不同类型猴樟混交林种间关系和冠层结构
2016-02-20韦小丽陈瀚林
韦小丽,陈瀚林
(1.贵州大学 林学院,贵州 贵阳 550025;2.贵州习水县林业局,贵州 习水 564600)
·专家特稿·
不同类型猴樟混交林种间关系和冠层结构
韦小丽1,陈瀚林2
(1.贵州大学 林学院,贵州 贵阳 550025;2.贵州习水县林业局,贵州 习水 564600)
以贵州人工栽培的猴樟纯林和3种类型的混交林为研究对象,采用Hegyi单木竞争指数模型对其种内、种间及全林分竞争强度进行定量研究,分析了不同混交组成树种的生态位宽度、生态相似系数和生态位重叠并测定了林分的冠层结构参数。结果表明:猴樟纯林的竞争最强,猴樟×杜英×楠木混交林是一个生态相似性较大、生态位重叠较高的混交组合,但目前竞争最小;猴樟×广玉兰×女贞混交林则是一个生态相似性小、生态位重叠较低且竞争比较强的混交组合,需要进行结构调整。研究结果反映出了不同混交组合中物种生态位宽度、生态位重叠和种间竞争关系的复杂性。混交林的冠层结构参数因树种组成、混交比例不同而异,总体上混交林的冠层参数空间异质性大于纯林。
猴樟;混交林;竞争强度;冠层结构
混交林种间关系是树种间通过复杂相互作用而对彼此产生利害作用的最终结果,种间关系的发展演变直接影响到混交林营造的效果。因此,混交林种间关系的研究一直是混交林的研究热点。前人对混交林种间关系的研究主要涉及混交林种间养分关系[1]、林木的种内和种间竞争关系及生态位关系[2-6]等方面,这些研究为混交林的组织经营、树种配置及种间关系的调节奠定了基础。
猴樟(CinnamomumbodinieriLevl.)是我国特有植物,在亚热带常与其它树种形成针阔混交或阔叶混交林,是一个集用材、园林绿化、化工原料、医药用途于一身的多用途树种。前人对猴樟的苗期生长规律、生态适应性、不同地理种源苗期生物学特性[7-10]等均做过细致研究。猴樟原生于天然混交林,培育混交林无疑是一个理想的选择。猴樟人工混交林在贵州遵义、修文,贵阳市白云区等有栽培,但这些混交搭配是否适宜尚不清楚。本文采用典型样地调查法,对贵州现有的猴樟×女贞(Ligustrumlucidum)×广玉兰(Magnoliagrandiflora)、猴樟×杜英(Elaeocarpussylvestris)×楠木(Phoebezhennan)、猴樟×马尾松(Pinusmassoniana)×楸树(Catalpabungei)和猴樟纯林的种间竞争关系、生态位及冠层结构特征进行研究,以探索不同混交组合种间关系的适宜性,为猴樟混交林培育提供理论依据和技术指导。
1 研究地点概况
分别在贵州遵义市红花岗区和贵阳市白云区进行样地调查。两地均属于亚热带季风气候,遵义市红花岗区地处东经105°36′~108°13′,北纬27°8′~29°12′,贵阳市白云区的地理位置为东经106°43′,北纬26°43′,两地的年均温度分别为15.1℃、15.3℃;绝对最高温分别为38.7℃,35.1℃,最低温分别为-7.1℃, -7.3℃;年均相对湿度分别为79%、77%;年降水量1000~1300 mm, 全年无霜期270~300天。可见两地的自然条件相似。两地的林分均为人工栽植的11年生林分,行状混交。各调查点林分概况见表1。
2 研究方法
2.1 样地调查
本次调查共涉及4个林分类型,即猴樟×马尾松×楸树、猴樟×杜英×楠木、猴樟×女贞×广玉兰和猴樟纯林,每个类型设置3个重复样地,总共12个样地。选择样地时,尽量选择林相完好,立地条件基本一致,有代表性的地段为标准地,标准地面积为400 m2。在样地内进行每木检尺,记录树号、树种,实测树高、胸径、地径、坐标值。生态位研究参照范迎新等[6]的方法,以样地东西方向为x轴,南北方向为y轴,确定样地内每株树的空间位置。根据每株树的空间位置,在进行生态位计算时将每块固定样地再拆分成4个l0×10 m2的小样方。
2.2 竞争强度(竞争指数)测定
在调查样点的林分中,随机选择30株猴樟作对象木,以该对象木为中心,将半径6 m(包括6 m)范围内的所有乔木(胸径≥2 cm,包括猴樟)定义为竞争木,并测定与对象木的距离,利用胸径和竞争木与对象木的间距求算竞争指数,从而确定种内种间的竞争关系。
2.3 林分冠层参数测定
采用CI110冠层图象分析仪进行测定。测定时间为2013年7月份,在每个林分样地中随机测定5个样点。将仪器鱼眼镜头放在样点内地表上1 cm处拍摄林冠层影像,将数字图象输入计算机后用CI110 软件获得半球灰度图象,计算出叶面积指数(LAI)、平均叶片倾角(MLA)和透光率等参数。
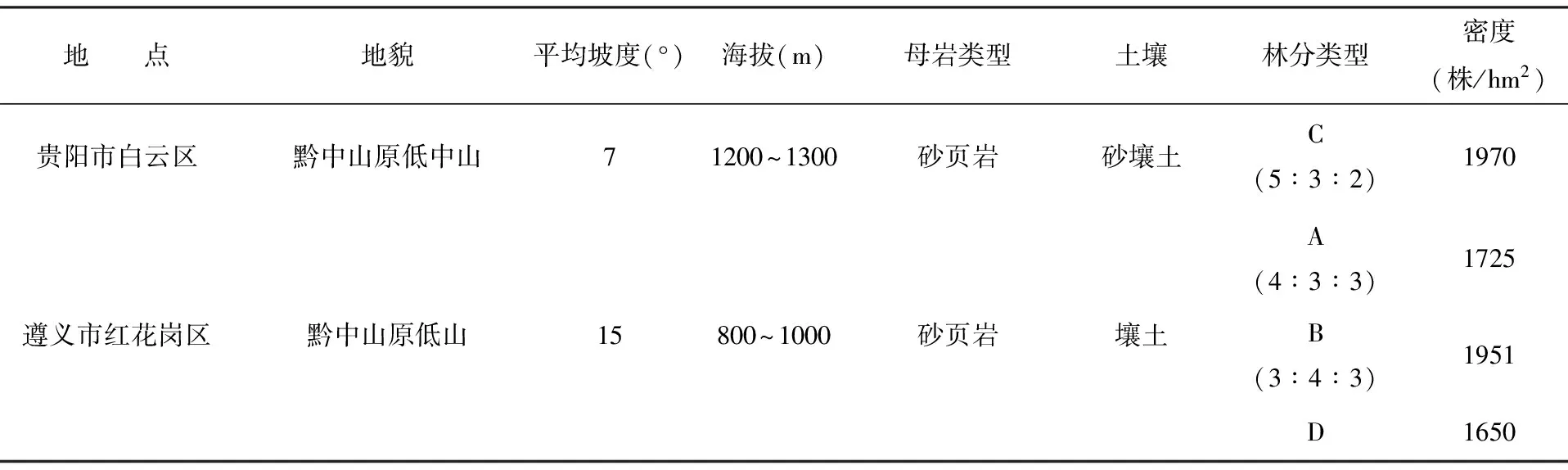
表1 各调查区域的林分情况Tab.1 Thebasic conditions of investigation sites
注:表中A、B、C、D分别代表猴樟×杜英×楠木、猴樟×女贞×广玉兰、猴樟×马尾松×楸树混交林和猴樟纯林,下同;括号中数字为混交比例。
2.4 数据计算及分析
2.4.1 竞争指数计算
采用Hegyi提出的单木竞争指数模型[2]来计算竞争指数的大小,其计算公式为:
CI为竞争指数(Competition index),其值越大竞争越激烈,反之竞争越小;Di为对象木i的胸径;Dj为竞争木j的胸径;Lij为对象木i与竞争木j之间的距离,N为竞争木的株数。
2.4.1 生态位宽度
采用Shannon-Weinner公式计算:
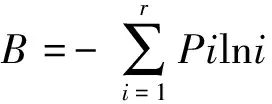
式中:B为物种的生态位宽度;Pi为物种在一个资源序列中第i位的重要值该种重要值总数的比例;r为资源位数(小样方数)。
2.4.2 生态位相似性比例
生态位相似性比例计算公式:
式中:Cih表示物种i与物种h的相似程度, 且有Cih=Chi, 具有值域[0 , 1 ] ;Pij,Phj分别为物种i和物种h在资源位j上的重要值百分率,r为资源位数(小样方数)。
2.4.2 生态位重叠
用Levins 重叠指数计算生态位重叠值:
式中Lih为物种i重叠物种h的生态位重叠指数,Lhi为物种h重叠物种i的生态位重叠指数; B(L)i为物种i的Levins生态位重叠度指数, B(L)h为物种h的Levins生态位重叠指数, B(L)i和B(L)h具有值域值[1/r,1] ;Lih和Lhi具有值域值[0,1]。
2.5 统计分析
采用SPSS version 17.0 统计软件进行统计分析,采用最小显著差法( LSD) 进行多重比较。
3 结果与分析
3.1 不同类型猴樟混交林的种间和种内竞争强度
根据调查结果,以猴樟为对象木分别计算了不同混交类型猴樟与其它林木的种间竞争指数和猴樟种内的竞争指数及全林分竞争指数(表2)。3种混交类型中,种间竞争关系以猴樟与女贞、广玉兰竞争最为激烈,两者的平均竞争指数均>2.2,猴樟和马尾松的竞争指数最小,平均竞争指数为1.131,表明两者之间的竞争最弱。4种林分类型中,猴樟的种内竞争关系最强的是猴樟×马尾松×楸树混交林,其次为猴樟纯林,最小的为猴樟×女贞×广玉兰混交林。猴樟×马尾松×楸树混交林中猴樟种内竞争最强是因为猴樟与马尾松的竞争较弱,与楸树竞争处于中等水平,而马尾松、楸树树冠较窄,对猴樟影响较小,在有限的环境中,林分内的竞争重点就转向了种内竞争,因而其竞争指数反而高于纯林。从全林分的竞争指数看,三种混交林中猴樟×马尾松×楸树混交林的竞争最激烈,猴樟×女贞×广玉兰混交林次之,猴樟×杜英×楠木竞争最小,从三树种的搭配看猴樟×杜英×楠木混交关系较适宜。
3.2 对象木、竞争木与竞争指数的相关性分析
以竞争指数为因变量(y),对象木胸径(x1)、竞争木胸径(x2)、对象木与竞争木距离(x3)为自变量,进行了逐步回归分析(表3),结果表明:大多数情况下对象木与竞争木之间的平均竞争指数与对象木胸径(x1)、竞争木胸径(x2)、对象木与竞争木距离(x3)的相关性达极显著水平(P<0.01),可以用拟合的回归方程预估种间竞争关系。三个自变量中,竞争木胸径(x2) 、对象木与竞争木距离(x3)与平均竞争指数的偏相关关系大多数均达到极显著水平(P<0.01),呈典型的负相关,即随着竞争木胸径增大,竞争木与对象木距离越远,对象木与竞争木的竞争关系越弱。除猴樟×杜英×楠木混交林中猴樟与楠木、猴樟种内和猴樟×马尾松×楸树混交林中猴樟与马尾松的竞争指数与对象木相关性不明显外,其它情况下对象木胸径与平均竞争指数均呈正相关,即对象木胸径越大,其占据的生态空间越大,竞争能力越强。
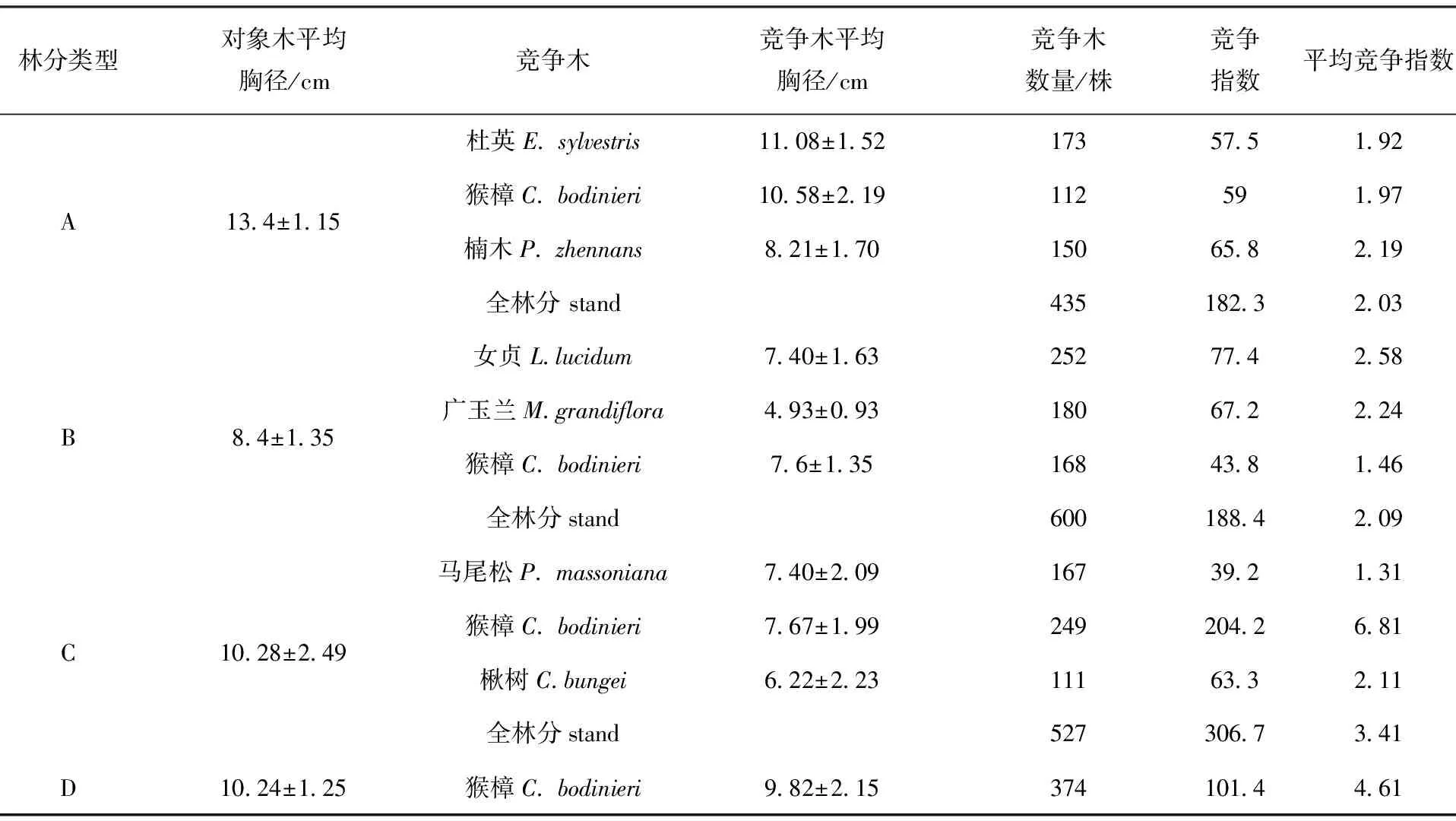
表2 不同林分类型种间、种内竞争指数Tab.2 The competitive indices(CI) between species and within species for difference stand types
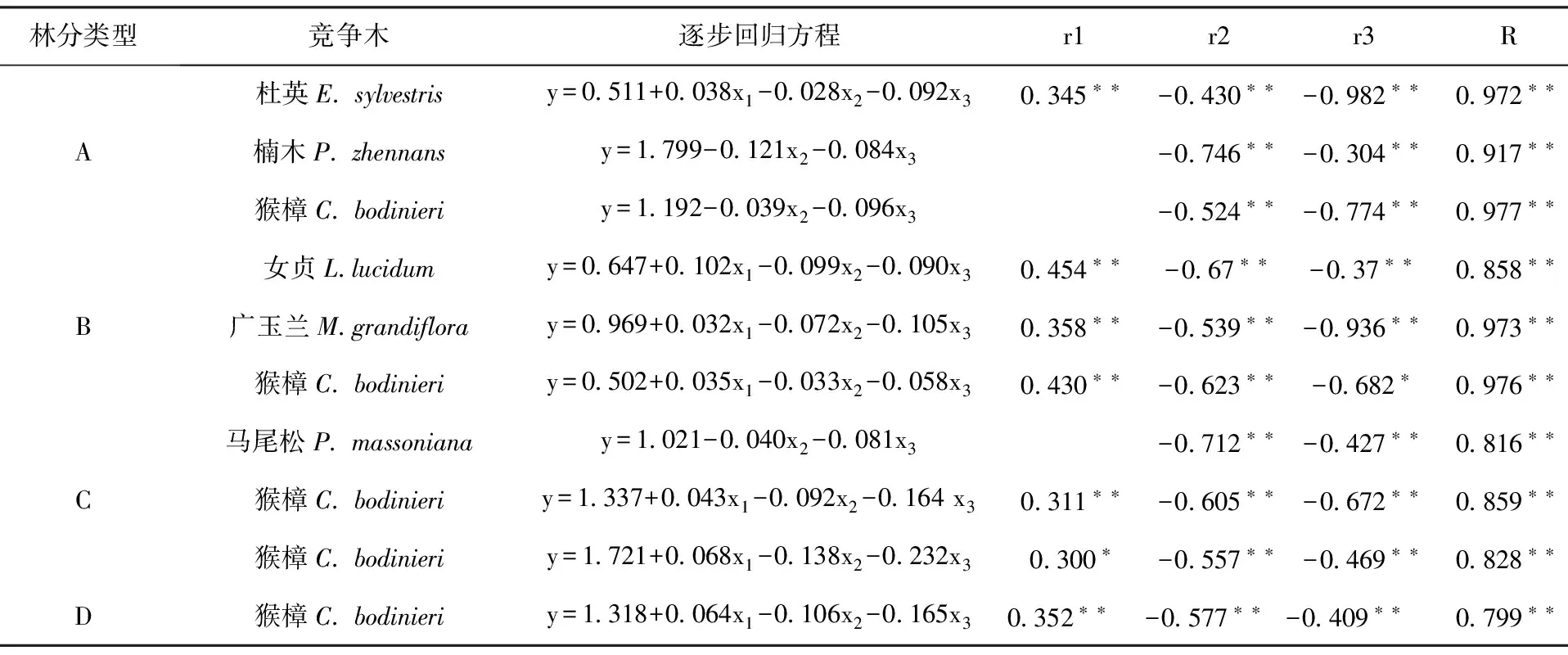
表3 对象木、竞争木与竞争指数的相关性分析结果Tab.3 The results of correlation analysis between object tree,competition tree and CI
注: R、r1、r2、r3分别表示竞争指数(y)与对象木胸径(x1)、竞争木胸径(x2)、对象木与竞争木距离(x3)之间的复相关系数和偏相关系数;“*”、“**”分别表示在0.05、0.01水平上相关性显著。
3.3 不同林分类型组成树种的生态位宽度与生态位重叠
3.3.1 生态位宽度
由图1可知,在不同类型混交林中,猴樟×杜英混交林中猴樟的生态位宽度值最大,为2.27,说明该混交类型中猴樟对资源环境有较强的适应性,猴樟的生态位宽度值最小的是猴樟×广玉兰×女贞混交类型,说明在这种混交环境下,受到生态位宽度大的女贞的竞争,猴樟利用资源的能力相对较弱。在该组合中广玉兰的生态位宽度最窄,仅为1.18,说明广玉兰受到猴樟女贞的强烈竞争,可利用的资源空间最小;在猴樟×杜英×楠木混交林中,生态位宽度表现为猴樟>杜英>楠木,但三者差异不大,表明三者混交,各树种对资源空间的利用均均衡;在猴樟×马尾松×楸树混交林中,猴樟的生态位宽度最大,马尾松、楸树生态位宽度较接近。
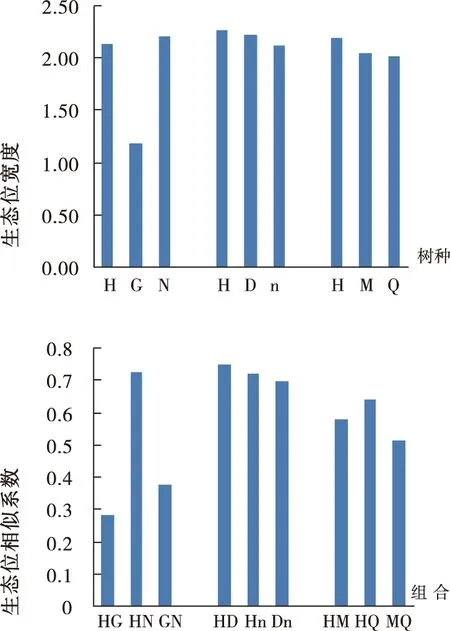
图1 不同混交林中物种的生态位宽度和生态位相似性Fig.1 The niche breadth and niche similarity between species in different mixed stand
注:图中H、G、N、D、n、M、Q分别表示猴樟、广玉兰、女贞、杜英、楠木、马尾松、楸树,HG 、HN、GN 、HD、Hn、Dn、HM、HQ、MQ分别表示猴樟与广玉兰、猴樟与女贞、广玉兰和女贞、猴樟和杜英、猴樟和楠木、杜英和楠木、猴樟和马尾松、猴樟和楸树、马尾松和楸树组合,下同。
3.3.2 生态位相似系数
从图1可以看出,生态位相似系数在0.7以上的有猴樟与楠木、猴樟与杜英及猴樟与女贞3个种对,说明这些种对之间对资源的需求具有很大相似性。猴樟与广玉兰、广玉兰与女贞的生态位相似系数小于0.4,说明它们之间对环境的需求有较大的差异。其它种对的生态位相似系数在0.5~0.7之间。猴樟×杜英×楠木混交林是一个生态相似性较大的混交组合,而猴樟×广玉兰×女贞混交林是一个生态差异较大的组合。
3.3.3 生态位重叠
生态位重叠反映种群之间对资源利用的相似性程度和竞争关系。从表4可以看出,虽然3种混交组合中不乏生态位宽度较大的树种,但在整体上3种混交林中物种两两间生态位重叠都不大,其Lhi和Lih值均小于0.1,这可能是因为林分是人工营造的,人为进行的种植点配置降低了物种间的生态位重叠。3种混交林中物种间的生态位重叠因样地不同存在一定的差异,这是由于环境资源位的异质性所致。在混交林中物种间的生态位重叠呈现出现三种情况:第一种是两种生态位宽度均较大的物种,其Lhi与Lih也较大,如猴樟×杜英×楠木混交林就属于这种情况,该混交组合中物种间的生态位重叠整体上大于其它组合。第二种是两物种生态位宽度均大,但其Lhi和Lih值较小,如猴樟与女贞(HN);第三种是生态位宽度大的物种和生态位宽度小的物种组合,但其Lhi和Lih值较大,典型的如猴樟和广玉兰,均变动在0.07左右,表明两物种在同一资源位上出现的频率相当,利用资源的能力和要求的生境相似;且生态位宽度大的物种和生态位宽度小的物种组合,其生态位重叠均表现为Lhi>Lih,最典型的是女贞与广玉兰,女贞对广玉兰的重叠度高,而广玉兰对女贞的重叠度低;整体上猴樟与各物种的生态位重叠都呈现Lhi>Lih,这一方面与猴樟速生、适应性强的特性有关,另一方面与人为配置赋予其优势地位有关。
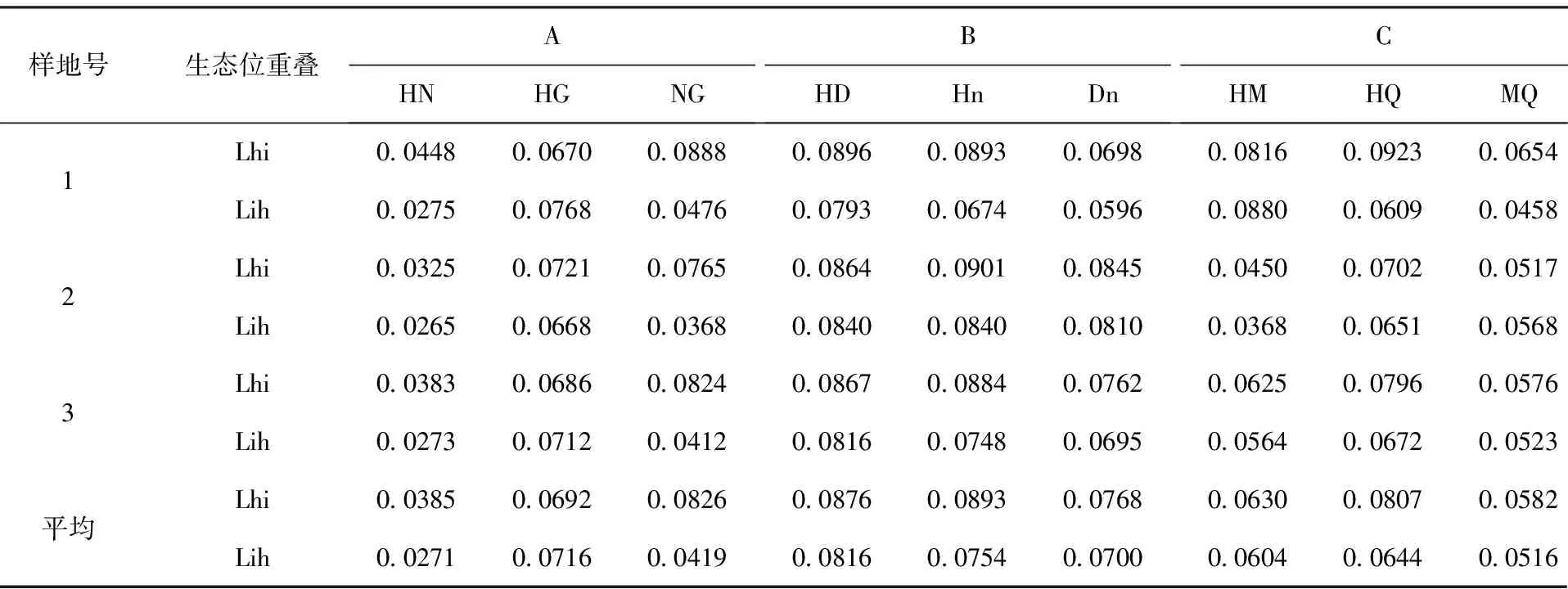
表4 不同混交类型物种间生态位重叠比较Tab.4 Comparison with the niche overlap between species of different mixed types
3.4 不同混交林林分叶面积指数、叶倾角及透光率比较
3.4.1 林分叶面积指数
叶面积指数(LAI) 是表征群落结构的重要特征参数之一[11]。由表5可知,4种林分类型中,猴樟×杜英×楠木的LAI最大,其次为猴樟纯林,猴樟×女贞×广玉兰的LAI最小,仅为猴樟×杜英×楠木混交林、猴樟纯林的45.1%和81.8%。方差分析表明,不同混交类型林分LAI差异显著(P<0.05)。4种林分类型中,猴樟×马尾松×楸树的LAI空间变异最小,猴樟×女贞×广玉兰混交林的LAI空间变异最大。
3.4.2 林分的叶倾角
平均叶倾角指叶表面垂线与铅垂线的夹角,自然界的多数植物能够以锐角截获阳光,这样可以避免直接辐射造成灼伤[11]。由表5可知,猴樟×马尾松×楸树混交林的平均叶倾角最大,为51.3°,其它三种林分叶倾角差异不大,在10~11°之间。方差分析表明,4种林分的平均叶倾角差异显著(P<0.05),但主要表现为猴樟×马尾松×楸树混交林与其它三种林分差异显著。从平均叶倾角的空间变异看,猴樟纯林最小,猴樟×杜英×楠木混交林平均叶倾角空间变异最大。
3.4.3 林分透光率
林分透光率和林分冠层结构、郁闭度有关,林分冠层越浓密,郁闭度越高,林分透光率越低,反之亦然。由表5可以看出,猴樟×马尾松×楸树混交林的透光率最高,分别是猴樟×女贞×广玉兰、猴樟×杜英×楠木和猴樟纯林的1.67倍、1.87倍和1.87倍。猴樟×马尾松×楸树混交林的透光率与其它三种林分差异显著(P<0.05),但其它三种林分之间差异不显著(P>0.05)。猴樟×马尾松×楸树混交林的透光率高是因为其叶倾角大,叶面积指数小。林分透光率的空间变异以猴樟纯林最大,最小的是猴樟×马尾松×楸树混交林。
4 结论与讨论
协调的种间关系是混交林营造成功的关键。协调的种间关系体现为混交树种生物生态学特性、养分吸收特性、生物化学关系等方面的互补与协调。本研究主要从树种生物生态学特性角度研究混交林的种间关系,结果表明:从混交树种两两间的竞争关系看,猴樟与女贞、广玉兰的竞争最为激烈,猴樟与楸树、杜英、楠木的竞争强度居中,猴樟与马尾松竞争最弱。从全林分看,猴樟纯林的竞争最强,猴樟×杜英×楠木竞争最小,是较适宜的混交组合。对于猴樟种内竞争关系看,存在两种情况,一是混交后猴樟种内竞争关系减弱,一种情况是混交后种内竞争关系增强,前者如在猴樟×女贞×广玉兰和猴樟×杜英×楠木混交林中的猴樟种内竞争指数均小于猴樟纯林;后者如猴樟×马尾松×楸树混交林,猴樟的种内竞争关系反而大于纯林。说明只有恰当的种间搭配才能减弱种内竞争。
前人的研究表明,竞争指数和对象木胸径之间呈幂函数关系,并建立了幂函数方程预估种间关系调节的时间[2]。本研究对竞争指数和对象木胸径进行拟合没有得到相同的规律。相反,有3组对象木与竞争木的竞争指数与对象木胸径之间的相关性不明显,逐步回归中该变量被剔除掉;这可能与研究的混交林对象不同、混交方式及其所处的年龄阶段有关。
混交树种能否在竞争中占据优势地位取决于树种对资源空间的利用能力,生态位宽度大的物种通常在竞争中占据优势地位。在3种混交类型中,猴樟的生态位都比较接近,在2.13~2.27之间;除广玉兰的生态位宽度较窄(1.14)外,其它树种的生态位宽度与猴樟比较接近,变动在2.01~2.23之间,说明这些树种与猴樟都有相近的资源空间利用能力。生态位宽度较大的种群生态位相似性比例值高, 在不同的资源位上对生境的需求和资源的利用有较大的相似性,而生态位宽度较小的种群之间生态位相似性比例值则很小[12]。本研究中猴樟×杜英×楠木混交林中三个树种的生态位宽度都较大,是一个生态相似性较大的混交组合,而猴樟×广玉兰×女贞混交林则是一个生态差异较大的组合。

表5 不同混交林林分叶面积指数、叶倾角和透光率比较Tab.5 Comparison with leaf area index, leaf angle and light ransmittance of different mixed stand
生态位宽度与生态位重叠值有一定的联系, 一般生态位宽度大的物种, 其生态重叠值较大, 生态位较窄的树种之间及其与生态位较宽的树种之间生态位重叠值较小[13-14]。张桂萍等[12]发现生态位宽度较小的种群间也可以出现较大的生态位重叠。本研究中混交树种的生态位重叠呈现出三种情况:(1)生态位宽度大生态位重叠也较大;(2)生态位宽度大生态位重叠小;(3)生态位宽度大的和生态位宽度小的物种呈现出较大的生态位重叠,且均表现为Lhi>Lih。这些均反映了自然界物种对环境资源位利用的多样性,不同的物种组合呈现出不同的资源利用方式,第二种情况应该是比较理想的混交组合。尽管两个物种生态位都较大,但生态位重叠小,各自利用了不同的环境资源位。
本研究中猴樟与女贞生态位重叠属于第二种情况,但从目前的竞争强度看却较大,似乎存在矛盾。事实上,对于生态位重叠是否会导致种间激烈的竞争一直存在争论。Colwell 和Futuyma 、Hespenheide 、Heck、Hurlbert、Abrams 等认为生态位重叠与竞争无关,而Pianka 等认为竞争与生态位重叠是密切相关的[15]。但至少认为有生态位重叠就一定有竞争的观点是不全面的[16]。在自然界,生态位经常发生重叠但并不表现有竞争排除现象,生态位重叠本身显然并不一定伴随着竞争,如果资源很丰富,两种生物就可以共同利用同一资源而彼此并不给对方带来损害。本研究中在猴樟×杜英×楠木混交林中虽然3个树种间的生态位重叠均较高,但三者混交形成的林分竞争强度却较其它林分小。在猴樟×女贞×广玉兰混交林中,猴樟与女贞的生态位重叠最小,但其竞争强度却较大。因此认为,对重叠较大的种对,存在两种可能:一是树种间共享资源的同时存在竞争关系,二是树种间资源利用相似的同时,彼此促进,共生共荣[5]。对于重叠度较小的种对如发生了较激烈的竞争则是因为混交比例、混交方式配置不当导致资源空间不足,说明需要进行结构调整缓和种间矛盾。
林冠结构是植物用以适应环境和提高群体光合效能所采取的一种生态对策, 是群落长期演变过程的生理生态特征[17]。营造混交林有助于形成复层冠层结构,可提高对资源空间的利用率和改善林地环境条件,从而促进林木生长。因此,通过合理的混交组合、混交比例和混交方式,培育结构和功能高效的冠层结构是混交林成功培育的理论基础。叶面积指数是一个与树种组成、栽植密度、种植点配置紧密相关的参数,它决定了林冠的光合作用、蒸腾作用和林分生产力。有研究得出结论,混交林的LAI大于纯林,且混交林的LAI的空间异质性高于纯林[18]。本研究中猴樟×杜英×楠木的LAI最大,但其它两种混交林的LAI略小于猴樟纯林;除猴樟×马尾松×楸树混交林的LAI空间异质性较低外,其它均高于猴樟纯林,与前人的结论基本一致。
植物群体光分布和传递都以叶倾角为基础,是植物群体结构中的重要参数。很大程度上,叶倾角受植物本身特性决定。柴中林认为树木的叶倾角与树木的叶柄长、节间长和叶面积相关,并建立了反映了叶面积与叶柄长、节间长、叶倾角之间的关系模型[19]。目前尚无培育技术措施对林分叶倾角影响的研究。本研究中猴樟×马尾松×楸树混交林的平均叶倾角最大,其它三种林分叶倾角差异不大,在10~11°之间,这与针叶树种马尾松的存在有关,与阔叶树种相比,大多数针叶树叶片倾角都较大。整体看,混交林的叶倾角空间异质性较大。由于林分LAI和叶倾角不同,林分透光率表现出明显的差异。猴樟×马尾松×楸树混交林LAI小,平均叶倾角大,因而其透光率最高。
本研究对现有猴樟混交林的竞争关系和冠层结构进行了探讨,得出了一些关于猴樟混交林的初步的认识。但由于现存林分不是经过严格试验设计的,因此得出的结论具有一定的局限性。在本研究的基础上,还需要进一步解决的问题有:(1)上述树种按不同混交比例、不同混交方式两两组合后的种间竞争与调节;(2)培育措施对林分叶倾角、LAI的影响,什么样的LAI、叶倾角更适合于提高林分截获光能、提高光能利用率和改善林地环境,如何降低冠层结构的空间异质性等,都待进一步研究。
[1] 蒋三乃,翟明普,贾黎明.混交林种间养分关系研究进展[J].北京林业大学学报,2001,23(2):72-77.
[2] 封 磊,洪 伟,吴承祯,等.杉木一观光木人工混交林种内和种间竞争研究[J].西北植物学报,2008,28(1):141-146.
[3] 林思祖,黄世国,洪 伟.杉阔混交林杉木与其混交树种种间竞争研究[J].林业科学,2004,40(2):160-164.
[4] 陶 岩,殷秀琴,田育红,等.长白山红松针阔混交林种内、种间竞争关系研究[J].北京师范大学学报(自然科学版),2011,47(4):428-431.
[5] 陈存及,陈新芳,刘金福,等.人工—天然杉阔混交林种群生态位及竞争研究[J].林业科学,2004,40(1):78-83.
[6] 范迎新,李传荣,许景伟,等.沙质海岸人工混交林种群竞争及其生态位[J].中南林业科技大学学报,2008,28(2):40-50.
[7] 韦小丽,熊忠华.香樟和猴樟1年生播种苗的生长发育规律[J].山地农业生物学报,2005,24(3):205-208.
[8] 韦小丽.不同光环境下香樟、猴樟苗木的生态适应[J].山地农业生物学报,2003,22(3):208-213.
[9] 田小琴,韦小丽.生态环境对贵州猴樟人工林生长的影响[J].贵州农业科学,2011,39(4):184-186.
[10] 王 娈.不同地理种源猴樟苗期生物学特性研究[D].贵阳:贵州大学,2011.
[11] 宋子炜,郭小平,赵廷宁,等.北京市顺义区公路绿化植物群落的光环境特性[J].生态学报,2008,28(8):3779-3788.
[12] 张桂萍,张 峰,茹文明.山西绵山植被优势种群生态位研究[J].植物研究,2006,26(2):176-181.
[13] 邓嘉农,黄 茹,何丙辉,等.璧山县生态恢复过程中水土保持林主要乔木树种生态位特征研究[J].西南大学学报(自然科学版),2011,33(7):103-108.
[14] 陈子林,张志祥,刘 鹏.浙江六十田常绿阔叶林主要乔木种生态位研究[J].中南林业科技大学学报,2007,27(6):77-82.
[15] 杨效文,马继盛.生态位有关术语的定义及计算公式评述[J].生态学杂志,1992,11(2):44-49,35.
[16] 尚玉昌.现代生态学中的生态位理论[J].生态学进展,1988,5(2):77-84.
[17] 任 海,彭少麟.鼎湖山森林群落的几种叶面积指数测定方法的比较[J].生态学报,1997,17(2):220-223.
[18] 贾小容,苏志尧,区余端,等.三种人工林分的冠层结构参数与林下光照条件[J].广西植物,2011,31(4):473-478.
[19] 柴中林,王兰州.树木叶片面积与叶柄长、节间长与叶倾角的关系初探[J].生物数学学报,2009,24(2):329-334.
Theintraspecific relationship and canopy structure of different typeCinnamomumbodinierimixed stand
WEIXiao-li1,CHENHan-lin2
(1.ForestrycollegeofGuizhouuniversity,Guiyang,Guizhou550025,China;2.ForestryBureauofXishuicounty,Xishui,Guizhou564600,China)
Pure plantation and three types mixed plantation ofCinnamomumbodinieriin Guizhou was chosen as researh object, and the competition intensities of intraspecific, interspecific and total stand in quantitatively was analyzed by applying Hegyi'Scompetition index model for individual tree. The niche breadth, niche similarity and niche overlap were analyzed to determine the canopy structure parameter. The results showed that competition intensities in the pureC.bodinieristand was the largest. Mixed stand ofC.bodinieri,E.sylvestrisandP.zhennanswas the combinination of high niche similarity and niche overlap, but with weak competitiveness. Mixed stand ofC.bodinieri,L.lucidumandM.grandiflorawas the one with low niche similarity and niche overlap, but competited strongly so that it was necessary to adjust the structure. The results reflected the complexity between niche breadth, niche overlap and interspecific relationship of species for different mixed combination. The canopy structure parameters of mixed stand were different with tree composition, mixed rate. The spatial heterogeneity of canopy parameters of mixed stand was obviously larger than pure stand.
Cinnamomumbodinieri; Mixed Stand; Competition Intensity; Canopy Structure
2016-07-24;
2016-10-11
“阔叶乡土树种猴樟培育关键技术研究与示范”(黔科合NY字[2009]3061)。
Q145
A
1008-0457(2016)06-0001-08 国际
10.15958/j.cnki.sdnyswxb.2016.06.001
