广西崇左叉叶苏铁种群结构与分布格局研究
2016-02-08林建勇何应会梁瑞龙
李 娟,林建勇,何应会,蒋 燚,梁瑞龙
(广西林业科学研究院,广西 南宁 530002)
广西崇左叉叶苏铁种群结构与分布格局研究
李 娟,林建勇,何应会,蒋 燚,梁瑞龙
(广西林业科学研究院,广西 南宁 530002)
在调查广西崇左叉叶苏铁种群结构特征的基础上,根据年龄结构图和存活曲线,分析种群动态,应用扩散系数、聚集指数、平均拥挤度、聚块性指数、集群指数等方法研究分布格局。结果表明,叉叶苏铁幼苗个体比例较大(其中Ⅰ、Ⅱ级幼苗占总个体数的59.17%),种群结构呈金字塔型,属于增长型种群,种群结构比较稳定;种群内开花植株少,仅发现5株开花,未见结实个体;叉叶苏铁种群分布格局呈聚集型,这与叉叶苏铁的生物学和生态学特性密切相关,同时还受到生境异质性影响。
叉叶苏铁;种群结构;分布格局;广西
苏铁属(Cycas)植物是现今地球上最古老的植物类群,进化起源于晚石炭纪,至今已有2.8亿年的历史[1],其中我国的苏铁属所有种都是Ⅰ级保护植物[2]。从20世纪80年代起,就有学者对中国的苏铁属植物进行了研究,其中对攀枝花苏铁(Cycas panzhihuaensis)、德保苏铁(Cycas debaoensis)和苏铁(Cycas revoluta)等的研究较多[3-10]。叉叶苏铁(Cycas bifida)通常生长在石灰岩低峰丛石山中下部石灰岩土上,在我国仅零星分布于广西龙州、大新、崇左及云南弥勒的局部地区,分布海拔为150~400 m的石山季雨林林下。据2012年广西壮族自治区《第二次全国重点保护野生植物资源调查》试点调查报告,叉叶苏铁在广西约有1 397株,16个分布点,分布范围狭窄,种群数量较少,长期受到外界因素胁迫干扰。目前只有少数学者对叉叶苏铁的外部形态、分子生物学等方面进行研究[11-12],而关于叉叶苏铁种群结构和分布格局方面,至今未见有研究报道。为此,我们对叉叶苏铁的种群结构和分布格局进行研究,以了解其种群的发展趋势,对于探索叉叶苏铁濒危机制、合理保护和促进种群扩张具有重要的理论和现实意义,同时也为叉叶苏铁群落生态学的进一步研究提供理论依据。
1 材料与方法
1.1 研究区自然概况
广西崇左市江州区位于广西西南部,西南、东南部为十万大山余脉四方岭,北部为西大明山,东、中部为左江河谷台地、丘陵、小平原,最高山峰为西大明山,海拔1 071 m。气候属于南亚热带季风气候,受东南季风和西南季风的影响,形成较明显的干湿交替季节,年平均气温21.0~22.3℃,年平均降雨量1 150 mm。
研究地点位于江州区太平镇宜村念金上屯的风水林,地理位置为107°17′19″E、22°30′01″N,面积约10 hm2,分布叉叶苏铁约220株(大部分生长良好),海拔160~230 m,地貌为喀斯特地貌发育后期的残存低丘,坡度5°~15°,基岩裸露率为52%,土壤为偏碱性的棕色石灰土;植被类型为以仪花为优势种的石山季节性雨林,林分郁闭度0.7~0.8。伴生植物主要有黄牛木(Cratoxylum cochinchinense)、龙眼(Dimocarpus longan)、灰毛浆果楝(Cipadessa baccifera)、乌墨(Syzygium cumini)、皂荚(Gleditsia sinensis)、盐肤木(Rhus chinensis)、枫香(Liquidambar formosana)等。
1.2 研究方法
采用样方调查方法进行野外调查。通过野外踏查,根据本区域内植被的类型和分布特点,在林分中选择典型位置共设置3个样地,样地面积2 000~4 000 m2,调查总面积为8 400 m2。将3个样地划分为84个10 m×10 m样方,用GPS测定坐标,记录叉叶苏铁的生境条件,包括海拔、坡度、坡向、坡位、枯落物厚度、岩石裸露率等;测量样方内每株叉叶苏铁的自然高、最大叶片长、叶片数量,记录每株生长情况和性别。
种群分布格局采用方差均值法分析,用样方数据进行方差均值(v/m,亦称扩散系数C)的t检验,计算集群度指数(CI) 、平均拥挤度(m*) 、聚块性指数(m*/m) 、集群指数(1/ K)、负二项指数(K),分析种群分布格局类型[13-14,20]。
数据采用Excel 2007软件整理、绘制图表。
2 结果与分析
2.1 叉叶苏铁种群年龄结构
种群年龄结构能够反映林木发展趋势。通常,林木的年龄可以根据胸径和立木级来确定,但是苏铁类植物生长极为缓慢,且有些苏铁种类主干不明显,其年龄大小难以判断。有学者研究了攀枝花苏铁、仙湖苏铁等的年龄判断方法,例如,攀枝花苏铁可以根据叶痕和茎的解剖结构来确定年龄[3],汪殿蓓等[15]通过残存的营养叶叶基数、开花痕数以及生长单元发生率等特征来判断仙湖苏铁种群年龄。但是在野外极少能看到叉叶苏铁地上茎,大部分植株要扒开表土层3~4 cm,才能看到鳞片和茎;而每年新发的叶片数量不等,大部分是1~2片,叶痕呈不明显环状,自然高度容易受到叶片下垂等因素的影响,且叉叶苏铁属于国家Ⅰ级保护植物,不能用大量的材料进行解剖分析。因此,本研究只能根据其最大叶片长度结合实地调查的开花结实情况来绘制其年龄结构图。叉叶苏铁按最大叶片长度可划分成以下10级:Ⅰ级:20~49 cm;Ⅱ级:49~78 cm;Ⅲ级:78~107 cm;Ⅳ级:107~136 cm;Ⅴ级:136~165 cm;Ⅵ级:165~194 cm;Ⅶ级:194~223 cm;Ⅷ级:223~252 cm;Ⅸ级:252~281 cm;Ⅹ级:281~310 cm。
由图1可知,叉叶苏铁的种群结构呈下宽上窄的金字塔型,种群个体大小(株高)基本在Ⅵ级以下,幼苗储备丰富,占个体总数的87.5%,其中Ⅰ级苗数量最多(43株),表现出较强的更新能力,呈稳定增长趋势。可见叉叶苏铁种群结构较为稳定,各阶段都有一定数量的个体,且生长良好,种群数量能维持在稳定水平,但是成年植株数量明显偏少,成为限制种群扩大的主要因素之一。
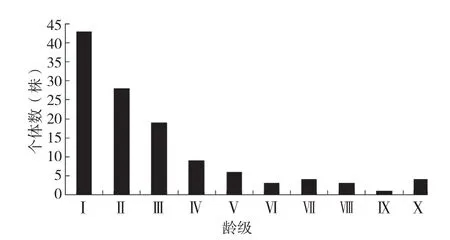
图1 叉叶苏铁种群年龄结构
2.2 存活曲线
用空间替代时间对种群进行研究时,调查数据并非完全符合数学假设样本,会存在前一个龄级的存活个体数小于后一个龄级的情况,但是仍具有生态学意义[16-18]。通过匀滑方法对各年龄级存活个体数进行修正[16],以龄级作横坐标,以存活个体数的对数值(以10为底)作纵坐标,绘制叉叶苏铁种群存活曲线,结果(图2)表明,Ⅰ~Ⅴ级之间存活曲线呈直线下降趋势,该阶段个体死亡率较高,说明幼苗能否长成大苗是影响叉叶苏铁种群稳定的关键;第Ⅴ~Ⅹ级之间的存活曲线下滑趋势有所减缓。
Deevey[17]把存活曲线分成3种类型:Ⅰ型(a型)存活曲线呈凸型,Ⅱ型(b型)存活曲线呈对角线型,Ⅲ型(c型)存活曲线呈凹型。从图2可以看出,叉叶苏铁种群存活曲线符合Deevey-Ⅱ型(b型),表明各龄级死亡率近似,种群结构相对稳定。
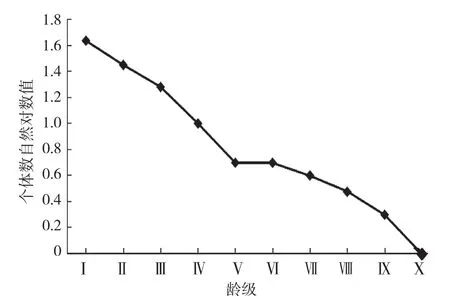
图2 叉叶苏铁种群存活曲线
2.3 开花频率和性别比
苏铁属植物要很长时间才能开花,在华南地区通常需要10年以上,在华北地区则需要60年或更长时间[1]。由于人为采挖的原因,在本次调查区域内叉叶苏铁Ⅵ级以上个体的数量较为稀少,以致在调查到的120株叉叶苏铁中,仅有5株开花,占总株数0.42%,其中2株雌株,3株雄株。在开花植株中,1株为Ⅶ级,1株为Ⅷ级,3株为Ⅹ级。表明在自然条件下,叉叶苏铁Ⅴ级(可能包括Ⅵ级)及以下级的个体不开花,为幼年植株。两次调查均未发现叉叶苏铁结实个体,在开花植株周围未见种子。
2.4 空间分布格局
种群空间分布格局是指组成种群的个体在其生活空间的位置状态或布局[13-14]。将各样方调查数据用方差均值法进行种群分布格局分析,结果(表1)表明,叉叶苏铁在本调查区域为集群分布,其中Q1样地的平均拥挤度(m*)、聚块性指数(m*/m)和聚集指数(1/ k)均明显高于其他两块样地,这说明与样地内叉叶苏铁的数量较多有关。

表1 不同样地叉叶苏铁种群分布格局
叉叶苏铁分布格局受种子传播和环境异质性限制[13,18]。叉叶苏铁以种子或萌蘖的方式来延续其种群[1,14],而其种子主要依靠重力和啮齿动物传播,但重力传播距离较短,多数散布在母树周围;虽然啮齿动物散布种子的距离大于借助重力散布的距离,但啮齿动物对种子的破坏极大,传播效率低。而叉叶苏铁所生长的地方岩石裸露率高,裸露的岩石较大,土层薄,叉叶苏铁的种子只有落入土壤较厚的石缝或没有岩石的区域才能萌发;而且裸露的岩石区域属于本区域植被中的异质性斑块生境,生境异质性导致了叉叶苏铁分布的集群性。在这种生境中叉叶苏铁的根系拓展极其困难,无性繁殖只能限于局部区域,从而进一步加剧了其聚集程度。
3 结论与讨论
3.1 叉叶苏铁种群结构特征
分析种群年龄结构是揭示种群结构现状和更新策略的重要途径之一,是探索种群动态的有效方法[18-20]。本研究运用“空间序列代替时间变化”的方法[13],根据叶片长度划分大小等级,用其代替年龄结构对叉叶苏铁种群的结构特征进行分析,结果表明样地内有大量的Ⅰ、Ⅱ级幼苗,叉叶苏铁种群有较为稳定的结构。但是在幼苗向大苗生长过程中,由于存在较强的环境筛,幼苗难以通过而致大量死亡。环境筛的构件是多方面的,可能是微环境条件的变化,也可能是种内或种间竞争[13-14]。研究环境筛的构建将有助于了解叉叶苏铁濒危的原因。
本研究调查发现,叉叶苏铁成年个体数很少,开花结实的植株更少,仅占总株数的0.42%。这主要是因为近年来当地叉叶苏铁成年植株被人为采挖较多,而且叉叶苏铁通常每隔2~3年才开花结实一次,因此结实个体较少。此外,调查区域的林地周边已被开荒种植农作物,使得叉叶苏铁的生境受到破坏。这些因素导致调查区域内的叉叶苏铁自我繁衍困难,限制了种群增长。但是,目前调查区域内有一定数量的Ⅰ、Ⅱ级幼苗,这些幼苗中有一部分是叉叶苏铁母树通过萌蘖方式产生的,还有一部分为种子实生苗。通过走访得知在2012年该区域的叉叶苏铁结实量较多,因此种子实生苗大部分应该为2014年之前萌发的。这些幼苗的存在表明叉叶苏铁种群能够较好的进行自然更新,如果加强保护,可以提高其成年个体的数量。
从空间分布格局看,叉叶苏铁呈集群分布,造成这种分布格局有其自身繁殖特性、生态适应对策的因素,也有人类干扰、生境恶化的因素。调查样地内的叉叶苏铁个体大多为幼苗,成年个体少,这种集群分布格局在一定程度上能更好地改变小生境(如根系分泌物的影响,或利于其生长发育的微气候),更利于叉叶苏铁幼苗的生长,同时也增强了种群对其他植物侵入等不利因素的抗性,提高其在群落中的竞争力,增加种群对养分和生存空间的占据与巩固[13,18-19]。
3.2 叉叶苏铁种群的保护
叉叶苏铁属于国家I级濒危保护植物,分布范围小,对生境要求较高。在调查过程中发现周围农民砍伐林木,开荒种地,导致叉叶苏铁适生生境面积减少和片段化,其种群破碎成较小单元。目前叉叶苏铁的年龄结构较为稳定,建立保护小区就地保护叉叶苏铁,是有效的保护途径之一,既能保护其现有个体,免遭人为破坏;又能保护其生境,减少人类的干扰和生境占用,使其能在现有生境中生存、繁衍。此外,针对叉叶苏铁繁殖的局限性,可通过采集成熟的种子,选择郁闭度和温湿度适宜、土层较厚的区域进行播种,这是人工扩大叉叶苏铁种群、解除其濒危状况的可行策略。
[1] 王发祥,梁惠波,陈潭清,等.中国苏铁[M].广州:广东科技出版社,1996.
[2] 国家林业局,农业部.国家重点保护野生植物名录(第一批)[J].中华人民共和国国务院公报,2000(13):39.
[3] 何永华,王乾,石培礼.攀枝花苏铁的生物学特性、树干解剖和生长模式[J].植物学报,1995,37(6):443-451.
[4] 何永华,李朝銮.攀枝花苏铁种群生态地理分布、分布格局及采挖历史的研究[J].植物生态学报,1999,23(1):23-30.
[5] 熊亚,李敏杰.攀枝花苏铁种植园土壤微生物数量及土壤酶活性分析[J].广东农业科学,2015,42(22):52-56.
[6] 朱利英,赵春章,莫旭,等.计划烧除攀枝花苏铁林区地面覆盖物对苏铁生长和土壤理化性质的影响[J].应用与环境生物学报,2012,18(3):381-390.
[7] 周明昆,李正文,李志刚,等.模拟酸雨对德保苏铁叶片光合作用及根系分泌有机酸的影响[J].南方农业学报,2012,43(5):587-591.
[8] 骆文华,唐文秀,黄仕训,等.珍稀濒危植物德保苏铁迁地保护研究[J].浙江农林大学学报,2014,31(5):812-816.
[9] 李正文,陈丽丽,李志刚,等.德保苏铁回归后几个生理指标的比较研究[J].广西植物,2012,32(2):243-247.
[10] 田美华,赵正武,唐安军.苏铁种子的脱水敏感性及温湿条件对种子后熟的影响[J].植物生理学报,2016,52(2):225-233.
[11] 王跃华.叉叶苏铁的遗传分化和谱系地理研究[D].昆明:云南大学,2015.
[12] 莫鹏巧,黄玉源,钟晓青,等.利用正交设计法对叉叶苏铁ISSR-PCR反应体系的优化研究[J].植物研究,2008,28(3):304-309.
[13] 茹文明,张桂萍,毕润成,等.濒危植物脱皮榆种群结构与分布格局研究[J].应用与环境生物学报,2007,13(1):14-17.
[14] 张志录,刘中华,陈明辉,等.河南伏牛山区天然红豆杉种群结构与动态研究[J].水土保持研究,2016,23(3):262-268.
[15] 汪殿蓓,暨淑仪,陈飞鹏,等.仙湖苏铁种群年龄判断及年龄结构特征[J].应用生态学报,2007,18(3):476-480.
[16] 江洪.云杉种群生态学[M].北京:中国林业出版社,1992.
[17] Deevey E.Life table for natural populations of animals[J].Quart Rev Biol,1974,22:283-314.
[18] 黄应锋,廖绍波,陈勇,等.深圳梅林仙湖苏铁的种群特征与保护研究[J].林业科学研究,2013,26(5):668-672.
[19] 田波,龚洵,张启泰,等.潭清苏铁的生境、种群结构与动态的研究[J].西北植物学报,2005,25(1):133-137.
[20] Lin Y C,Chang L w,Yang K C,et a1.Point patterns of tree distribution determined by habitat heterogeneity and dispersal limitation[J].Oeeologia,2011,165(1):175-184.
(责任编辑 邹移光)
Population structure and distribution pattern of Cycas bifida in Chongzuo of Guangxi
LI Juan,LIN Jian-yong,HE Ying-hui,JIANG Yi,LIANG Rui-long
(Guangxi Forestry Research Institute,Nanning 530002,China)
The population of Cycas bifida was investigated in Chongzuo,Guangxi.The population structure of C.bifida was analyzed by age structure and survivorship curve.The pattern of C.bifida was studied by using dispersal index,clump intensity,mean crowding,patchiness index,cassie index,etc.The results showed that the population dynamics belonged to growth type because there were lots of seedlings,and the population structure was relatively stable.It was found that the blooming plant was very few,only 5 in the population.There was no seeding plant.Spatial distribution pattern of C.bifida population conformed to clumped distribution,which closely related to its ecological and biological characteristics,as well as affected by its habitat heterogeneities.
Cycas bifida;population structure;distribution pattern;Guangxi
S791.11
A
1004-874X(2016)12-0025-05
10.16768/j.issn.1004-874X.2016.12.005
2016-09-21
国家林业局部门预算广西项目支出预算项目(桂林护预2014005)
李娟(1984-),女,工程师,E-mail:lijuan1434@163.com
李娟,林建勇,何应会,等.广西崇左叉叶苏铁种群结构与分布格局研究[J].广东农业科学,2016,43(12):25-29.
