青藏高原东缘2种优势灌木的光合特性研究
2016-02-06高丽楠成都大学旅游与经济管理学院四川成都6006四川师范大学地理与资源科学学院四川成都60066
高丽楠,张 宏,肖 艳(.成都大学 旅游与经济管理学院,四川 成都 6006; .四川师范大学 地理与资源科学学院,四川 成都 60066)
青藏高原东缘2种优势灌木的光合特性研究
高丽楠1,张 宏2,肖 艳1
(1.成都大学 旅游与经济管理学院,四川 成都 610106; 2.四川师范大学 地理与资源科学学院,四川 成都 610066)
在夏季7月利用Li-6400便携式光合仪器和PAM-2100调制叶绿素荧光仪,选择川西北高寒灌丛草甸2种优势灌木窄叶鲜卑花(Sibiraeaangustata)和高山绣线菊(Spiraeaalpine)为对象,研究光合作用日变化及其对环境因子的响应。结果表明:高山绣线菊的光合速率(Pn)和蒸腾速率(Tr)日变化趋势均呈“双峰”型,而窄叶鲜卑花的Pn和Tr日变化趋势均呈“单峰”型,都存在光合“午休”现象,窄叶鲜卑花的Pn日均值和Tr日均值都显著大于高山绣线菊。随日间光合有效辐射强度(PAR)的增强,2种灌木的最大光能转化效率(Fv/Fm)下降,19:00均未恢复到黎明时的水平,表明光合机构因逆境而受到不可恢复的损伤。通径分析表明,Tr为高山绣线菊和窄叶鲜卑花Pn日变化的主要决定因子,叶温(Tleaf)则是主要限制因子。研究表明,高山绣线菊以较小的叶面积、低Pn、低Tr和较高的水分利用效率(WUE)显示出在受水分胁迫的强光高温环境下的适应性;但是窄叶鲜卑花属于高光合、高蒸腾和低WUE灌木,其半饱和光强(Ek)和光强最弱时光能利用效率(α)都较高,表明对弱光或较强的PAR均能利用,PAR适应范围相对最广。
高寒灌丛草甸; 光合速率; 水分利用效率
青藏高原有世界面积最大的高寒灌丛草甸,面积约有11.64万km2,是青藏高原高寒草地生态系统的重要组成部分之一[1]。高寒灌丛草甸对全球变化更为敏感,其土壤层的碳释放和植被层的碳吸收在区域生态系统碳平衡过程中起着非常重要的作用;然而,植物生长发育的决定性因素是光合作用,同时光合作用所吸收的CO2是陆地生态系统碳循环的入口,并对减缓全球温室效应起着重要作用,光合作用也是一个对环境变化异常敏感的生理过程[2]。因此,当前迫切需要开展高寒灌丛草甸的光合特性研究。
高寒灌丛是由耐寒的中生或旱生灌木为优势种而形成的一类植被,能很好地发挥遮蔽地表、减少蒸发、固持土壤和改善土壤结构作用,是高寒灌丛草甸水源涵养和生态系统稳定的重要植被。研究高寒灌木光合特性的适应机制,对高寒草甸水资源的持续利用以及生态环境建设有着极其重要的现实意义。目前,关于高寒灌丛的生物量[3]、抗旱特性[4]和根系变化特征[5]已有不少报道,而高寒灌丛的光合作用研究极少[6]。
川西北高寒灌丛草甸位于青藏高原东部,是四川重要的水源地,也是黄河上游重要的水源集水区和涵养区,是我国主要牧区之一。目前川西北高寒灌丛草甸在人为因素与自然因素干扰的双重影响下,呈现出明显的退化态势。其中,窄叶鲜卑花和高山绣线菊是该地区特有的、典型的、具有代表性的高寒灌丛植被,对维持青藏高原的生态平衡起着重要作用,目前还未见对其生理生态学方面的专题报告。鉴于此,本研究重点讨论青藏高原东缘高寒灌丛草甸2种灌木生理特性和光照生态适应性,测定了2种灌木的光合及其叶绿素荧光参数的日变化,以探讨高寒环境对植物光合作用的影响及高山植物的光合特性,为高原地区植被恢复与重建提供理论科学依据。
1 材料和方法
1.1 试验样地概况
试验地位于四川省阿坝州红原县(101°51′~103°23′E、31°51′~33°19′N),平均海拔3 600 m,地处青藏高原东部边缘,地势为东南向西北倾斜。大陆性高原气候,无明显四季之分,日温差大、霜冻期长,年平均气温1.1 ℃,7月平均气温是10.8 ℃,1月平均气温是-10.2 ℃,年均相对湿度60%~70%;年降水量792.83 mm,多集中于5—10月;年蒸发量为1 254.3 mm,一年中除6、7、8月外,其余月份降水量均小于蒸发量,空气干燥;太阳辐射强,日照时间长,年均日照2 146.8 h[7]。
采样点位于红原县城北10 km的龙壤乡草场,选取了约200 m2的亚高山灌丛草甸(102°36′E、33°11′N;海拔3 495 m左右)样地,灌丛以高山绣线菊(Spiraeaalpine)和窄叶鲜卑花(Sibiraeaangustata)为优势种,伴生灌木有西藏忍冬(Loniceratibetica)、藏沙棘(Hippophaethibetana)和金露梅(Dasiphorafruticosa),样地灌木分盖度为60%左右,由灌木+草本植物构成复合群落的总盖度在95%以上[7]。
1.2 项目测定方法及数据处理
1.2.1 光合参数日变化的测定 于灌木生长旺季的7月9—18日,采用Li-6400便携式光合仪器(Li-Cor,Lincoln,NE,USA)对2种灌木的光合生理指标进行测定。选择晴朗无云的天气,7:00—19:00,每2 h测定一次。分别随机选取5株生长良好且树龄相近的窄叶鲜卑花和高山绣线菊,并将健康枝条自顶端以下第3~4片健康叶作为测量对象,每株选取2片叶子进行测定,测定指标有光合速率[Pn,μmol/(m2·s)]、蒸腾速率[Tr,mmol/(m2·s)]、大气CO2浓度(Ca,μmol/mol)、光合有效辐射强度[PAR,μmol/(m2·s)]、大气相对湿度(RH,%)、气温(Tair,℃)、胞间CO2浓度(Ci,μmol/mol)、叶片气孔导度[Gs,mmol/(m2·s)]和叶片温度(Tleaf,℃)。最后根据以上参数,计算出叶片水分利用效率(WUE,μmol/mol)(WUE=Pn/Tr)[8]和气孔限制值(Ls)(Ls=1-Ci/Ca)[9]。使用AM300叶面积仪进行叶面积的测定。
1.2.2 叶绿素荧光参数日变化的测定 在自然条件下2种灌木叶绿素荧光参数测定采用PAM-2100(Walz,Germany)便携式调制叶绿素荧光仪,叶片暗适应30 min后测量相关指标。测得光系统Ⅱ(PSⅡ)的最大光能转化效率(Fv/Fm)[Fv/Fm=(Fm-Fo)/Fm],实际光化学反应效率(Yeild)[Yeild= (Fm′-Ft)/Fm′][10]。这里,Fo和Fm分别代表暗适应后的初始荧光和最大荧光,Fm′代表光适应后的最大荧光,Ft代表光下稳态荧光。测定快速光响应曲线由电脑控制,光化光强度设定为70、125、200、320、500、700、1 000、1 750、2 300 μmol/(m2·s),打开测量光和光化光,适应10 s后打开饱和脉冲,之后升高光化光强度并适应10 s,饱和脉冲再打开,如此重复8次。打开饱和脉冲前的荧光为Ft,打开饱和脉冲得到最大荧光为Fm′。根据Yeild和PAR可以计算出相对电子传递速率[rETR,μmol/(m2·s)](rETR=Yeild×PAR×0.5×0.84)[10]。
采用Platt等的方程式对快速光曲线进行拟合[11]:rETR=rETRmax×(1-e-α×PAR/rETRmax)×e-β×PAR/rETRmax,式中,rETRmax为无光抑制时的最大潜在相对电子传递速率,α为快速光曲线的初始斜率,β为光抑制参数。由此可以得出半饱和光强[Ek,μmol/(m2·s)],Ek=rETRmax/α。
所有数据统计与分析均在Originpro 7.5、SPSS 12软件下完成。
2 结果与分析
2.1 环境因子日变化规律
由图1可知,PAR的日变化趋势表现为典型的“单峰”型曲线,13:00左右达到高峰值2 153.36 μmol/(m2·s),之后逐渐下降,PAR日平均值为1 406.24 μmol/(m2·s)。Ca基本保持不变,日均值为379.98 μmol/mol。在7:00左右RH最高(48.68%),其后随着PAR、Tair的增加而逐渐降低,到13:00降至最低值(29.41%),之后有所回升。Tair随着PAR的增强而迅速上升,到15:00左右达到全天最高(28.87 ℃),Tair的最高峰值比PAR落后2 h,之后随着PAR的减弱而降低,日平均气温为 22.83 ℃。
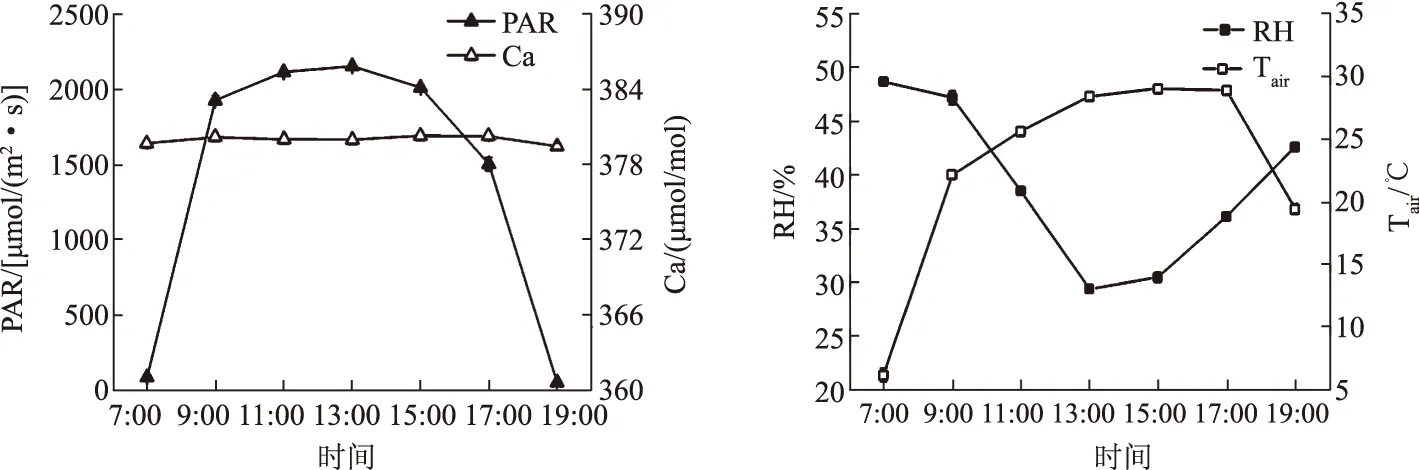
图1 窄叶鲜卑花和高山绣线菊的环境因子日变化
2.2 Pn及其参数日变化特征
2.2.1 Pn和Tr日变化特征 由图2可知,窄叶鲜卑花Pn日变化表现为典型的“单峰”曲线,其最高峰值出现在9:00,为21.51 μmol/(m2·s),此后随着PAR的增强而持续下降。高山绣线菊的Pn日变化趋势为“双峰”型,Pn的2个峰值分别为17.24 μmol/(m2·s)和12.19 μmol/(m2·s),分别出现在9:00和15:00,低谷值出现在13:00,高山绣线菊Pn的第2个峰值低于第1个峰值,表现出明显的光合“午休”现象。另外,窄叶鲜卑花Pn日均值极显著高于高山绣线菊(P<0.01)。
Tr日变化趋势和Pn日变化相似,窄叶鲜卑花Tr日动态表现为“单峰”型曲线,但高山绣线菊为“双峰”型曲线。窄叶鲜卑花Tr最高峰值仅出现在9:00左右,而高山绣线菊Tr的2个峰值分别出现在9:00和15:00,9:00—13:00窄叶鲜卑花Tr的下降速率小于高山绣线菊。窄叶鲜卑花Tr日均值极显著高于高山绣线菊(P<0.01)。

图2 窄叶鲜卑花和高山绣线菊的Pn、Tr日变化
2.2.2 Gs和Ci日变化特征 由图3可见,窄叶鲜卑花Gs日变化趋势表现为“单峰”曲线,从7:00开始随着PAR和Tair的增加,Gs逐渐增加,9:00左右出现峰值,之后因高温低湿的环境引起Gs降低。但高山绣线菊Gs日变化趋势表现为微弱的“双峰”态势,最高值出现在9:00左右,之后呈降低趋势,13:00—15:00逐渐上升。窄叶鲜卑花的Gs日均值极显著高于高山绣线菊(P<0.01)。
高山绣线菊的Ci变化趋势与Pn变化趋势恰好相反,呈近“V”型变化曲线,表现为早晚高、中午低的趋势。这主要是由于随着Pn的增强,消耗的Ci增多,并且随着温度的升高,Gs也逐渐降低,植物吸收外界的CO2减少而形成的。7:00时窄叶鲜卑花的Ci为342.76 μmol/mol,随着Pn的加强,叶片固定较多的CO2致使Ci迅速下降;13:00左右出现低谷,之后呈上升趋势,15:00—17:00又降低,之后再上升,到19:00几乎达到350 μmol/mol。窄叶鲜卑花与高山绣线菊的Ci日平均值无显著性差异(P>0.05)。
2.2.3 Ls及Tleaf日变化特征 由图3可见,从Ls日变化趋势来看,与Ci呈现出相反的趋势,高山绣线菊的Ls日动态呈现出“单峰”型曲线,最高峰值均出现在13:00左右。窄叶鲜卑花的Ls日动态呈现出“双峰”型曲线,2个峰值分别出现在13:00和17:00左右。9:00—11:00窄叶鲜卑花Ls显著高于高山绣线菊(P<0.01)。
Tleaf日变化趋势与Tair日变化相似,最高Tleaf出现在15:00左右,窄叶鲜卑花和高山绣线菊的日均Tleaf分别为24.87 ℃和22.52 ℃。窄叶鲜卑花(r=0.98,P<0.01)和高山绣线菊(r=0.986,P<0.01)的Tleaf都与Tair呈极显著正相关,所以Tleaf主要受Tair影响。

图3 窄叶鲜卑花和高山绣线菊的Gs、Ci、Ls及Tleaf日变化
2.2.4 WUE日变化特征 由图4可见,窄叶鲜卑花WUE日变化趋势呈“双峰”型曲线,7:00—9:00呈迅速上升趋势,之后缓慢降低,13:00—17:00又缓慢上升。高山绣线菊的WUE日变化都呈现出“单峰”型曲线,其WUE在11:00时达到最大,为2.61 μmol/mol,之后逐渐降低。11: 00时,高山绣线菊的WUE极显著高于窄叶鲜卑花(P<0.01),但15:00和17:00恰好相反。另外,高山绣线菊WUE日均值(2.0 μmol/mol)显著高于窄叶鲜卑花(1.7 μmol/mol)(P<0.05)。

图4 窄叶鲜卑花和高山绣线菊的WUE日变化
2.3 Pn与生理生态因子的相关分析及通径分析
2.3.1 窄叶鲜卑花Pn与生理生态因子的关系 为了解生理生态因子对窄叶鲜卑花Pn的影响,对窄叶鲜卑花Pn与生理生态因子进行了相关分析和通径分析,结果(表1和表2)表明:从相关系数的大小看来,各生理生态因子对Pn影响的大小顺序为Tr>PAR>Gs>Ci>Tleaf>Tair>RH>Ca。其中Tr、PAR、Gs、Tleaf、Tair、RH与Pn呈极显著正相关,而Ci与Pn呈极显著的负相关。各生理因子对Pn直接通径系数的大小依次为Tair>Tr>Gs>PAR>Ca>RH>Ci>Tleaf;Tleaf负向的直接作用小于其对PAR、Tair、Gs、Ca、Ci和Tr的正向间接作用,故表现为其与Pn呈极显著的正相关性。生理生态因子的决策系数顺序为R2(Tr)>R2(Ci)>R2(Tair)>R2(Gs)>R2(PAR)>R2(Ca)>R2(RH)>R2(Tleaf),其中的R2(RH)和R2(Tleaf)小于0。所以,影响窄叶鲜卑花光Pn日变化规律的决定生理生态因子为Tr、Gs、PAR、Ca、Tair和Ci,其中Tr为主要决定因子;Tleaf为主要限制因子。

表1 窄叶鲜卑花的Pn与生理生态因子的通径系数
表2 Pn与生理生态因子的相关系数
注:*、**表示分别在 0.05、0.01水平上显著、极显著相关。
2.3.2 高山绣线菊Pn与生理生态因子的关系 由表2和表3看出,高山绣线菊Pn与生理因子相关分析和通径分析结果表明:PAR、Ca、RH、Tair、Gs、Tr、Tleaf与Pn呈极显著正相关,而Ci与Pn呈显著的负相关。各生理生态因子对高山绣线菊的Pn直接通径系数的大小为Tr>RH>Tair>Ca>PAR>Gs>Tleaf>Ci;尽管RH通过PAR、Ca、Gs和Tleaf对Pn的间接作用均为负值,但RH直接作用(0.591)远高于这些间接作用,所以没有影响RH与Pn呈极显著正相关; Gs对Pn的直接作用不大(-0.363),但被Tr正间接作用(0.574)所掩盖而使其与Pn正相关;Tr正向的直接作用远大于其对PAR、Gs、Ca和Tleaf的负向间接作用,故表现为其与Pn呈显著的正相关性。生理因子的决策系数顺序为R2(Tr)>R2(RH)>R2(Ci)>R2(Tair)>R2(Ca)>R2(Gs)>R2(PAR)>R2(Tleaf),其中的R2(PAR)、R2(Ca)、R2(Gs)和R2(Tleaf)均小于0。所以,影响高山绣线菊Pn日变化规律的决定生理生态因子为RH、Tair、Ci和Tr,其中Tr为主要决定因子;限制因子为PAR、Ca、Gs和Tleaf,其中Tleaf为主要限制因子。

表3 高山绣线菊的Pn与生理生态因子的通径系数
2.4 叶绿素荧光参数日变化特征
2.4.1 Fv/Fm和Yeild 2种灌木的Fv/Fm和Yeild的日变化趋势相似(图5),总体呈现出先降低后升高的趋势,一天中7:00最高,上午随着PAR上升而逐渐降低,13:00降为最低值,下午随着PAR的降低又逐渐回升,19:00时2种灌木Fv/Fm并未完全恢复到7:00的水平。窄叶鲜卑花13:00 Fv/Fm相对于其7:00时降低了75.3%,而高山绣线菊降低了72.2%。7:00时,2种灌木Fv/Fm之间存在显著差异(P<0.05),窄叶鲜卑花的日均Fv/Fm和Yeild均高于高山绣线菊。

图5 窄叶鲜卑花和高山绣线菊的Fv/Fm及Yeild日变化
2.4.2 快速光响应曲线 窄叶鲜卑花的rETRmax日变化趋势呈“单峰”曲线,在9: 00时达到峰值,之后rETRmax逐渐降低。高山绣线菊rETRmax日变化趋势表现为“双峰”型,第1峰出现在9:00左右,之后降低,低谷出现在13:00左右,之后略有上升,第2峰出现在15:00左右。另外,窄叶鲜卑花和高山绣线菊的Ek日变化趋势都呈“单峰”曲线,均在9:00达到峰值。窄叶鲜卑花的rETRmax和Ek日平均值均极显著高于高山绣线菊(P<0.01)。
高山绣线菊的α一天中呈早晚高、中午低的近“V”型变化趋势,13:00时最低,之后逐渐回升。另外,窄叶鲜卑花α日变化,7:00—9:00时逐渐降低,9:00—13:00时波动较小,之后继续降低,15:00左右出现低谷,之后再上升。窄叶鲜卑花α分别在7:00、13:00和19:00时显著高于高山绣线菊(P<0.01)。

图6 窄叶鲜卑花和高山绣线菊的rETRmax、α及Ek日变化
3 结论与讨论
3.1 窄叶鲜卑花与高山绣线菊的光合特性
植物叶片对光能的吸收、传递与利用能力不仅与环境因子有关,也与自身生理状态密切相关。2种灌木Pn日变化曲线不尽相同,高山绣线菊Pn日变化为“双峰”型曲线,而窄叶鲜卑花Pn日变化为单峰曲线,表明2种灌木对外界不断变化的环境因子采取了不同的响应策略。本试验中高山绣线菊Pn和Tr的日变化趋势均表现为“双峰”型。清晨随着PAR、Tair、Gs逐渐增加,PAR对光合作用仍具有正效应,9:00左右Pn和Tr达到峰值;之后PAR使叶片蒸腾失水过多,当在水分供应不充足时通过Gs的减小来降低水分的过度消耗,与外界气体交换受阻,使Ci大幅度降低,进入叶肉细胞内用于光合作用的CO2供应不足,致使Pn下降。在午后,由于温度的降低,PAR减弱,使Gs有所回升,主要依靠其发达的根系组织,扎根深或向水平方向扩展[12-13],可充分吸收土壤深层水分来满足生长需要。但是,窄叶鲜卑花Pn、Tr和Gs的日变化趋势呈“单峰”型,清晨PAR逐渐增强,窄叶鲜卑花Pn迅速提高,Tr加快,在9: 00达到峰值;9:00以后随着PAR的增强和Tair升高,植物蒸腾和土壤蒸发量逐渐增大[14],使植物体剧烈失水,对其造成局部高温胁迫和叶片水分胁迫。而窄叶鲜卑花是浅根类型[15],主要利用土壤浅层水分,植物根系得不到充足的水分供应,使叶片的Gs减少蒸腾失水[16],Pn也随之下降处于较低水平,所以窄叶鲜卑花Pn、Tr和Gs的日变化趋势呈“单峰”型,这与青海省的乌柳(Salixcheilophila)研究结果相一致[17]。
窄叶鲜卑花和高山绣线菊Pn的日变化都呈现出光合“午休”现象,这与青藏高原东部裂叶沙参(Adenophoralobophylla)[18]的研究结果相一致。引起植物叶片Pn降低的自身因素主要有2类:一是气孔限制由气孔部分关闭引起,二是非气孔限制由叶肉细胞光合活性下降引起。高山绣线菊在9:00—13:00 Pn下降分别对应Ls上升和Ci下降,所以可以推断出引起光合“午休”的主要原因是气孔因素;午后,PAR减弱,15:00—19:00阶段,Pn的下降伴随着Ci上升,Ls下降,说明非气孔因素是其Pn下降的决定因素之一。而窄叶鲜卑花在9:00—13:00时段,Pn与Ci均下降,Ls上升,说明Pn下降主要是气孔因素造成;13:00—15:00时段和17:00—19:00时段,Pn下降同时伴随着Ci上升,Ls下降,表明非气孔因素是其Pn下降的决定因素之一。有研究表明,植物水分亏缺时Pn降低并不是由于水分供应不足直接引起的,轻度水分胁迫通过气孔因素限制光合作用的进行,而重度水分胁迫下光合作用的降低主要是由非气孔因素造成的[19]。受到轻度水分胁迫的高山绣线菊,较低的Tr和较高的WUE显示出其在受水分胁迫的强光高温环境下的适应性,所以,午后由于PAR减弱,Pn在15:00出现次高峰,日变化趋势呈“双峰”型。然而,受到较重水分胁迫的窄叶鲜卑花叶片光合午休表现得更严重,其Pn日变化类型表现为“一降不起型”。午间强PAR将导致高温,地表蒸发和植物蒸腾加剧以及空气湿度显著下降,水分亏缺引起Gs降低,Pn也随之下降,因为窄叶鲜卑花是浅根系植物,之后难以恢复到较高水平,日变化趋势呈“单峰”型。有研究也表明,适度水分胁迫时红砂Pn的日变化表现出“双峰”型,但当土壤含水量降低产生严重水分胁迫时,Pn的日变化表现出“单峰”型[20]。所以,水分供应状况对2种植物的光合作用非常重要,植物通过蒸腾作用调节叶面温度、供应光合作用所需水分等,与Pn关系密切[21]。本研究结果也表明,2种灌木植物光合作用Pn的主要决定因子是Tr,表明2种灌木的Pn在气孔严格调控的基础之上,与叶片Tr存在紧密联系,Pn的变化是植物自身生理和环境生态因子综合作用的复杂过程。这与对克里雅河流域的胡杨(Populuseuphratica)和柽柳(Tamarixramosissima)的研究结果一致,Tr是2种灌木Pn的主要影响因子[22]。
对于高寒灌丛草甸来说,上午植物水分状况较好,Tr能反映植物的蒸腾能力、气孔传导能力;午后,因上午植物的水分吸收小于强烈蒸腾,引起水分亏缺,促使Tr下降,此时高原植物的Tr主要取决于植物的水分状况。午前,窄叶鲜卑花Tr大表明其蒸腾能力、气孔传导能力强,午后高山绣线菊Tr上升可能是由于保水能力强,导致水分状态好的结果。从蒸腾作用来看,高山绣线菊节水能力优于窄叶鲜卑花。另外,高山绣线菊日均WUE较高,说明生产相同量的物质消耗水分数量较少。因为,高山绣线菊具有发达的根系组织,可以充分吸收土壤水分补充蒸腾失水来满足生长需要;而且其叶面积显著小于窄叶鲜卑花,减少蒸腾和受光面积,从叶片水平上来讲,高山绣线菊节水能力也优于窄叶鲜卑花。因高山绣线菊叶面积小,不利于与外界环境进行气体交换,在降低水分散失的同时也影响了CO2吸收速度,降低了自身的Pn,具有低光合、低蒸腾、高WUE的特点。2种灌木之间WUE日变化特点也不同,在早晨温、光、空气湿度等条件都较好的情况下,高山绣线菊以高光合低蒸腾的方式来提高WUE;到下午当一切环境条件均处于不利情况下,适应环境变化的高山绣线菊,出现高蒸腾低光合的现象。这表明在不利环境出现之前高山绣线菊具有在短暂时间内充分利用水与光资源的能力,在其他荒漠植物上这一特点也得到了验证[23]。而9:00—13:00窄叶鲜卑花Tr的下降速率小于高山绣线菊,窄叶鲜卑花要保持较高的Tr,不但没提高其WUE,相反往往会通过很高的Tr来抵御高温对其叶片伤害。
3.2 窄叶鲜卑花与高山绣线菊的叶绿素荧光特性
正常生理状态下Fv/Fm一般在0.80~0.85,其大小代表了植物潜在最大光合能力[24],且不受生长条件和物种的影响,当Fv/Fm降低时,表明植物受到了胁迫。凌晨的Fv/Fm理论上应该是最大的,因太阳光尚未直射到植物体上,植物体经过一整夜的暗适应,所有电子门均处于开放状态。但本研究结果表明,窄叶鲜卑花和高山绣线菊的Fv/Fm最大值出现在7:00,分别为0.76和0.69,均低于0.80。这可能是由其独特的环境条件所造成的,当地属高寒灌丛草甸,即使在7月,早晨7:00平均气温也只有6.2 ℃左右,植物遭受低温胁迫,参与光反应电子传递的各种光合色素及蛋白质并没有完全被活化,PSⅡ反应中心活性显著降低,导致PSⅡ传递电子的能力降低,所以出现早晨Fv/Fm值较正常值偏低的现象[25]。而且,7:00窄叶鲜卑花Fv/Fm显著高于高山绣线菊,表明窄叶鲜卑花具有较高的光能转化效率,有利于其适应气温的急剧变化和维持较高的能量消耗,因此窄叶鲜卑花的rETRmax较高。本研究结果表明,2种灌木植物的Fv/Fm随PAR增强呈不断降低趋势,表明这2种灌木遭受光抑制,PAR越强光抑制现象越严重[26]。13:00窄叶鲜卑花Fv/Fm相对于其7:00降低了75.3%,与高山绣线菊相比光抑制程度更大,窄叶鲜卑花遭受到较重水分胁迫,从而导致其Pn迅速降低。另外,19:00时2种灌木Fv/Fm有恢复但没有达到7:00的水平,表明PSⅡ反应中心可能发生了不可逆失活,光合机构因逆境而受到不可恢复的损伤。这可能与青藏高原极端环境有关,高原植物遭受较严重的光抑制是由低温、强PAR和低气压等胁迫因子造成[27]。进一步说,在遭受低温胁迫的情况下,高原植物的光抑制恢复都较慢[28]。通径分析结果也表明,Tleaf则是2种灌木植物Pn主要的限制因子。
通过快速光响应曲线不仅能判断当前状态下植物的光合作用能力,而且能够评估环境中各种光强对植物潜在光合作用能力的影响。对于窄叶鲜卑花,其rETRmax和Ek均显著高于高山绣线菊,这说明,在强光生境下的窄叶鲜卑花具有较高电子传递速率、耐光抑制能力[29]。另外,一天中,2种灌木植物有12 h处在光饱和状态,这是因为7:00—19:00 Ek均低于PAR。本研究表明,2种灌木植物α日变化趋势表现为早晚高、中午低。这是因为,中午PAR较强,植物可以通过降低捕光能力α,从而避免吸收太多的光能造成损伤,说明2种灌木有高效利用强光和调控光合作用的能力。7:00、13:00和19:00,在PAR最弱和最强时,窄叶鲜卑花α高于高山绣线菊,表明窄叶鲜卑花对弱光和强光的利用能力都较强。
川西北高寒灌丛草甸2种灌木的光合生理特征之间有着较大的差异,说明不同植物对同一环境的适应能力存在很大差别。高山绣线菊Pn和Tr的日变化趋势呈“双峰”型,但是窄叶鲜卑花则呈“单峰”型。高山绣线菊是低光合、低蒸腾和较高WUE的灌木,具有发达的根茎和较小的叶面积,显示出在受水分胁迫的强光高温环境下的适应性;但是窄叶鲜卑花属于高光合、高蒸腾和低WUE灌木,其Ek、Pn和光强最弱时α都较高,表明其对弱光或较强的PAR均能利用,PAR适应范围相对最广。Tr是2种灌木Pn的主要决定因子,即高寒灌丛草甸2种灌木光合作用对高原环境的适应取决于水分状况。在生长季节通过植物蒸腾散失过多的土壤水分,导致土壤含水量进一步减少,从而加剧高寒灌丛草甸的退化。所以要保护和恢复高寒灌丛草甸资源,合理利用高寒灌丛草甸的植物资源,以及在已经退化的高寒灌丛草甸中应尽可能选择低Tr、高WUE的土著灌木物种,避免水分消耗过多而影响当地水资源供需平衡,这些都是在现有条件下保护和恢复高寒灌丛草甸的策略。
[1] 王启基,李世雄,王文颖,等.江河源区高山嵩草(Kobresiapygmaea)草甸植物和土壤碳、氮储量对覆被变化的响应[J].生态学报,2008,28(3):885-893.
[2] 张新时,周广胜,高琼,等.中国全球变化与陆地生态系统关系研究[J].地学前缘,1997,4(1/2):137-144.
[3] 邱权,李吉跃,王军辉,等.干旱胁迫下青藏高原4种灌木生物量和根系变化特征及抗旱[J].西北林学院学报,2013,28(3):1-6.
[4] 邱权,潘昕,李吉跃,等.青藏高原20 种灌木生长时期根系特征及抗旱性初探[J].中南林业科技大学学报,2014,34(3):29-37.
[5] 卢鑫,张丽静,王瑞,等.玛曲高寒沙化草地3种灌木根际土壤磷素含量特征[J].草业科学,2012,29(2):167-173.
[6] 潘昕,李吉跃,王军辉,等.干旱胁迫对青藏高原4种灌木生理指标的影响[J].林业科学研究,2013,26(3):352-358.
[7] 张伟,张宏.青藏高原东缘红原地区三种不同草甸土壤活性碳特征[J].山地学报,2008,26(2):205-211.
[8] 张正斌,山仑.作物水分利用效率和蒸发蒸腾估算模型的研究进展[J].干旱地区农业研究,1997,15(1):73-78.
[9] 许大全.光合作用效率[M].上海:上海科学技术出版社,2002:86-89.
[10] Schreiber U,Gademann R,Ralph P,etal.Assessment of photosynthetic performance ofProchloroninLissoclinumpatellain hospite by chlorophyll fluorescence measurements[J].Plant and Cell Physiology,1997,38(8):945-951.
[11] Platt T,Gallegos C L,Harrison W G.Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton[J].Journal of Marine Research,1980,38(4):687-701.
[12] 邱权,潘昕,李吉跃,等.青藏高原20种灌木抗旱形态和生理特征[J].植物生态学报,2014,38(6):562-575.
[13] 舒乃醒.高山绣线菊育苗和造林技术探讨[J].园艺与种苗,2013(5):41-43.
[14] 李英年,赵亮,徐世晓,等.祁连山海北高寒湿地植物群落结构及生态特征[J].冰川冻土,2006,28(1):76-84.
[15] 吴宁.川西北窄叶鲜卑花灌丛的类型和生物量及其与环境因子的关系[J].植物学报,1998,40(9):860-870.
[16] Zhang J W,Davies J.Increased synthesis of ABA in partially dehydrated root tips and ABA transport from roots to leaves[J].Journal of Experimental Botany,1987,38(12):2015-2023.
[17] 刘海涛,贾志清,朱雅娟,等.林龄对高寒沙地乌柳光合特性的影响[J].东北林业大学学报,2012,40(12):20-26.
[18] 祖元刚,阎秀峰,张文辉,等.青藏高原东部不同海拔高度裂叶沙参气体交换与水分利用效率[J].植物学报,1998,40(10):947-954.
[19] 高丽,杨劼,刘瑞香.不同土壤水分条件下中国沙棘雌雄株光合作用、蒸腾作用及水分利用效率特征[J].生态学报,2009,29(11):6025-6034.
[20] 刘玉冰,张腾国,李新荣,等.红砂忍耐极度干旱的保护机制:叶片脱落和茎中蔗糖累积[J].中国科学C辑,2006,36(4):328-333.
[21] Sparks J P,Black R A.Regulation of water loss in populations ofPopulustrichocarpa:The role of stomatal control in preventing xylem cavitation[J].Tree Physiol,1999,19(7):453-459.
[22] 丁俊祥,邹杰,唐立松,等.克里雅河流域荒漠-绿洲交错带3种不同生活型植物的光合特性[J].生态学报,2015,35(3):733-741.
[23] 蒋高明,朱桂杰.高温强光环境条件下3种沙地灌木的光合生理特点[J].植物生态学报,2001,25(5):525-531.
[24] Genty B,Briantais J M,Baker N R.The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence[J].Biochimica et BiophysicaActa(BBA):General Subjects,1989,990(1):87-92.
[25] Huner N P A,Oquist G,Hurry V M.Photosynthesis,photoinhibition and low temperature acclimation in cold to-lerant plants[J].Photosynth Res,1993,37(1):19-39.
[26] 李翊华,张芬琴,陈修斌,等.温室水肥耦合对甜椒生长和果期叶片光合特性的影响[J].江苏农业学报,2015,31(2):415-421.
[27] 郭连旺,沈允钢,许大全,等.几种高寒草甸常见植物的光合特性及其光合作用的光抑制[C]//周兴民,周立,王启基,等.高寒草甸生态系统(第4集).北京:科学出版社,1995:65-74.
[28] Franklin L A.The effects of temperature acclimation on the photoinhibitory responses ofUlvarotundataBlid [J].Planta,1994,192(3):324-331.
[29] Ralph P J,Gademann R,Dennison W C.In situ seagrass photosynthesis measured using a submersible,pulse-amplitude modulated fluorometer[J].Marine Biology,1998,132(3):367-373.
Study on Photosynthetic Characters of Two Dominant Shrub Species in Eastern Tibet Plateau
GAO Linan1,ZHANG Hong2,XIAO Yan1
(1.College of Tourism and Economic Management,Chengdu University,Chengdu 610106,China; 2.College of Geography and Resource Science,Sichuan Normal University,Chengdu 610066,China)
The LI-6400 portable photosynthesis analysis system and PAM-2100 chlorophyll fluorometer were used to study the diurnal variation of photosynthesis of two dominant shrub speciesSibiraeaangustataandSpiraeaalpinein alpine shrub meadow and the response to environment factors in July.The results showed that diurnal variation of photosynthetic rate(Pn) and transpiration rate(Tr)ofSpiraeaalpineappeared to be a marked double-peak curves,but the diurnal variation of Pn and Tr ofSibiraeaangustataappeared to be single-peak curves,there were all obvious midday depression.The average values of Pn and Tr values were significantly lower inSpiraeaalpinethan that inSibiraeaangustata.With the diurnal light increase,photoinhibition of photosynthesis was intensified in two shrubs as indicated by the decrease of Fv/Fm.This kind of photoinhibition resulted from photodamage,because Fv/Fm at dusk could be not recovered to the levels at predawn.Path coefficients between Pn and eco-physiological factors revealed that the primary influencing factors for Pn in two shrubs were Tr,Tleafwas the main limiting factor for Pn.Moreover,the results showed thatSpiraeaalpinekept lower leaf area,lower Pn,lower Tr and higher water use efficiency(WUE) under soil water stress with high temperature and low water,so as to adapt to the stress environment.But,Sibiraeaangustatawas characterized by higher Pn,higher Tr,lower WUE and higher saturating photon flux(Ek),moreover it exhibited higher photosynthetic efficiency(α) at low light intensity,indicating that the photosynthesis of this species had a wider range of light adaptability.
alpine shrub meadow; photosynthetic rate; water use efficiency
2015-12-26
四川省教育厅自然科学基金项目 (13ZB0346);四川景观与游憩研究中心项目(JGYQ2015009)
高丽楠(1983-),女,内蒙古呼伦贝尔人,讲师,博士,主要从事植物生理生态学与环境保护方面的研究。 E-mail:gaolinan20081212@163.com
Q945.79
A
1004-3268(2016)06-0096-09
