斑节对虾南海1号和非洲品系自交、杂交群体的形态差异分析
2016-02-08蔡云川江世贵孙苗苗黄建华杨其彬杨丽诗邱丽华周发林
蔡云川,江世贵,孙苗苗,黄建华,杨其彬,姜 松,杨丽诗,邱丽华,周发林
(1.广东省海洋与渔业技术推广总站,广东 广州 510220;
2.中国水产科学研究院南海水产研究所/农业部南海渔业资源开发利用重点实验室,广东 广州 510300;
3.连云港市海洋与水产科学研究所,江苏 连云港 222044)
斑节对虾南海1号和非洲品系自交、杂交群体的形态差异分析
蔡云川1,江世贵2,孙苗苗3,黄建华2,杨其彬2,姜 松2,杨丽诗2,邱丽华2,周发林2
(1.广东省海洋与渔业技术推广总站,广东 广州 510220;
2.中国水产科学研究院南海水产研究所/农业部南海渔业资源开发利用重点实验室,广东 广州 510300;
3.连云港市海洋与水产科学研究所,江苏 连云港 222044)
以斑节对虾南海1号(N)和非洲品系(F)作为亲本,进行自交与杂交,获得4个子代群体NN(N♀×N♂)、NF(N♀×F♂)、FN(F♀×N♂)和FF(F♀×F♂)。运用多元分析方法,采用7个测量性状对其形态进行比较研究。主成分分析结果显示,共构建了3个主成分,累计贡献率为68.35%;判别分析结果显示,自交群体的准确判别率相对较高,杂交群体的正确判别率较低;聚类分析结果显示,NN独立聚为一支,FF与NF聚为一小支后与FN聚为一支。
斑节对虾;自交;杂交;形态特征;多元分析
斑节对虾(Penaeus monodon)俗称草虾、竹节虾,分类学上隶属于节肢动物门(Arthropoda)、甲壳纲(Crustacea)、十足目(Decapoda)、游泳亚目(Natantia)、对虾科(Penaeidae)、对虾属(Penaeus),是对虾属中最大型种。斑节对虾是世界上三大对虾主要养殖品种之一,也是中国南方沿海诸省的重要养殖对象。优良品种是养殖业健康持续发展的关键因素之一,因此对虾育种得到世界各国的高度重视,并培育出一系列具有应用价值的良种。南海水产研究所于“十五”期间,在攻克了斑节对虾全人工繁育技术难题后,于2005年启动了斑节对虾的遗传育种。经过10年多努力,建立了斑节对虾遗传育种技术体系,培育出了斑节对虾南海1号新品种、非洲品系、抗氨氮品系以及适应低蛋白的家系等一批新品系和家系[1-6]。
经济性状是对虾遗传育种的重要指标,形态参数对于经济性状的贡献率是选育参数的重要依据。形态特征是物种遗传特性的外在表现,是遗传多样性最直接的体现,是进行群体结构鉴定的有效手段[7-13],具有研究手段直接且经济等优点[14]。近年来,多元分析被用于鱼类[15]、甲壳类[16-19]、贝类[20]的形态特征分析而采用多元分析方法分析虾类形态差异的相关研究也较多。李朝霞等[21]比较分析了中国对虾黄海1 号选育群体与野生群体的形态特征差异;郭惠等[22]分析了不同罗氏沼虾种群形态差异;赵晓勤[23]分析了日本沼虾种群的形态差异;孙苗苗等[24]分析了斑节对虾4 个地理种群自交与杂交F1的形态特征。本研究从形态学角度入手,以主成分分析、判别分析和聚类分析3种多元分析方法比较和分析了斑节对虾南海1号选育群体和非洲品系的自交与杂交子代的形态特征差异,旨在为其种群鉴定、亲缘关系研究以及后续的良种选育提供参考。
1 材料与方法
1.1 试验材料
南海1号:2005年,以我国南海海南岛三亚、临高、文昌和泰国南部(普吉岛)海域野生斑节对虾为基础种群,经过5代的连续群体选育,获得了快速生长新品种1个,即南海1号,并建立相应的核心种质群体。
非洲品系:2009—2010年,从非洲南部的坦桑尼亚引进斑节对虾野生种质资源群体,经过多代家系和BLUP选育,培育出非洲品系,并建立相应的核心种质群体。
2015年在中国水产科学研究院南海水产研究所斑节对虾遗传育种中心,以斑节对虾“南海1号”和非洲品系作为亲本,进行自交与杂交,获得两个自交群体NN(N♀×N♂)、FF(F♀×F♂)和两个杂交群体NF(N♀×F♂)、FN(F♀×N♂)。为消除环境因素对斑节对虾形态的影响,对4个群体斑节对虾荧光标记后混养于同一个池塘中。供试对虾样本的数量和规格见表1。
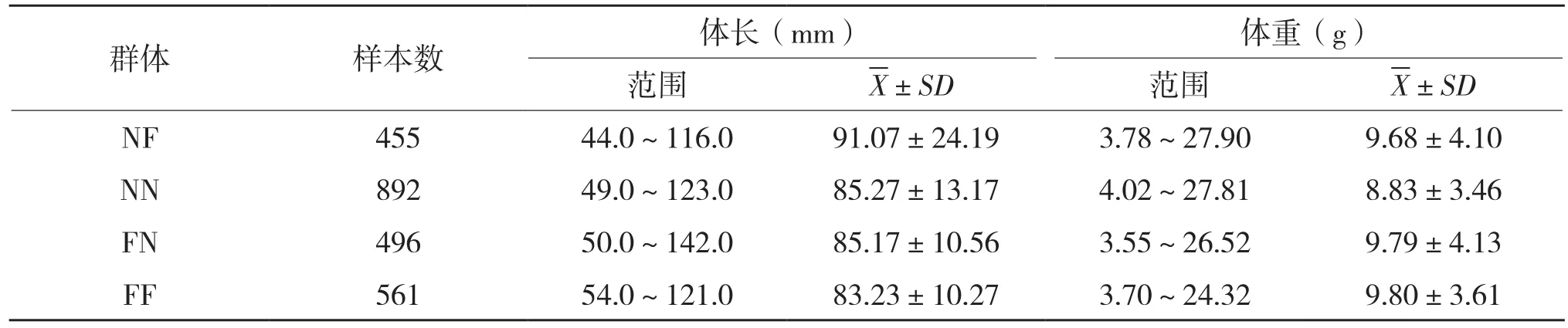
表1 斑节对虾4个群体样本数量和规格
1.2 4个群体形态性状的测量
通过随机抽样,利用游标卡尺测量了两个自交群体NN、FF和两个杂交群体NF、FN的7个形态学性状,分别是体长(BL)、头胸甲长(CL)、头胸甲高(CH)、头胸甲宽(CW)、第一腹节长(FSL)、第二腹节长(SSL)、第六腹节长(SISL)。
1.3 数据分析
对4个群体的7个可量性状(体长、头胸甲长、头胸甲高、头胸甲宽、第一腹节长、第二腹节长、第六腹节长)数据用SPSS(version10.1,SPSS Inc.1999)分别进行形态差异分析。通过主成分分析,得出各主成分的得分、方差和累计方差贡献率。通过判别分析,选出对判别贡献较大的参数建立判别公式。根据4个群体7个性状数据间的欧氏距离进行聚类分析,并构建聚类图。
在进行数据分析前,每尾对虾的所有实测可量数据通过下列公式进行转化[11,24-26],以消除因样品个体规格不整齐对形态分析造成的影响。

式中,Ms为各性状转化后的标准化数据,Mo为各性状的实际测量值,Ls为头胸甲长测量值的平均数,Lo为头胸甲长测量值。
2 结果与分析
2.1 4个群体7个形态性状的方差分析
从表2可以看出,NF与NN除头胸甲外,体长、第一腹节长、第二腹节长和第六腹节长存在显著差异;NN与FF头胸甲高、体长这两个性状差异不显著,其他性状均有显著差异;FF与FN各性状无显著差异;NF与FF体长、第二腹节长、第六腹节长这3个性状间存在显著差异。
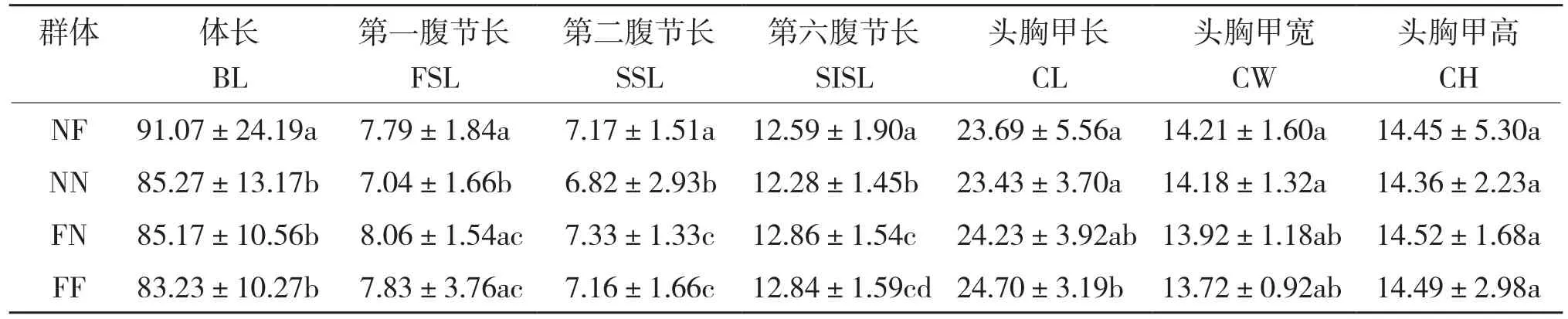
表2 斑节对虾不同群体间7个形态特征的差异
2.2 4个群体7个形态性状的主成分分析
对4个群体7个形态性状进行主成分分析,共获得3个主成分,其贡献率分别为27.88%、23.37%、17.10%,累计贡献率为68.35%。在第一主成分中,BL、CL和CW的影响最大,其负荷值分别为0.64、-0.83和0.89;第二主成分中作用最大的是FSL、SSL和SISL;第三主成分中作用最大的是CH(表3)。
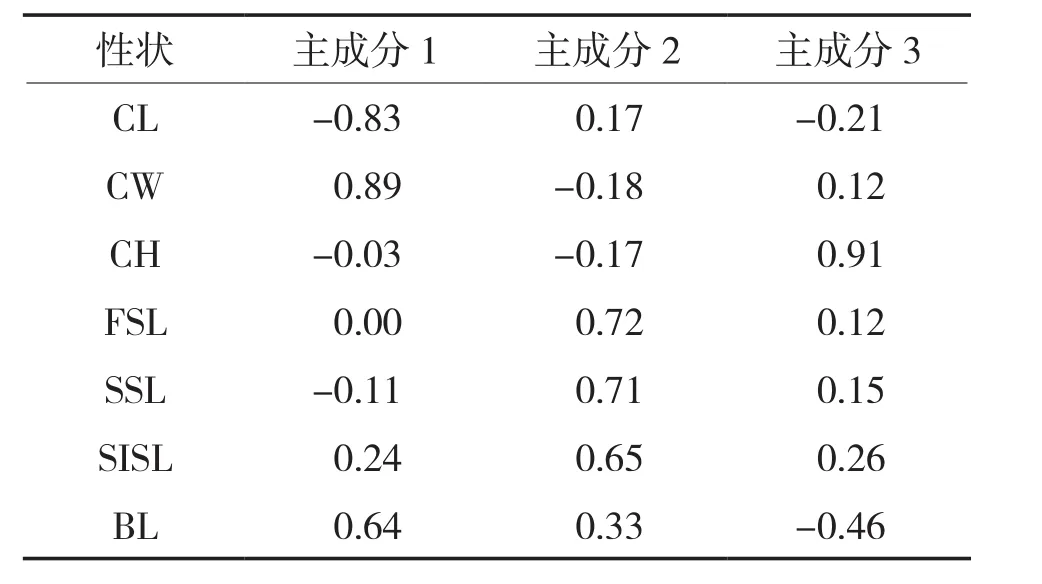
表3 斑节对虾7个性状对3个主成分的特性向量及主成分贡献率
2.3 4个群体7个形态性状的判别分析
利用逐步判别法,对7个形态性状进行分析,并建立了4个种群的判别函数式:

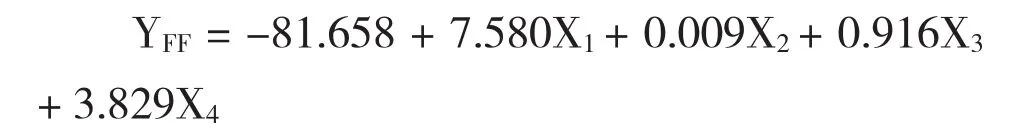
式中,X1~X4分别代表CW、CH、FSL、SISL。
要判断某尾虾的种群归属,只需将该虾7个形态参数测出并整理形成X1~X4,分别代入上述4个判别函数中,所得Y值最大的,则该虾属对应种群。
4个群体斑节对虾判别结果见表4,自交群体的准确辨别率相对较高,其中NN的准确辨别率最高为57.7%,FF准确辨别率为36.4%,杂交群体FN准确辨别率为33.2%,NF准确辨别率为11.9%。

表4 4个群体斑节对虾的判别结果
2.4 4个群体7个形态性状的聚类分析
对4个斑节对虾群体7个测量数据进行聚类分析,结果(图1)表明,4个群体斑节对虾可以明显聚为2大支:NN独立聚为一支,FF与NF聚为一小支后与FN聚为一支。
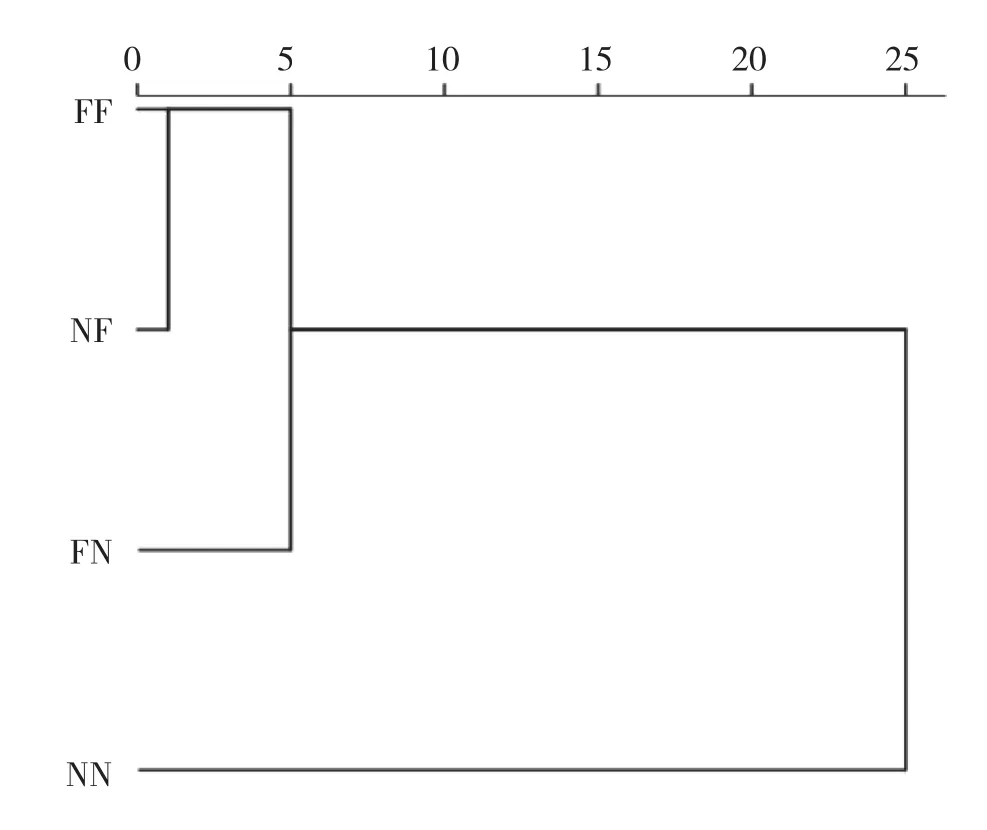
图1 4个群体斑节对虾的形态聚类分析结果
3 讨论
3.1 4个群体斑节对虾的形态差异
在物种系统分类研究中,地理隔离一直被认为是种及种下阶元形成的重要因素。Sun等[7]对印度尼西亚、泰国、非洲、三亚4个地理群体斑节对虾进行形态学分析,结果显示不同群体存在显著的形态变化,认为群体间形态特征存在差异是由地理因素造成的。孙苗苗等[24]运用多元分析方法对斑节对虾4个地理种群自交与杂交F1的形态特征进行了分析,认为杂交F1的性状是继承了其父母本的遗传信息,因此亲本的遗传特性在F1的形态变化上起到了重要作用。
本研究方差分析结果显示,两个自交群体NN与FF除头胸甲高、体长这两个性状外其他形态性状存在显著差异,说明这两个群体间在形态特征上具有较大区别。这可能与这两个自交群体NN与FF的初始亲本有关。斑节对虾海南1号初始亲本源自我国海南三亚周边3个自然群体及泰国南部(普吉岛)自然群体,经过多代选育获得的新品种。斑节对虾非洲品系初始亲本源自非洲南部海域自然群体,经过多代选育获得的新品系。Sun等[7]研究表明,非洲自然群体与三亚、泰国自然群体的形态特征存在显著差异。
3.2 斑节对虾形态特征多元分析
主成分分析在形态分析中广泛应用,可以将多个彼此相关的指标转换为新的、个数较少且互相独立或不相关的综合指标,便于统计分析[27]。研究表明,当主成分分析累计贡献率大于85%时,可以通过几个相互独立的因子来概括种群的形态差异[28]。本研究4个群体斑节对虾获得3个主成分,累计贡献率为68.35%,因此不能采用少数几个方差贡献率较大的因子来表现该4个群体的多个性状。但是通过分析可知,斑节对虾头胸甲的形态特征对其整体的影响较大,在贡献率较大的第一主成分中CL和CW的影响较大。
判别分析从7个特征性状中筛选出有显著贡献的4个变量CW、CH、FSL、SISL,可见头胸甲对斑节对虾形态特征有较大的影响作用,进一步证实了主成分分析的结果。4个群体的平均正确判别率为34.80%,处于较低水平。孙苗苗等[13]研究认为亲本的遗传特性在子代的形态变化上起重要作用。在本研究中,通过两个群体亲本间的自交与杂交产生的4个群体,其误判率较高。自交群体NN的正确辨别率(57.70%)显著高于其他3个群体,形态聚类结果依然显示其独立聚为一支,因此推测斑节对虾南海1号具有特有的形态特征,但这一结论需要进一步验证。
[1] 黄忠,江世贵,林黑着,等.5个斑节对虾半同胞家系生长、消化及免疫的比较[J].海洋渔业,2014(2):163-169.
[2] 姜松,杨其彬,黄建华,等.蛋白水平对不同斑节对虾家系生长与消化酶活性的影响[J].海洋渔业,2013(1):86-94.
[3] 黄建华,李永,杨其彬,等.斑节对虾家系氨氮耐受性的比较[J].南方水产科学,2012(6):37-43.
[4] 孙苗苗,黄建华,杨其彬,等.13个斑节对虾家系的生长及抗氨氮特性比较[J].上海海洋大学学报,2011(4):510-516.
[5] 王专伟,黄建华,杨其彬,等.15个斑节对虾家系生长及抗白斑病毒分析[J].海洋科学进展,2011(4):521-528.
[6] 全国水产技术推广总站.2010 水产新品种推广指南[M].北京:中国农业出版社,2011:178-205.
[7] Sun M M,Huang J H,Jiang S G,et al.Estimates of heritability and genetic correlations for growthrelated traits in the tiger prawn Penaeus monodon[J].Aquaculture Research,2013,doi:10.111/ are.12290.
[8] Tidu C,Sardá R,Pinna M,et al.Morphometric relationships of the European spiny lobster Palinurus elephas from northwestern Sardinia[J].Fisheries Research,2004,69(3):371-379.
[9] Konan K M,Adepo-Gourene A B,Ouattara A,et al.Morphometric variation among male populations of freshwater shrimp Macrobrachium vollenhovenii Herklots,1851 from Côte d’Ivoire Rivers[J].Fisheries Research,2010,103(1-3):1-8.
[10] Thomas H,Clay D.Morphometric and meristic differences between shallow and deep-water populations of white hake(Urophycis tenuis)in the southern Gulf of St Lawrence[J].Canadian Journal of Fisheries and Aquatic Sciences,1998,55(10):2274~2282.
[11] Ferrito V,Mannino M C,Pappalardo A M,et al.Morphological variation among populations of Aphanius fasciatus Nardo,1827(Teleostei,Cyprinodontidae)from the Mediterranean[J].Fish Biology,2007,70(1):1-20.
[12] Anvarifar H,Khyabani A,Farahmand H,et al.Detection of morphometric differentiation between isolated up- and downstream populations of Siah Mahi(Capoeta capoeta gracilis)(Pisces:Cyprinidae)in the Tajan River(Iran)[J].Hydrobiologia,2011,673(1):41.
[13] Begg G A,Waldman J R.An holistic approach to fish stock identification[J].Fisheries Research,1999,43(1-3):35-44.
[14] Pinheiro A,Teixeir C M,Rego A L.Genetic and morpholcal variation of Solca lascaris(Risso,1810)along the Portuguese coast[J].Fisheries Research,2005,73(1-2):67-78.
[15] 李冰,柴学森,张成锋,等.建鲤和黑龙江野鲤自交和正反交F1形态差异分析[J].中国水产科学,2014,21(1):59-66.
[16] 康伟,吴廉,刘金生,等.图们江水系绒螯蟹的形态差异与遗传混杂[J].中国水产科学,2016,23(3):555-564.
[17] 张萌,白俊,金辉,等.不同地理群体的克氏原螯虾形态差异多元分析[J].南昌大学学报(理科版),2016,40(2):188-196.
[18] 王志铮,祝本强,任夙艺,等.日本囊对虾(Marsupenaeus japonicus)秋繁同生群仔虾不同耐干露性能群体间的形态表型差异[J].海洋与湖沼,2016,47(3):673-680.
[19] 郑友,胡火根,唐建清,等.不同水域克氏原螯虾群体的形态差异分析[J].南昌大学学报(理科版),2014,38(1):96-102.
[20] 农莹,杨明柳,邢永泽,等.红树蚬与歪红树蚬形态差异分析[J].广西科学院学报,2015,31(4):268-272.
[21] 李朝霞,李健,王清印,等.中国对虾“黄海1号”选育群体与野生群体的形态特征比较[J].中国水产科学,2006,13(3):384-388.
[22] 郭惠,陈立侨,杨国梁,等.不同罗氏沼虾种群形态差异的比较研究[J].中国水产科学,2006,13(4):531-535.
[23] 赵晓勤,倪娟,陈立侨,等.日本沼虾4种群的形态差异分析[J].中国水产科学,2006,13(2):224-229.
[24] 孙苗苗,黄建华,杨其彬,等.斑节对虾4 个地理种群自交与杂交F1的形态特征分析[J].水产学报,2012,36(10):1512-1519.
[25] Hurlbut T,Clay D.Morphometric and meristic differences between shallow and deep-water populations of white hake(Urophycis tenuis)in the southern Gulf of St Lawrence.Canadian Journal of Fisheries and Aquatic Sciences,1998,55,2274-2282.
[26] Chryssa A,Roman L,Ioannis D L.Biometric analysis of lacustrine and riverine populations of Palemonetes antennarius(H.Milne-Edwards,1837)(Crustacea,Decapoda,Palaemonidae)from north-western Greece.Limnologica,2009,39:244-254.
[27] 张尧庭,方开秦.多元统计分析引论[M].北京:科学出版社,1982:393- 401.
[28] Veasey E A,Schammass E A,Vencovsky R,et al.Germplasm characterization of Sesbania accessions based on multivariate analysis[J].Genetic Resources and Crop Evolution,2001,48(1):79-90.
(责任编辑 崔建勋)
Morphological variance analysis of inbred and hybrid offsprings of African strain and“Nanhai No.1”of Penaeus monodon
CAI Yun-chuan1,JIANG Shi-gui2,SUN Miao-miao3,HUANG Jian-hua2,YANG Qi-bin2,JIANG Song2,YANG Li-shi2,QIU Li-hua2,ZHOU Fa-lin2
(1.Guangdong Ocean and Fishery Technology Extension Center,Guangzhou 510220,China;
2.Key Lab.of South China Sea Fishery Resources Exploitation & Utilization,Ministry of Agriculture/ South China Sea Fisheries Research Institute,Chinese Academy of Fishery Sciences,Guangzhou 510300,China;
3.Lianyungang Marine and Fishery Sciences Research Institution,Lianyungang 222044,China)
Three muhivariation analysis methods(principal component analysis,discriminant analysis and cluster analysis)were used to investigate the morphological variations among two inbreds(NN and FF)and two corresponding hybrids(NF and FN)in Penaeus monodon from two populations of African strain and “Nanhai No.1” based on seven morphological indexes.In the principal component analysis,three principal components were constructed,and the cumulative contributory ratio was 68.35%.The discriminant analysis indicated that the discriminant accuracy of inbreds populations was higher than that of hybrid populations.The cluster analysis revealed that two major groups were observed for the four populations.The first group was NN,and the second group was FF,NF and FN.
Penaeus monodon;selfing;hybridize;morphological characteristics;multivariation analysis
S911;S968.22
A
1004-874X(2016)12-0102-06
10.16768/j.issn.1004-874X.2016.12.018
2016-09-22
国家现代农业产业技术体系建设专项(CARS-47);广东省科技计划项目(2014B020202003);广东省海洋渔业科技与产业发展专项(A201501A06,B201500B05);深圳市生物产业发展专项资金现代农业生物产业推广扶持计划项目(NYSW201400331010053t)
蔡云川(1977-),男,硕士,高级工程师,E-mail:caiyc77@163.com
周发林(1975-),男,博士,副研究员,E-mail:zhoufalin@aliyun.com
蔡云川,江世贵,孙苗苗,等.斑节对虾南海1号和非洲品系自交、杂交群体的形态差异分析[J].广东农业科学,2016,43(12):102-107.
