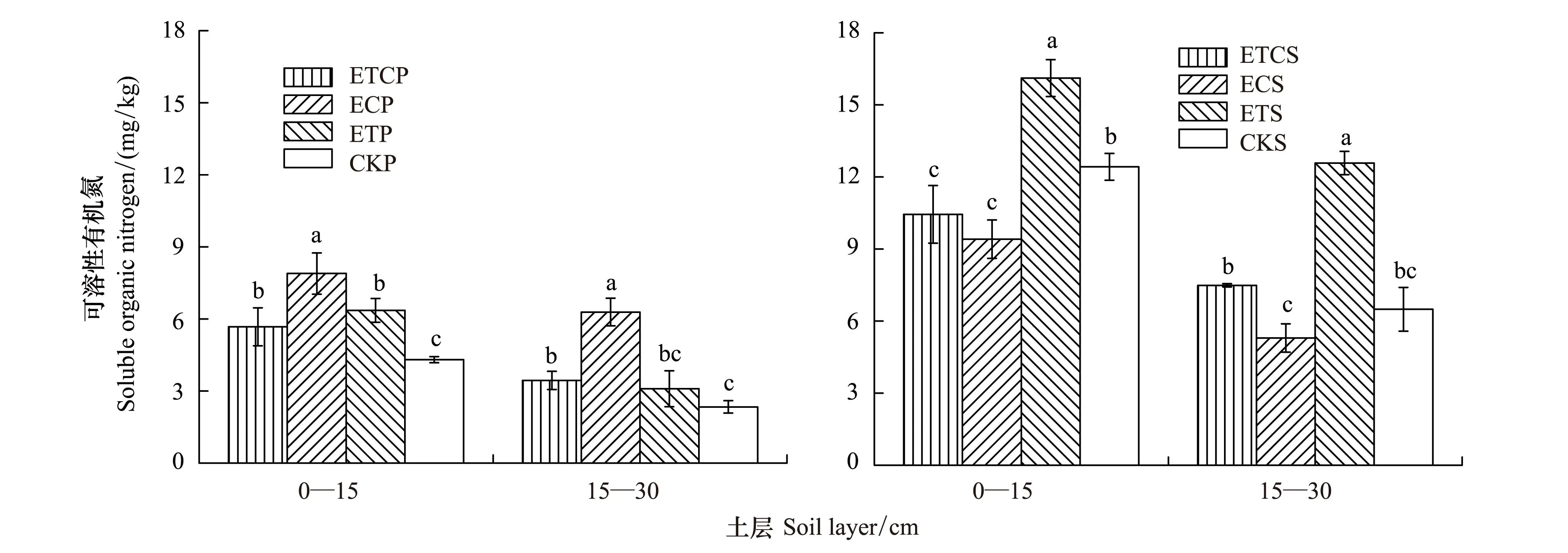
图4 增温、增CO2及其共同作用对不同土层土壤可溶性有机氮含量的影响[18]Fig.4 Effects of ET、EC and ETC on the content of soil soluble organic nitrogen (DON) at different soil layers.直方柱上不同小写字母表示同一土层不同处理间差异显著 (P < 0. 05)

图5 增温、增CO2及其共同作用对不同土层土壤可溶性总氮含量的影响Fig.5 Effects of ET、EC、ETC on the content of soil total soluble nitrogen (TSN) at different soil layers.直方柱上不同小写字母表示同一土层不同处理间差异显著 (P < 0. 05)
2.5增温与增CO2对可溶性总氮含量的影响
由图5可知,与未种树相比较,种植油松苗木显著降低了土壤TSN含量,且0—15 cm层土壤TSN含量显著高于15—30 cm土层。在种植油松苗木组,0—15 cm土层,与CKP相比,ETP、ECP和ETCP均显著增加了土壤TSN含量,ETCP、ETP、ECP三者间差异不显著;15—30 cm土层,ETCP、ECP和ETP均增加了土壤TSN含量,且ECP和ETP达到显著水平。在未种树组,0—15 cm土层,ETS显著增加了土壤TSN含量,增加幅度达48.6%,而ETCS、ECS与CKS差异不显著;15—30 cm土层,ETCS、ETS显著增加了土壤TSN含量,而ECS显著降低了土壤TSN含量。
3讨论
3.1增温与增CO2对可溶性氮的影响
3.1.1增温与增CO2对硝态氮的影响

3.1.2增温与增CO2对土壤铵态氮含量的影响
3.1.3增温与增CO2对土壤游离氨基酸的影响
在种植油松苗木组和未种树组,增温处理均显著增加了土壤游离氨基酸含量,而EC、ETC与CK的差异不显著。土壤游离氨基酸主要来源于根系、土壤微生物分泌物和土壤中各种有机物质的降解产物,虽然游离氨基酸占土壤 TSN的比例较小,但其在土壤中具有很高的周转速率。增温处理使土壤微生物活性及代谢作用增强,促进了土壤氮矿化作用[20],同时增温有利于促进种树组植物根系分泌物的增加[29],根系和土壤微生物分泌物是土壤游离氨基酸的重要来源,因此增温有利于土壤游离氨基酸的含量增加。
3.1.4增温与增CO2对土壤可溶性有机氮的影响
种植油松苗木组,ETCP、ETP和ECP均显著增加了土壤DON含量。ETCP、ETP和ECP处理加速了植物生长,有利于根系分泌物的增加[29-30],加速土壤DON含量的生成。 Bengtson和Barker[31]进行的增温实验也得出了相似结论,增温有利于土壤有机质快速降解,使可溶性有机质含量增加。
未种树组,增温显著增加了土壤DON含量,而ECS却显著降低了土壤表层DON含量。增温处理有利于增加微生物数量[28],有利于对土壤有机质的降解,且微生物代谢产物是可溶性有机物的重要组成部分,因此增温增加了土壤DON含量。而增CO2处理使土壤空隙的CO2浓度增加[22],在一定程度上改变了土壤中微生物的生长环境和群落结构[32],进而引起土壤DON含量的变化。
DON是林地土壤中可溶性氮的主要组分[33],本研究得出土壤DON含量变化于2.3—16.1 mg/kg之间,平均值约占土壤可溶性总氮的38.9%,这与大量研究结果一致,Chen等[34]综合统计了116份已发表数据,表明林地土壤生态系统DON含量变化于1—448 mg/kg,占土壤可溶性总氮比例的48%。
3.1.5增温与增CO2对土壤可溶性总氮的影响
在种植油松苗木组,ETCP、ETP和ECP均显著增加了土壤可溶性总氮含量,可溶性有机氮和铵态氮含量的增加是其增加的主要原因。温度的升高和 CO2浓度的上升有利于增加植物细根的生长、根系周转速率、根际微生物数量和根系分泌物数量等[29,35- 36],因此促使土壤可溶性总氮含量增加。
在未种树组,增温显著增加了土壤可溶性总氮含量,这主要源于增温促使了可溶性无机氮和可溶性有机氮的整体增加。由于增温处理加快了土壤有机质的降解和微生物的转化速率,因此增温处理使土壤可溶性总氮含量增加。
3.2植物对不同形态氮的吸收
种植油松苗木组土壤硝态氮、铵态氮、游离氨基酸、可溶性有机氮和可溶性总氮含量均显著低于未种树组,这可能是由于土壤有效氮与植物生长密切相关,油松苗在生长过程中对土壤养分的消耗使土壤有效氮含量显著降低。Carrillo等[37]在草原上进行的研究也表明,是否栽种植物显著地影响着可溶性有机氮和无机氮含量,未栽植物的土壤中硝态氮含量是栽种植物土壤的6倍。

4结论

参考文献(References):
[1]薛晓萍, 王建国, 郭文琦, 陈兵林, 尤军, 周治国. 氮素水平对初花后棉株生物量、氮素累积特征及氮素利用率动态变化的影响. 生态学报, 2006, 26(11): 3631- 3640.
[2]莫良玉, 吴良欢, 陶勤南. 高等植物对有机氮吸收与利用研究进展. 生态学报, 2002, 22(1): 118- 124.
[3]Chapin F S, Moilanen L, Kielland K. Preferential use of organic nitrogen for growth by a non-mycorrhizal arctic sedge. Nature, 1993, 361(6408): 150- 153.
[4]Weigelt A, Bol R, Bardgett R D. Preferential uptake of soil nitrogen forms by grassland plant species. Oecologia, 2005, 142(4): 627- 635.
[5]Jones D L, Healey J R, Willett V B, Farrar J F, Hodge A. Dissolved organic nitrogen uptake by plants—an important N uptake pathway?. Soil Biology and Biochemistry, 2005, 37(3): 413- 423.
[6]Jones D L, Hodge A. Biodegradation kinetics and sorption reactions of three differently charged amino acids in soil and their effects on plant organic nitrogen availability. Soil Biology and Biochemistry, 1999, 31(9): 1331- 1342.
[7]Jones D L, Owen A G, Farrar J F. Simple method to enable the high resolution determination of total free amino acids in soil solutions and soil extracts. Soil Biology and Biochemistry, 2002, 34(12): 1893- 1902.
[8]Kuzyakov Y, Xu X L. Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance. New Phytologist, 2013, 198(3): 656- 669.
[9]IPCC. Climate Change 2007: the physical science basis // Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt K B, Tignor M, Miller H L, eds. Contribution of Working Group I to The Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 2007.
[10]尹华军, 赖挺, 程新颖, 蒋先敏, 刘庆. 增温对川西亚高山针叶林内不同光环境下红桦和岷江冷杉幼苗生长和生理的影响. 植物生态学报, 2008, 32(5): 1072- 1083.
[11]Xu Z F, Pu X Z, Yin H J, Zhao C Z, Liu Q, Wu F. Warming effects on the early decomposition of three litter types, Eastern Tibetan Plateau, China. European Journal of Soil Science, 2011, 63(3): 360- 367.
[12]Xu Z F, Yin H J, Xiong P, Wan C, Liu Q. Short-term responses ofPiceaasperataseedlings of different ages grown in two contrasting forest ecosystems to experimental warming. Environmental and Experimental Botany, 2012, 77: 1- 11.
[13]Yin H J, Liu Q, Lai T. Warming effects on growth and physiology in the seedlings of the two conifersPiceaasperataandAbiesfaxonianaunder two contrasting light conditions. Ecological Research, 2008, 23(2): 459- 469.
[14]Xu Z F, Wan C, Xiong P, Tang Z, Hu R, Cao G, Liu Q. Initial responses of soil CO2efflux and C, N pools to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China. Plant and Soil, 2010, 336(1/2): 183- 195.
[15]Xu G, Jiang H, Zhang Y B, Korpelainen H, Li C Y. Effect of warming on extracted soil carbon pools ofAbiesfaxonianaforest at two elevations. Forest Ecology and Management, 2013, 310: 357- 365.
[16]Zhao C Z, Liu Q. Growth and photosynthetic responses of two coniferous species to experimental warming and nitrogen fertilization. Canadian Journal of Forest Research, 2009, 39(1): 1- 11.
[17]江元明, 庞学勇, 包维楷. 岷江上游油松与云杉人工林土壤微生物生物量及其影响因素. 生态学报, 2011, 31(3): 801- 811.
[18]刘芙蓉, 王红梅, 张咏梅. 增温和CO2浓度加倍对川西亚高山针叶林土壤可溶性有机碳、氮的影响. 生态学杂志, 2013, 32(11): 2844- 2849.
[19]Zhang Y B, Duan B L, Qiao Y Z, Wang K Y, Korpelainen H, Li C Y. Leaf photosynthesis ofBetulaalbosinensisseedlings as affected by elevated CO2and planting density. Forest Ecology and Management, 2008, 255(5/6): 1937- 1944.
[20]Yin H J, Chen Z, Liu Q. Effects of experimental warming on soil N transformations of two coniferous species, Eastern Tibetan Plateau, China. Soil Biology and Biochemistry, 2012, 50: 77- 84.
[21]Rustad L, Campbell J, Marion G, Norby R, Mitchell M, Hartley A, Cornelissen J, Gurevitch J. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia, 2001, 126(4): 543- 562.
[22]Jackson R B, Cook C W, Pippen J S, Palmer S M. Increased belowground biomass and soil CO2fluxes after a decade of carbon dioxide enrichment in a warm-temperate forest. Ecology, 2009, 90(12): 3352- 3366.
[23]贺纪正, 张丽梅. 土壤氮素转化的关键微生物过程及机制. 微生物学通报, 2013, 40(1): 98- 108.
[24]俞慎, 李振高. 稻田生态系统生物硝化-反硝化作用与氮素损失. 应用生态学报, 1999, 10(5): 630- 634.
[25]Luo Y Q, Gerten D, Le Maire G, Parton W J, Weng E, Zhou X H, Keough C, Beier C, Ciais P, Cramer W, Dukes J S, Emmett B, Hanson P J, Knapp A, Linder S, Nepstad DAN, Rustad L. Modeled interactive effects of precipitation, temperature, and [CO2] on ecosystem carbon and water dynamics in different climatic zones. Global Change Biology, 2008, 14(9): 1986- 1999.
[26]Dieleman W I, Vicca S, Dijkstra F A, Hagedorn F, Hovenden M J, Larsen K S, Morgan J A, Volder A, Beier C, Dukes J S. Simple additive effects are rare: a quantitative review of plant biomass and soil process responses to combined manipulations of CO2and temperature. Global Change Biology, 2012, 18(9): 2681- 2693.
[27]Wang L, Pedas P, Eriksson D, Schjoerring J K. Elevated atmospheric CO2decreases the ammonia compensation point of barley plants. Journal of Experimental Botany, 2013, 64(10): 2713- 2724.
[28]Ziegler S E, Billings S A, Lane C S, Li J W, Fogel M L. Warming alters routing of labile and slower-turnover carbon through distinct microbial groups in boreal forest organic soils. Soil Biology & Biochemistry, 2013, 60: 23- 32.
[29]Yin H J, Li Y F, Xiao J, Xu Z F, Cheng X Y, Liu Q. Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming. Global Change Biology, 2013, 19(7): 2158- 2167.
[30]Phillips R P, Bernhardt E S, Schlesinger W H. Elevated CO2increases root exudation from loblolly pine (Pinustaeda) seedlings as an N-mediated response. Tree Physiology, 2009, 29(12): 1513- 1523.
[31]Bengtson P, Bengtsson G. Rapid turnover of DOC in temperate forests accounts for increased CO2production at elevated temperatures. Ecology Letters, 2007, 10(9): 783- 790.
[32]Billings S, Ziegler S. Linking microbial activity and soil organic matter transformations in forest soils under elevated CO2. Global Change Biology, 2005, 11(2): 203- 212.
[33]Xing S H, Chen C R, Zhou B Q, Zhang H, Nang Z M, Xu Z H. Soil soluble organic nitrogen and active microbial characteristics under adjacent coniferous and broadleaf plantation forests. Journal of Soils and Sediments, 2010, 10(4): 748- 757.
[34]Chen C R, Xu Z H. Analysis and behavior of soluble organic nitrogen in forest soils. Journal of Soils and Sediments, 2008, 8(6): 363- 378.
[35]Phillips D A, Fox TC, Six J. Root exudation (net efflux of amino acids) may increase rhizodeposition under elevated CO2. Global Change Biology, 2006, 12(3): 561- 567.
[36]Phillips R P, Finzi AC, Bernhardt E S. Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2fumigation. Ecology Letters, 2011, 14(2): 187- 194.
[37]Carrillo Y, Dijkstra F A, Pendall E, Morgan J A, Blumenthal D M. Controls over soil nitrogen pools in a semiarid grassland under elevated CO2and warming. Ecosystems, 2012, 15(5): 761- 774.
[38]何文寿, 李生秀, 李辉桃. 六种作物不同生育期吸收铵、硝态氮的特性. 作物学报, 1999, 25(2): 222- 226.
[39]周晓红, 王国祥, 杨飞, 何伟, 杨周. 空心菜对不同形态氮吸收动力学特性研究. 水土保持研究, 2008, 15(5): 84- 87.
[40]Owen A G, Jones D L. Competition for amino acids between wheat roots and rhizosphere microorganisms and the role of amino acids in plantNacquisition. Soil Biology &Biochemistry, 2001, 33(4/5): 651- 657.
Effects of elevated temperature and CO2concentration doubling on soil total
soluble nitrogen in subalpine coniferous forest of western Sichuan, China
LIU Furong1,2, ZHANG Yongmei1,*, DENG Shulin1,2
1ChengduInstituteofBiology,ChineseAcademyofSciences,Chengdu610041,China2UniversityofChineseAcademyofSciences,Beijing100049,China
Abstract:The Qinghai-Tibetan Plateau, often referred to as “the Third Pole” of the world, plays an important role in the Earth′s climate system. Chinese pine (Pinus tabulaeformis) forest is one of the most important vegetation types in the subalpine regions of western Sichuan, China, but our knowledge about the response of soil in this forest ecosystem, especially soil total soluble nitrogen to climate change is limited. The effects of elevated temperate (ET, ambient temperature + (2.5±0.5)℃), CO2concentration doubling (EC, ambient CO2 concentration + 350 μmol/mol) and their interaction (ETC) on soil total soluble nitrogen, including nitrate nitrogen -N), ammonium nitrogen -N), free amino acid (FAA), dissolved organic nitrogen (DON) and total soluble nitrogen (TSN) of Chinese pine forest soils were investigated by using an automatic micro-climate controlled system.1) Compared with the control (CKP), ET significantly decreased -N concentrations in the seedling treatment, and the concentration of -N in the 0—15 cm soil layer was lower than that in the 15—30 cm layer. By contrast, ET markedly increased -N concentrations in the plant-free treatment, and the concentration of -N in the 0—15 cm soil layer was higher than that in the 15—30 cm layer. These results indicated that the absorption of -N by Chinese pine seedlings of was enhanced under ET to meet the demands of growth, especially in the 0—15 cm soil layer. This was likely due to the occurrence of more fine roots in the upper soil layer than the deeper layer. -N appears to be one of the most important forms of soil soluble nitrogen utilized by Chinese pine. 2) Furthermore, ET, EC and ETC induced an increase in the concentrations of -N, DON and TSN in the seedling treatment. However, in the plant-free treatment -N, FAA, DON and TSN concentrations were significantly enhanced under ET; but EC and ETC had little influence on their concentrations. These results suggest that EC and ETC increased -N, DON and TSN concentrations mainly through the plant roots, but ET acted by influencing both soil microorganisms and plant root systems. 3) -N, -N, FAA, DON and TSN concentrations in the seedling treatment were significantly lower than those in the plant-free group, which might be attributable to the absorption of soil soluble nitrogen by plants to meet growth demands. Overall, in the plant-free group, the signicant increases in -N, -N, FAA, DON and TSN under ET compared with the control (CKS) indicate that warming contributed to the enhanced efficiency of soil microbes. However, in the seedling treatment, -N, -N, DON and TSN concentrations were influenced by both soil microorganisms and plants. Moreover, the amount of soil soluble nitrogen absorbed by Pinus tabulaeformis mainly varied among the different forms of nitrogen.
Key Words:elevated temperature; CO2 concentration doubling; nitrate nitrogen -N); ammonia nitrogen -N); free amino acid (FAA); dissolved organic nitrogen (DON); total soluble nitrogen (TSN)
DOI:10.5846/stxb201405080906
*通讯作者
Corresponding author.E-mail: zhangym@cib.ac.cn
收稿日期:2014- 05- 08; 网络出版日期:2015- 06- 12
基金项目:国家自然科学基金项目(30972345,31100383)
刘芙蓉, 张咏梅, 邓书林.增温和CO2浓度加倍对川西亚高山针叶林土壤可溶性氮的影响.生态学报,2016,36(3):652- 660.
Liu F R, Zhang Y M, Deng S L.Effects of elevated temperature and CO2concentration doubling on soil total soluble nitrogen in subalpine coniferous forest of western Sichuan, China.Acta Ecologica Sinica,2016,36(3):652- 660.
