葡萄砧木枝条的抗寒性比较
2016-03-17钟海霞潘明启张付春艾尔买克才卡斯木韩守安
钟海霞,潘明启,张付春,艾尔买克·才卡斯木,韩守安,
张 雯,谢 辉,陈 锐,梅 闯,伍新宇
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测试验站,乌鲁木齐 830091)
葡萄砧木枝条的抗寒性比较
钟海霞,潘明启,张付春,艾尔买克·才卡斯木,韩守安,
张 雯,谢 辉,陈 锐,梅 闯,伍新宇
(新疆农业科学院园艺作物研究所/农业部新疆地区果树科学观测试验站,乌鲁木齐830091)
摘要:【目的】探讨葡萄砧木枝条的抗寒性,为新疆葡萄砧木的选择、引进及抗寒栽培提供理论依据。【方法】以8 种葡萄砧木为研究对象,对低温胁迫下各砧木品种枝条的相对电导率、游离脯氨酸、丙二醛、可溶性糖、可溶性蛋白、超氧化物歧化酶、过氧化物酶和过氧化氢酶的活性进行测定。应用隶属函数法对各项生理生化指标进行抗寒综合评价。【结果】随温度的降低,所有砧木品种枝条的相对电导率、脯氨酸、丙二醛和可溶性糖含量均逐渐增加,但增加幅度有所不同;可溶性蛋白、SOD和POD的含量均呈现先升高后降低的趋势; CAT含量变化不规律。根据隶属函数法综合判断得出葡萄砧木抗寒性强弱顺序为:脱毒贝达>5C>110R>101-14>188-08>3 309C>5BB>SO4。根据恢复生长验证得出各品种抗寒性强弱与各抗寒理化指标进行隶属函数法得出的砧木抗寒性排序结果基本一致。【结论】8种砧木中脱毒贝达抗寒性最强,能抵御的最低温度达-30℃以下;5C、110R、101-14、188-08、3 309C和5BB抗寒性较强,能抵御的最低温度为-20~-30℃;SO4抗寒性最弱,能抵御的最低温度为-20℃。
关键词:低温胁迫;葡萄砧木;抗寒性
0引 言
【研究意义】新疆是国内最大的葡萄产区,栽培面积超过了全国的20%。北疆地区冬季严寒,易发生低温冻害,影响了果农的经济效益和葡萄产业的发展。利用葡萄抗寒砧木进行嫁接栽培,一方面能减轻或避免葡萄冻害,另一方面可改善树体营养,具有广阔的发展前景[1]。研究葡萄砧木的抗寒性,筛选出适宜新疆发展的抗寒砧木品种,对新疆葡萄抗寒砧木的引进、推广和新品种的选育具有重要意义。【前人研究进展】前人对葡萄抗寒性鉴定方面已有一些研究。牛锦凤等[2]研究发现7 个鲜食葡萄品种中矢富罗莎、奥古斯特和京秀抗寒性较强, 红地球和美人指居中, 而乍娜和里扎马特抗寒性最弱;王燕凌等[3]对3个鲜食葡萄抗寒研究表明随温度的降低,游离脯氨酸和可溶性糖的含量均逐渐升高,SOD和POD活性呈显著上升的趋势;鲁金星等[4]研究认为贝达抗寒性最强,双优+贝达、双红+贝达、维代尔和赤霞珠抗寒性居中,梅鹿辄抗寒性最弱。隶属函数法能对各项抗寒指标综合评价能分析出品种间的抗寒性差异,在苹果和扁桃等果树上已得到应用[5-7]。【本研究切入点】新疆葡萄需要选择和利用抗寒砧木进行抗寒栽培,而目前有关葡萄砧木的抗寒性研究报道较少。试验对8 种葡萄砧木品种的一年生枝条的抗寒性理化指标进行测定,比较分析和综合评价各品种抗寒性,结合可恢复生长试验进行各品种抗寒能力验证。【拟解决的关键问题】对8个葡萄砧木品种的抗寒能力强弱进行排序比较,找出各砧木能抵御的最低温度,为新疆产区葡萄抗寒砧木选择、引进与抗寒栽培工作提供理论依据。
1材料与方法
1.1 材 料
以3年生8种葡萄砧木101-14、110 R、188-08、3309C、5C、5BB、SO4和脱毒贝达(均从河北引进)的1年生枝条为试材,2013年11月5日埋土前,于新疆农科院安宁渠综合试验场园艺所葡萄基地核心示范园采集,各品种分别选取15株,每株剪取3个1年生枝条,枝条直径0.5~0.8 mm。
1.2 方 法
将1 年生枝条用蒸馏水冲洗干净,每2~3节剪成一段,每个品种6份,每份30段,用塑料布包好,放入高低温交变试验箱中进行处理,试验设-2(对照)、-15、-20、-25、-30、-35℃ 6个温度梯度处理,以 4℃/h 降温速度,降温至不同处理温度后持续12 h,之后逐步升温至0℃,取出后在0℃下放置8 h,室温下再放置8 h,自封袋密封后置于-2℃冰箱待测。各指标测定前,将枝条节间切成 3~5 mm的薄片混匀后进行各指标测定,3次重复。
用DDS-12A型电导仪测定电导率,计算相对电导率;用茚三酮比色法测定脯氨酸含量[8];用蒽酮显色法测定可溶性糖含量;用G-250考马斯亮蓝法测定可溶性蛋白含量;用硫代巴比妥酸法测定丙二醛含量;用氮蓝四唑 (NBT)法测定超氧化物歧化酶(SOD)活性;用愈创木酚法测定过氧化物酶(POD)活性;用紫外分光光度计法测定过氧化氢酶(CAT)活性[9]。
隶属函数法[7]对各生理指标进行抗寒性综合评价,与抗寒性呈正相关的指标计算:Tij=(Xij-Xjmin)/(Xjmax-Xjmin);与抗寒性呈负相关的指标隶属函数值计算:Tij=1-(Xij-Xjmin)/(Xjmax-Xjmin)。其中Tij表示i葡萄砧木品种j生理生化指标的隶属函数值;Xij表示i葡萄砧木品种j生理生化指标的测定值;Xjmax和Xjmin分别为8种葡萄砧木品种6个温度梯度中j生理生化指标的最大值和最小值。对各项理化指标隶属函数值的算术平均值进行比较,综合评价排名次序即各葡萄砧木品种抗寒的强弱能力。数据处理及分析用Excel 2007和Spss 17.0软件。
可恢复性生长试验:2013年11月10日,将选取的1年生枝条用清水洗净、酒精(75%)消毒后,扦插在20×20(cm)的花盆(蛭石与珍珠岩=1∶1)中,各品种扦插3盆,定期浇水或除湿,用塑料膜覆盖做成拱形小棚,棚内温度维持在20~25℃,每 3 d观察一次,待枝条发芽、展叶、开花后,记录其萌芽率。
2结果与分析
2.1 低温胁迫对枝条相对电导率和游离脯氨酸的影响
果树遭受低温胁迫时,因质膜的透性发生变化或丧失功能,胞内电解质开始大量外渗,导致组织浸泡液的电导率增大。通过测定外渗液的电导度增加值能反映出质膜的受伤害程度。研究表明,随处理温度的降低,8个葡萄砧木品种的相对电导率均显著升高,对细胞膜的伤害也明显增大,说明相对电导率与葡萄砧木的抗寒性关系密切。各品种相对电导率在-2℃时最低,-35℃时均达到最高。-15℃时,8个砧木品种的相对电导率差别不大,-20℃时SO4枝条的相对电导率变化幅度最大,从45.53%上升至66.02%,增加了20.49%。188-08、3309C、5C和5BB枝条的相对电导率变化幅度居中;脱毒贝达、110 R 、101-14枝条的相对电导率变化幅度较小,分别上升了4.36%、2.58%、1.65%。说明-20℃的低温对砧木枝条组织细胞膜的损伤较小,脱毒贝达、110R 、101-14 的枝条对低温的忍耐力较强。188-08、3309C、5C和5BB在-20~-25℃时枝条的相对电导率有明显的上升,-25℃之后变得平缓。温度在-25℃时,8个品种的相对电导率均在50%以上。脱毒贝达、110R和101-14在-30~-35℃的相对电导率变化幅度显著上升,分别增加了27.76%、17.59%、11.30%,其余品种呈平缓升高态势,此时大部分组织已受冻害损伤,细胞甚至死亡,细胞膜透性功能丧失。图1
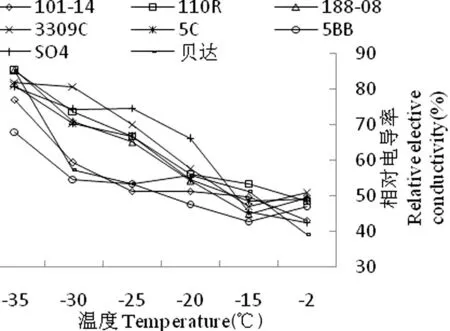
图1不同低温处理下葡萄砧木枝条相对电导率变化
Fig.1The effect of different low temperature treatment on relative elective conductivity of branches of grape rootstocks
果树在正常条件下,游离脯氨酸含量很低,但遇到低温逆境时,游离脯氨酸便会大量积累。脯氨酸是植物抗寒性的一项生化指标。研究表明,葡萄砧木品种游离脯氨酸含量随温度的降低而呈持续上升的态势。脱毒贝达、SO4和3309C在-25~-35℃枝条游离脯氨酸迅速增加且积累量大,3个品种增加幅度分别为43.77,69.24和46.48 μ/g。101-14、110R、188-08、5 C、5BB枝条游离脯氨酸在-2~-20℃迅速增加,增加幅度最大的为188-08,游离脯氨酸含量增加了50.77 μ/g。但这5个品种的游离脯氨酸含量在-25℃之后变化不大。总体而言脱毒贝达、SO4和3309C这 3个品种脯氨酸含量较高,积累量较大,对低温胁迫做出了积极响应。图2

图2不同低温处理下葡萄砧木枝条游离脯氨酸变化
Fig.2The effect of different low temperature treatment on freeproline content of branches of grape rootstocks
2.2 低温胁迫对丙二醛(MDA)和可溶性糖影响
植物遭受逆境胁迫后常发生膜脂过氧化作用,其最终产物是丙二醛(MDA)。MDA可与蛋白质和核酸反应,改变其构型或使之发生交联反应,从而丧失功能。MDA的积累是鉴别逆境伤害的指标之一。研究表明,8种葡萄品种的MDA含量均随温度的降低呈现逐渐增加的趋势。在-2~-35℃,脱毒贝达枝条MDA的含量一直处于较低水平,变化幅度不大;5C和3309C枝条的 MDA 含量居中;188-08枝条的 MDA含量大多处于较高水平,显著大于其余品种,在-20℃时188-08的枝条MDA 含量比脱毒贝达高出了44%。综合比较得出8个品种中188-08枝条细胞膜的损害度最高,膜透性最大。图3

图3不同低温处理下葡萄砧木枝条MDA含量变化
Fig.3The effect of different low temperature treatment on MDA content of branches of grape rootstocks
作为细胞渗透调节物质的可溶性糖在低温逆境中会大量积累,它与植物抗寒性密切相关。即可溶性糖含量越高,则植物抗寒性越强。葡萄砧木枝条内可溶性糖含量的变化因品种不同存在差异。随着处理温度的下降,砧木枝条中可溶性糖的含量逐渐增大。-25℃时8 个品种枝条中可溶性糖含量无显著性差异(P>0.05)。-25~-30℃时,各品种枝条可溶性糖含量迅速升高,脱毒贝达枝条的可溶性糖含量增加幅度最大,达0.28%。脱毒贝达、SO4、5BB、5C、188-08和3 309 C 这6个品种枝条可溶性糖含量均在-35℃时达到最大值,品种间呈显著性差异,其中增加幅度最大的为脱毒贝达。101-14和110R砧木枝条内可溶性糖含量在-30℃时达到峰值,之后逐渐下降,且品种间无显著性差异。图4

图4不同低温处理下葡萄砧木枝条可溶性糖变化
Fig.4 The effect of different low temperature treatment on soluble sugar content of branches of grape rootstocks
2.3 低温胁迫对可溶性蛋白含量和SOD的影响
低温胁迫后,葡萄砧木枝条可溶性蛋白含量会有所增加,对砧木抗寒起到调节作用。研究表明,随着处理温度的降低,不同品种的葡萄砧木枝条的可溶性蛋白质含量变化趋势有所不同。8个砧木品种在-2~-15℃的可溶性蛋白质含量迅速升高,之后上升较缓慢,达到峰值后呈缓慢下降趋势。各品种枝条可溶性蛋白质含量在-15℃时均低于其余温度的值,且品种间无显著差异。总体比较,低温胁迫下脱毒贝达、3309C、5BB的枝条可溶性蛋白含量均处于较高水平。图5

图5不同低温处理下葡萄砧木枝条可溶性蛋白变化
Fig.5 The effect of different low temperature treatment on soluble protein content of branches of grape rootstocks
超氧化物歧化酶(SOD)的活力与植物抗逆性有密切的关系。研究表明,随温度的逐渐降低,葡萄品种SOD 含量变化均呈现先升高后下降的趋势,但各品种SOD活性到达峰值的温度有所不同。SO4枝条的SOD活性在-25℃时达到峰值;101R、3309C、5C、5BB枝条的SOD活性均在-30℃左右时达到最大值,而脱毒贝达枝条SOD活性在-35℃时达到峰值。在-2~-30℃时各品种之间SOD 活性呈显著性差异。在-30~-35℃时,脱毒贝达枝条SOD 的活性呈现降低趋势,显著低于其他几个品种,说明脱毒贝达的酶系统遭受了严重破坏。图6
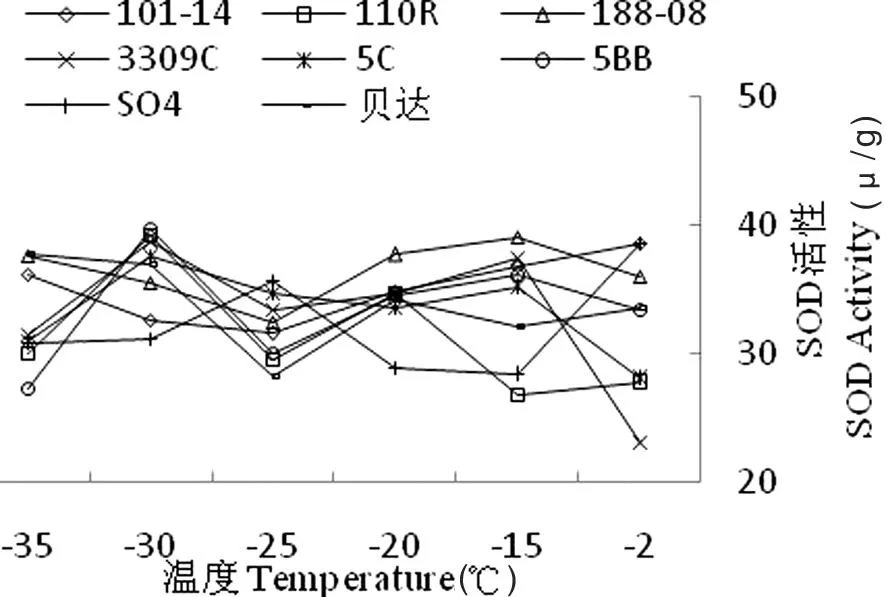
图6不同低温处理下葡萄砧木枝条SOD含量变化
Fig.6 The effect of different low temperature treatment on SOD content of branches of grape rootstocks
2.4 低温胁迫对POD和CAT活性的影响
8 个砧木品种的 POD活性随温度的逐渐降低明显呈先升后降趋势,均在-25℃时POD活性达到最大值,其中贝达的值最大,为17.62 μ/g;在 -2~-25℃低温下,101-14、110R、188和SO4枝条的POD活性显著低于其他品种;在-25~-30℃温度条件下,脱毒贝达、SO4、188、101-14枝条的POD 活性的降幅比-2℃大。推测-25℃左右是8种葡萄砧木的耐受温度,低于-25℃时,可采取一系列酶对葡萄砧木枝条细胞膜的保护措施。图7
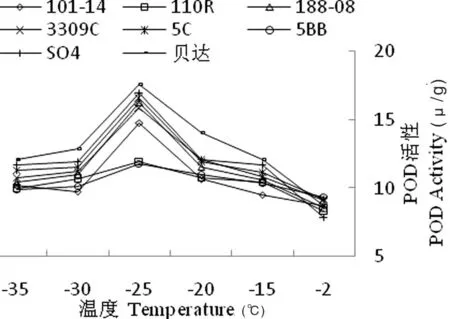
图7不同低温处理下葡萄砧木枝条POD含量变化
Fig.7 The effect of different low temperature treatment on POD content of branches of grape rootstocks
各砧木品种CAT活性随温度的降低各不相同,均呈现不规则变化。脱毒贝达、SO4、101-14、110R枝条品种的CAT 活性均呈现先缓慢下降后增加,随后又下降的趋势; 3309C和5C枝条的CAT 活性含量呈先升后降的趋势,而188-08和5BB呈先降后升的趋势。脱毒贝达、SO4、5BB、5C、110R枝条CAT 活性均在-35℃时达到峰值,其中110R枝条CAT 活性最高,为34.68 u/g。101-14、188-08、3309C在-30℃时CAT活性已经达到了峰值,其中101-14枝条CAT 活性最高,为28.85 μ/g,这3个品种的CAT活性在 -30~-35℃均呈下降趋势。 图8

图8不同低温处理下葡萄砧木枝条CAT含量变化
Fig.8 The effect of different low temperature treatment on CAT content of branches of grape rootstocks
2.5 8种葡萄砧木抗寒性的综合评价
用隶属函数法求得各项指标的平均隶属度,其平均隶属度越大,抗寒性越强,反之抗寒性越弱。脱毒贝达各指标综合的隶属函数值最大,为0.565,其次是5C、110R、101-14、188-08、3309C和5BB,隶属函数值最小的品种是SO4,为0.448。总体而言,8种葡萄砧木品种的抗寒性强弱依次为:脱毒贝达>5C>110R>101-14>188-08> 3309C>5BB>SO4。表1
表1低温胁迫下8种葡萄砧木抗寒指标的隶属度值
Table 1 Subordination value of cold resistance indicator of eight grape rootstock under cold stress
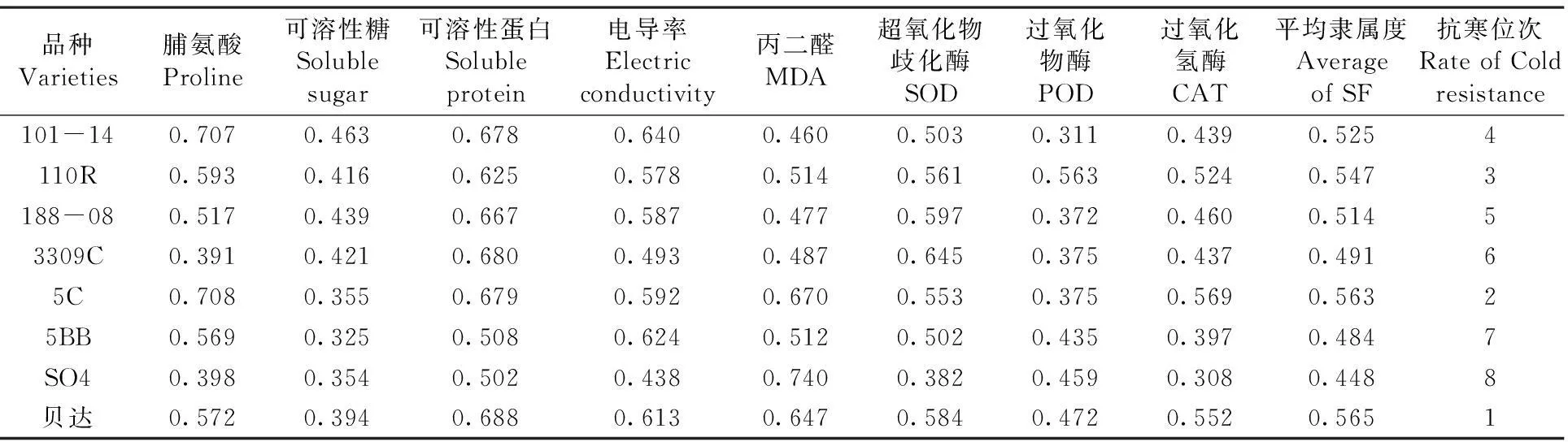
品种Varieties脯氨酸Proline可溶性糖Solublesugar可溶性蛋白Solubleprotein电导率Electricconductivity丙二醛MDA超氧化物歧化酶SOD过氧化物酶POD过氧化氢酶CAT平均隶属度AverageofSF抗寒位次RateofColdresistance101-140.7070.4630.6780.6400.4600.5030.3110.4390.5254110R0.5930.4160.6250.5780.5140.5610.5630.5240.5473188-080.5170.4390.6670.5870.4770.5970.3720.4600.51453309C0.3910.4210.6800.4930.4870.6450.3750.4370.49165C0.7080.3550.6790.5920.6700.5530.3750.5690.56325BB0.5690.3250.5080.6240.5120.5020.4350.3970.4847SO40.3980.3540.5020.4380.7400.3820.4590.3080.4488贝达0.5720.3940.6880.6130.6470.5840.4720.5520.5651
注: 表中抗寒位次数字小表示抗寒性强
Note: The smaller number of rate of cold resistance represents the stronger cold resistance
2.6 可恢复生长验证
通过对低温胁迫后8个葡萄砧木品种的休眠枝条进行可恢复生长试验,得出各品种枝条在-2~-15℃低温胁迫后的萌芽率为60%~100%,均处于较高水平。在-20℃时,各品种萌芽率差异显著,其中脱毒贝达萌芽率最高,达68.2%,比萌芽率最低的SO4高出了62.9%;-25℃时,除脱毒贝达、110R和101-14有部分萌芽外,其余品种均无萌芽,说明5C、188-08、3309C、5BB和SO4的抗寒性较弱;-30℃时,脱毒贝达和110R仍有萌芽现象,其中脱毒贝达的萌芽率达12.8%,比110R的萌芽率高出7.2%,表明110R的抗寒性弱于脱毒贝达;-35℃时,8个砧木品种均无萌芽现象,说明此时砧木受低温冻害严重。总体看来,脱毒贝达砧木抗寒性最强, 能抵御的最低温度达-30℃以下,其次是110R和101-14,能抵御的最低温度分别为-30和-25℃,5C、188-08、3309C、5BB和SO4抗寒性较弱,能抵御的最低温度分别为-20℃。根据恢复生长验证得出各品种抗寒性强弱顺序为:脱毒贝达>110R>101-14>5BB>188-08>5C>3309C>SO4,这与通过各抗寒理化指标综合评价得出的砧木抗寒性排序结果基本一致。表2
表2低温胁迫下8种葡萄砧木枝条恢复生长萌芽率
Table 2 Eight types of grape rootstock under low temperature stress andthe shoot germination rateof recovery(%)
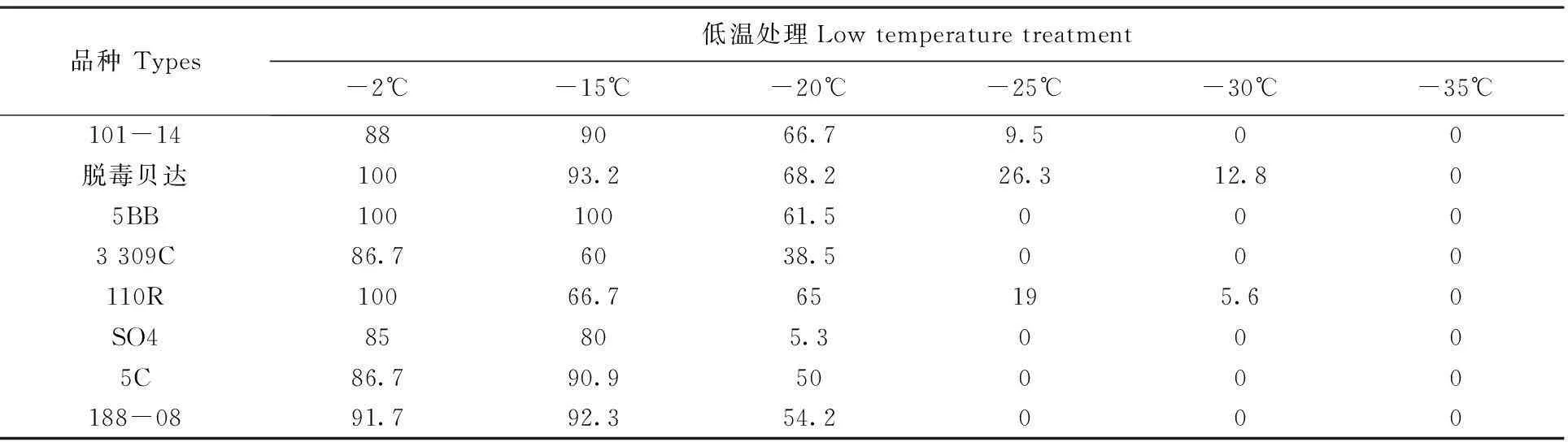
品种Types低温处理Lowtemperaturetreatment-2℃-15℃-20℃-25℃-30℃-35℃101-14889066.79.500脱毒贝达10093.268.226.312.805BB10010061.50003309C86.76038.5000110R10066.765195.60SO485805.30005C86.790.950000188-0891.792.354.2000
3讨 论
低温胁迫下树体的相对电导率增加,细胞代谢失调,严重时导致细胞死亡。试验结果表明随处理温度的降低,8种葡萄砧木枝条的相对电导率呈明显增加趋势,这与张连杰等[10]研究结果一致。果树在低温逆境中,体内会积累大量的游离脯氨酸,但脯氨酸的积累与葡萄抗寒性的关系尚不明确。脯氨酸能保护渗透活性物质,但其调节作用微小[11]。研究得出脯氨酸含量随处理温度的逐渐降低呈现了持续升高的趋势,这与付晓伟等[12]研究结果一致。MDA是膜脂过氧化作用的最终产物,其含量可判别细胞膜系统遭受损害的程度。低温胁迫中MDA含量越高,膜系统损害越严重[2]。研究发现随着温度的降低,MDA的含量呈现不断上升趋势,与谢丽芳等[13]研究结果一致。
可溶性糖是低温胁迫下植物体内产生的保护物质,可提高细胞体内的渗透浓度,降低水势,减缓细胞质过度脱水,提高其抗寒性,抗寒性强的品种可溶性糖含量增多,浓度较高,反之亦然[14]。研究结果表明低温胁迫下,脱毒贝达可溶性糖含量较高,抗寒性较强;5C和188-08可溶性糖含量较低,抗寒性较弱,这与曹建东等[14]研究结果一致。可溶性蛋白具有较强的亲水能力,可减少低温对组织的损害。而试验中8种葡萄砧木的可溶性蛋白含量随温度的降低呈先升后降趋势,增加幅度的大小可反映其抗寒性强弱。总体来看,3309C、5BB等枝条可溶性蛋白含量较高,而抗寒性强的贝达砧木的可溶性蛋白并未高于其余品种,这可能与品种间的差异有关。
SOD 、POD和CAT是低温逆境下树体的保护酶,它们3个酶协同作用能防御活性氧或其他过氧化物自由基对细胞膜系统的伤害。酶促保护系统性能增强可提高组织的抗寒性。试验8 种葡萄砧木枝条 SOD 和POD含量随着胁迫温度的降低而呈现先升高后降低的趋势,对低温胁迫产生了积极的响应,但各品种CAT 含量的变化不规律,这与鲁金星等[4]研究结果不一致,可能与试验材料与环境条件不同有关。
不同品种的抗寒性存在一定的差异,具体比较葡萄砧木抗寒性的强弱,需对各项理化指标进行综合评价。研究将人工模拟外界低温胁迫与实验室理化鉴定相结合,应用隶属函数法对 8种葡萄砧木的抗寒性进行综合评价,得出脱毒贝达的抗寒性最强,5C、110R、101-14、188-08、 3309C和5BB的抗寒性较强, SO4的抗寒性最弱。这与白茹等[15]研究得出贝达的抗寒性最强,SO4和5BB次之,5C抗寒性最弱的结果不尽相同,这可能与试验环境及方法不同等原因有关。恢复生长试验得出脱毒贝达砧木抗寒性最强,能抵御的最低温度达-30℃以下,其次是110R和101-14,能抵御的最低温度分别为-30和-25℃,5C、188-08、3309C、5BB和SO4抗寒性较弱,能抵御的最低温度分别为-20℃。根据恢复生长验证得出各品种抗寒性强弱顺序为:脱毒贝达>110R>101-14>5BB>188-08>5C>3309C>SO4,这与各抗寒理化指标综合评价得出的砧木抗寒性排序结果基本一致。脱毒贝达虽抗寒性最强,但实际生产中,在新疆乃至西北地区易发生黄化问题,而5C、110R、101-14、3 309C、5BB和SO4等抗寒能力虽不及脱毒贝达,但在新疆适应性强,生长良好,不易发生黄化,对新疆抗寒葡萄砧木的栽培和引种具有指导作用。
4结 论
8 种葡萄砧木中,脱毒贝达抗寒性最强;5C、101-14、188-08、110R、3 309C、5BB抗寒性较强;SO4抗寒性最弱。其中脱毒贝达能抵御的最低温度达-30℃以下,其余品种能抵御的最低温度在-20~-30℃。
参考文献(References)
[1]房玉林,孙伟,张振文,等.葡萄砧木的研究和利用概况[J]. 中外葡萄与葡萄酒,2010,(7): 74-80.
FANG Yu-lin, SUN Wei, ZHANG Zhen-wen, et al. (2010). General situation of research and utilization of grape rootstock [J].Sino-OverseasGrapevine&Wine,(7): 74-80. (in Chinese)
[2] 牛锦凤,王振平,李国,等. 几种方法测定鲜食葡萄枝条抗寒性的比较[J]. 果树学报,2006,23(1):31-34.
NIU Jin-feng, WANG Zhen-ping, LI Guo, et al. (2006). Comparison Of Different Methods For Examination Of Grape Hardiness [J].JournalofFruitScience, 23(1):31-34. (in Chinese)
[3] 王燕凌,廖康,刘君,等. 越冬前低温锻炼期间不同品种葡萄枝条中渗透性物质和保护酶活性的变化[J]. 果树学报,2006,23(3):375-378.
WANG Yan-ling, LIAO Kang, LIU Jun, et al. (2006). Changes of contents of osmosis substances and antioxidant enzyme activity in grape vines during cold exercise period before over-wintering [J].JournalofFruitScience, 23(3):375-378. (in Chinese)
[4] 鲁金星,姜寒玉,李唯. 低温胁迫对砧木及酿酒葡萄枝条抗寒性的影响[J]. 果树学报 ,2012,29(6):1 040-1 046.
LU Jin-xing, JIANG Han-yu, LI Wei. (2012). Effects of low temperature stress on the cold resistance of rootstock and branch of wine grapes [J].JournalofFruitScience, 29(6):1,040-1,046. (in Chinese)
[5] 李轶冰 , 杨顺强 , 任广鑫 , 等 . 低温处理下不同禾本科牧草的生理变化及其抗寒性比较[J]. 生态学报 ,2009, 29(3):1 341-1 347.
LI Yi-bing, YANG Shun-qiang, REN Guang-xin, et al. (2009). Changes analysis in physiological properties of several germanous grass species and cold resistance comparison on under cold stress [J].ActaEcologicaSinica, 29(3):1,341 -1,347. (in Chinese)
[6] 高爱农 , 沙守峰 , 刘志 , 等 . 苹果品种抗寒性测定方法的研究[J]. 果树科学 ,2000,17(1):17-21.
GAO Ai-nong, SHA Shou-feng, LIU Zhi, et al. (2000). Study on the hardiness of apple cultivars [J].JournalofFruitScience, 17(1):17- 21. (in Chinese)
[7] 李斌,刘立强,罗淑萍,等. 扁桃花芽的抗寒性测定与综合评价[J]. 经济林研究, 2012,30(3):16-21.
LI Bin, LIU Li-qiang, LUO Shu-ping, et al. (2012). Determination and comprehensive evaluation on cold resistance of flower bud in almond [J].NonwoodForestResearch, 30(3):16-21. (in Chinese)
[8] 许宏,王孝娣,邹英宁,等. 葡萄砧木及酿酒品种抗寒性比较[J]. 中外葡萄与葡萄酒,2003,(6):20-23.
XU Hong, WANG Xiao-di, ZOU Ying-ning, et al. (2003). Study on the cold resistance of grape rootstocks and wine grape cultivars [J].Sino-OverseasGrapevine&Wine, (6):20-23. (in Chinese)
[9] 邹琦. 植物生理学实验指导[M] .北京:中国农业出版社,2000:131-135.
ZOU Qi. (2000).Plantphysiologyexperimentinstruction[M]. Beijing:China Agriculture Press:131-135. (in Chinese)
[10] 张连杰,李鹏程,郭佳鑫,等. 低温处理对4个葡萄砧木品种抗寒生理指标的影响[J]. 中外葡萄与葡萄酒,2014,(1):26-29.
ZHANG Lian-jie, LI Peng-cheng, GUO Jia-xin, et al. (2014). Cryogenic treatment on cold resistance indexes of four grape rootstocks [J].Sino-OverseasGrapevine&Wine,(1): 26-29. (in Chinese)
[11] Sánchez, F. J., Andrés, E. F. D., Tenorio, J. L., & Ayerbe, L. (2004). Growth of epicotyls, turgor maintenance and osmotic adjustment in pea plants (Pisumsativuml.) subjected to water stress.FieldCropsResearch, 86(3):81-90.
[12] 付晓伟,张倩,刘崇怀,等. 评价葡萄根系抗寒性指标的确定[J]. 果树学报, 2014,31(1):52-59.
FU Xiao-wei, ZHANG Qian, LIU Chong-huai, et al. (2014). Index for the evaluation of grape root cold resistance [J].JournalofFruitScience, 31(1):52-59. (in Chinese)
[13] 谢丽芳. 葡萄砧木抗寒性鉴定及抗寒机理的研究[D]. 呼和浩特:内蒙古农业大学硕士论文,2007.
XIE Li-fang. (2007).Studiesoncoldresistanceidentificationandmechanismofgrapevinerootstocks[D]. Master Dissertation. Inner Mongolia University, Hohhot . (in Chinese)
[14] 曹建东,陈佰鸿,王利军,等. 葡萄抗寒性生理指标筛选及其评价[J]. 西北植物学报,2010,30(11):2 232-2 239.
CAO Jian-dong, CHEN Bai-hong, WANG Li-jun, et al. (2010). Cold resistance indexes identification and comprehensive evaluation of grape varieties [J].ActaBot.Boreal, 30(11): 2,232-2,239. (in Chinese)
[15] 白茹,高登涛,刘怀锋,等. 引入石河子地区的葡萄砧木抗寒性研究[J]. 新疆农业科学,2015,52(2):210-216.
Fund project:Supported by Special Fund for the Technical System of Modern Agricultural Industry (CARS-30-ZP-03); Financial Forestry Science and Technology Special Fund Project of Xinjiang Uygur Autonomous Region "Study on the cold resistance of grape special stock and its utilization technology"
BAI Ru, GAO Deng-tao, LIU Huai-feng, et al. (2015). Study on cold resistance of grapexine rootstocks introduced in shihezi area [J].XinjiangAgriculturalSciences,52(2):210-216. (in Chinese)
Comparison of Cold Resistance of Grape Rootstock Branches
ZHONG Hai-xia, PAN Ming-qi, ZHANG Fu-chun, ERMEK Cai Kasimu, HAN Shou-an,
ZHANG Wen, XIE Hui, CHEN Rui, MEI Chuang, WU Xin-yu
(ResearchInstituteofHorticulturalCrops,XinjiangAcademyofAgriculturalSciences/ScientificObservingandExperimentalStationofPomology(Xinjiang),MinistryofAgriculture,Urumqi830091,China)
Abstract:【Objective】 In order to provide a theoretical basis of cold resistance for choice, introduction and cultivation of cold resistance of Xinjiang grape rootstock, this paper aims to discuss the cold resistance of grape rootstock branches.【Method】Taking the 8 varieties of grape rootstock as research subjects, and relative electrolyte conductivity, free proline, MDA, soluble sugar, soluble protein content, the activities of SOD, POD and CAT were investigated on varieties of grape rootstock branches in different low temperature stress. And subordinate function method was applied to the physiological and biochemical indexes of cold resistance comprehensive evaluation.【Result】The relative electrolyte conductivity, free proline, MDA and soluble sugar of all grape rootstock varieties gradually increased in lowering of the temperature, but the increasing value was different. The content of soluble protein, SOD and POD increased first and then declined, and CAT content was showing irregular change. According to the comprehensive evaluation with subordinate function, the order of regarding the cold resistance strength of grape rootstock varieties was detoxification beta > 5C > 110R > 101-14 > 188-08 > 3309C > 5BB > SO4. According to restore growth's eduction and the cold hardiness strength of each type method of membership grade function, the physical and chemical indexes root stock of hardiness sort results were basically identical.【Conclusion】 The strongest cold resistance of rootstock was detoxification beta, which could resist the lowest temperature below -30℃, the cold resistance of 5C, 110R, 101-14, 188-08, 3309C and 5BB were much stronger which could resist the lowest temperature -20- -30℃, and SO4 cold resistance was the weakest which could resist the lowest temperature -20℃.
Key words:low temperature stress; grape rootstock; cold resistance; comparison
通讯作者:潘明启(1962-),男,副研究员,研究方向为葡萄栽培与质量控制,(E-mail)panmq3399@sohu.com
作者简介:钟海霞(1988-),女,助理研究员,硕士,研究方向为葡萄栽培与生理,(E-mail)zhonghaixia1@sina.cn
基金项目:现代农业产业技术体系专项资金(CARS-30-ZP-03);自治区财政林业科技专项资金项目“葡萄专用砧木抗寒性及利用技术研究”
收稿日期:2015-06-08
中图分类号:S663.1
文献标识码:A
文章编号:1001-4330(2016)02-0261-09
doi:10.6048/j.issn.1001-4330.2016.02.010
